
Arotupin Daniel Juwon
Department of Microbiology, Federal University of Technology, PMB704, Akure, Nigeria
Ogunmolu Funso Emmanuel
Department of Microbiology, Federal University of Technology, PMB704, Akure, Nigeria
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 1 | Page No.: 57-61







ABSTRACT
Fungi associated with tar sand samples from Ondo State, Nigeria were isolated and identified using standard microbiological method (serial dilution-spread plate technique). Also the isolates were screened for some enzymes of biotechnological importance. The fungal occurrence and frequency of the fungal isolates from the tar sand samples are Trichoderma viride BITRS-1001 (21%), Aspergillus fumigatus BITRS-1003 (10%), Rhizopus nigricans BITRS-1004 (10%), Penicillum italicum BITRS-1005 (16%), Spondylocladiella botrytioides BITRS-1006 (5.4%), Paecilomyces spp. BITRS-1007 (2.7%), Aspergillus flavus BITRS-1008 (8.1%), Mycotypha microsporium BITRS-1009 (13.5%), Articulospora inflata BITRS-1010 (5.4%) and Candida albicans BITRS-1002 (2.7%). All the isolates elaborated enzymatic activity, which were ranked as follows: Amylase 100% protease 100%, lipase 90%, polygalacturonase 60% and pectin methylesterase (60%). This study revealed the array of fungi associated with tar sand and their ability to elaborate hydrolytic enzymes. Harnessing these enzymes from the isolates would provide a clue to the biotransformation of essential nutrients of tar sand in addition to their application in industries.
PDF Abstract XML References Citation
Received: August 02, 2011;
Accepted: October 19, 2011;
Published: January 20, 2012
How to cite this article
Arotupin Daniel Juwon and Ogunmolu Funso Emmanuel, 2012. Screening of Fungal Isolates from Nigerian Tar Sand Deposit in Ondo State for Novel Biocatalysts. Journal of Biological Sciences, 12: 57-61.
DOI: 10.3923/jbs.2012.57.61
URL: https://scialert.net/abstract/?doi=jbs.2012.57.61
DOI: 10.3923/jbs.2012.57.61
URL: https://scialert.net/abstract/?doi=jbs.2012.57.61
INTRODUCTION
Microbes are a rich source of new biocatalysts (enzymes). In nature, microorganisms have been endowed with vast potentials. They produce an array of enzymes, which have been exploited commercially over the years. In recent years, the potentials of using microorganisms as biotechnological sources of industrially relevant enzymes has stimulated interest in the exploration of extracellular enzymatic activity in several microorganisms (Akpan, 2004; Jayani et al., 2005; Alva et al., 2007). Extracellular enzymes may be produced in liquid or solid media. However, the use of solid media permits a fast screening of large populations of fungi, allowing the detection of specific enzymes and helping in the chemotaxonomical differentiation of many microorganisms (Alves et al., 2002).
Although, enzymes have traditionally been extracted from plants and animals, microbial enzymes has formed the basis of commercial enzyme production due to the increasing availability of these microorganisms, their ease of improvement by the manipulation of their genes and environment, great diversity of enzymes have been found that cannot be obtained from plant and animal sources and high production capability at low cost among others (Onyeocha and Ogbonna, 1983; Alves et al., 2002; Akinyosoye et al., 2003; Carrim et al., 2006). Thus, microbial enzymes have found its application in textile (amylase, cellulase, oxidoreductase); detergents (protease, lipase, cellulase, oxidoreductase); food (pectinase, protease, cellulase, oxidoreductase); paper (xylanase, oxidoreductase and lipase) and leather (protease, lipase) industries (Alves et al., 2002).
Soils are the particulate materials of the outer crusts of the earth surface formed through the continuous weathering of the underlying parent rock (Arotupin et al., 2008). It overlies the earth’s bedrock and contains little organic matter (Robert et al., 2006). The soil is such a diverse environment that the microbial populations differ tremendously from soil to soil even within the same soil over the course of a season. The vast differences in the composition of soils, physical characteristics and the agricultural practices by which they are cultivated, result in corresponding large differences in the microbial population both in the numbers and kinds (Pelczar et al., 1993; Gokalp et al., 2010).
Tar sand is composed of sand, bitumen and clays that are rich in minerals and water (Oboh et al., 2006). It is present in vast amount in South Western Nigeria totaling about the largest deposit in the World (Adebiyi et al., 2005). It has an estimated reserve of about forty billion barrels of bituminous sand in place (Oguntimehin and Ipinmoroti, 2007). Several types of microorganisms have been found in association with the Nigerian tar sand deposit with their oil degrading ability reported by Oboh et al. (2006) and Adebayo et al. (2009). However, little or no work has been done in exploring these microorganisms as potential sources of industrially important enzymes.
This study therefore is aimed at isolating fungi associated with the tar sand samples from Ondo State, Nigeria and the evaluation of enzymes production namely amylase, lipase, protease, polygalacturonase and pectin methylesterase by the isolates
MATERIALS AND METHODS
Sources of soil samples: Soil samples were collected from five different sites with one serving as the control at the tar sand deposit in Gbelejuloda-Irele, Ondo State Nigeria. The samples were collected in sterile containers and transported to the Department of Microbiology, Federal University of Technology Akure for immediate laboratory investigations. Stock samples were kept in the refrigerator at 4°C.
Collection of samples: Surface of the soil sample sites for each of the plots was cleared and samples were obtained using the soil auger at depths of 10 cm intervals till a depth of 30 cm (Arotupin, 2007).
Isolation of fungi from soil samples: Fungal isolation was carried out on the various soil samples using the standard method of serial dilution spread plate technique. A 10 g of the soil samples were mixed with 90 mL of sterile distilled water. The suspension was thoroughly mixed and used for the isolation of associated fungi. A 0.1 mL of dilutions 10-2 and 10-3 was plated on Sabouraud Dextrose Agar (SDA) to which 0.1% tetracycline has been incorporated to inhibit bacterial growth (Cappucino and Sherman, 1998). The plates were incubated at 28±2°C for 72 h. Fungal colonies that developed were counted and sub-cultured to obtain pure cultures. The pure cultures of the fungal isolates were identified based on the cultural and morphological characteristics according to Barnett and Hunter (1972). The culture was maintained on Sabouraud Dextrose Agar (SDA) slants incorporated with 0.1% tetracycline kept at 4°C and subcultured at regular intervals.
Screening of fungal isolates for hydrolytic enzymes production: The isolated fungi were screened for the production of the following hydrolytic enzymes namely; amylase, protease, lipase, polygalacturonase and pectin methylesterase. The enzymatic activities were performed by initially growing the isolates on SDA for 72 h at 28±2°C. After that, 15 mm plug of the actively growing fungus was inoculated on the specific culture media for each enzyme to be investigated. The culture plates were incubated at 28±2°C for 96 h. The production of enzyme was determined within the incubation period, according to specific methodologies for each investigated enzyme (Onyeocha and Ogbonna, 1983; Alves et al., 2002; Akpan, 2004; Carrim et al., 2006; Damaso et al., 2008).
Determination of amylolytic activity: The method used was described by Alves et al. (2002) and Carrim et al. (2006). The isolates were inoculated in Nutrient Agar (NA) with 0.2% of soluble starch (g L-1), pH 6.0. After incubation, the cultures were treated under iodine solutions, which allowed the visualization of clear halos around the colonies.
Determination of lipolytic activity: The media used was as described by Carrim et al. (2006) and Damaso et al. (2008) containing (g L-1): peptone 10.0, NaCl 5.0, CaCl2 2H2O 0.1, agar 18.0, pH 7.4. To the sterilized culture media, previously sterilized olive oil was added in a final concentration of 1% (v/v). This medium was inoculated with the isolates and the presence of halos observed for lipases.
Determination of proteolytic activity: A medium that contained gelatin as the protein substrate was used to detect the production of proteolytic enzymes (Onyeocha and Ogbonna, 1983). The medium consisted of nutrient agar (1.5%), gelatin Gurr’s (0.4%) at pH 6.0. A stock solution of 8% gelatin was prepared by dissolving 4 g of it in 50 mL of water. This was sterilised separately. It was then mixed with pre-sterilised nutrient agar in the ratio of 5ml of 8% gelatin to 50 mL of nutrient agar. This was poured in to plates, allowed to solidify, inoculated with the isolates and incubated for 3 days. After incubation, complete degradation of gelatin was manifested by a clear zone around the colonies. However, observation of the clear zone was enhanced by flooding the plates with an aqueous saturated solution of ammonium sulphate (769 g L-1). This formed a precipitate with the intact gelatine, making the agar more opaque and leading to the development of clear zones around the colonies, the sizes of which were recorded.
Determination of polygalacturonase activity: The method employed was that described by Onyeocha and Ogbonna (1983) and Carrim et al. (2006). The composition of the medium included mineral salt solution (500 mL), peptone (1 g), agar (15 g), pectin (5 g) and distilled water (500 mL). The medium was brought to pH 5.0 with 0.1 M HCl and 0.1M NaOH. The plates were inoculated and incubated at 30°C for 96 h. After this, 5.0 mL of HCl (2 mol L-1) was added to the plates and the presence of clear halo around the colonies was indicative of the degradation of pectin.
Determination of pectin methylesterase activity: The medium for pectin methylesterase detection contained the basal medium including 1% pectin and 2% agar at pH 6. After incubation at 30°C, the plates were treated with 1% copper acetate. A clear zone around the fungal colony under a bluish background was recorded as a positive test. The sizes of the zones were recorded (Onyeocha and Ogbonna, 1983).
Statistics: The data of the halo diameter averages obtained during the investigations were subjected to Analysis of Variance and inferences made at p<0.05 using the SPSS 15.0 software package. Duncan’s New Multiple Range Test was used to separate means.
RESULTS
The fungal isolates and frequency of occurrence from the tar sand samples are shown in Table 1 with predomination of Trichoderma viride BITRS-1001 (21%), Aspergillus fumigatus BITRS-1003 (10%), Rhizopus nigricans BITRS-1004 (10%), Penicillum italicum BITRS-1005 (16%), BITRS-1006 (5.4%), Paecilomyces spp. BITRS-1007 (2.7%), Aspergillus flavus BITRS-1008 (8.1%), Mycotypha microsporium BITRS-1009 (13.5%), Articulospora inflata BITRS-1010 (5.4%) and Candida albicans BITRS-1002 (2.7%). No fungus consistently occurred in all the soil sample sites. T. viride occurred in sample sites A1, A2, A3, B1, B2, E1, E2 and E3; Aspergillus fumigatus in A2, B1, E1 and E3; Rhizopus nigricans in A2, C2, D2 and E2. Also, Penicillum italicum occurredin A3, B2, C2, D2, E1 and E2; Spondylocladiella botrytioides in C2 and D2; Mycotypha microsporium in A3, B3 C1, C2 and C3; Aspergillus flavus in A1, A3 and C1; Articulospora inflata in C3, C1 and D3; while Candida albicans and Paecilomyces occurred in C1 and D1, respectively.
In the screening of the fungal isolates for the production of the hydrolytic enzymes (Table 2), amylolytic activity was observed in all the isolates.
However, Mycotypha microsporium (BITRS-1009), Trichoderma viride (BITRS-1001) and Candida albicans (BITRS-1002) had the highest enzyme activity with means not significantly different at p<0.05.
| Table 1: | Occurrence of the fungal isolates in the soil samples |
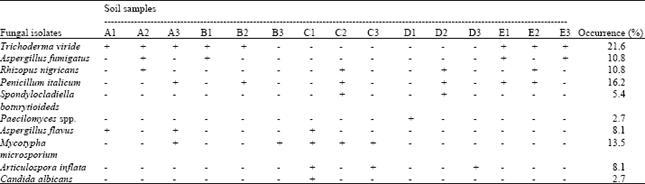 | |
| +: Present: -: Absent: A1 plot A (0 to 10 cm), A2: plot A (11 to 20 cm), A3: plot A (21 to 30 cm), B1 plot B (0 to 10 cm), B2: plot B (11 to 20 cm), B3: plot B (21 to 30 cm), C1 plot C (0 to 10 cm), C2: plot C (11 to 20 cm), C3: plot C (21 to 30 cm), D1 plot D (0 to 10 cm), D2: plot D (11 to 20 cm), D3: plot D (21 to 30 cm), E1 Control (0 to 10 cm), E2: Control (11 to 20 cm), E3: Control (21 to 30 cm) | |
| Table 2: | Average diameter halo (cm) for the enzymatic activities of the isolates |
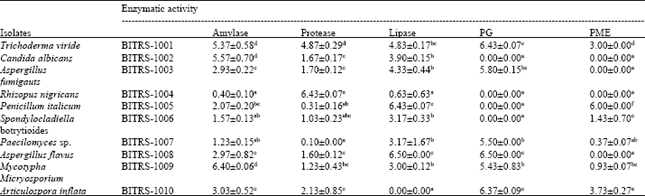 | |
| Values are as Mean±SD. PG: Polygalacturonase; PME: Pectin Methylesterase. Numbers followed by the same letter among the isolates for each type of enzyme were not significantly different according to New Duncan Multiple Range Test at p≤0.05. n = 3 | |
All the isolates equally showed evidence of protease activity, but the highest protease activity was recorded with Rhizopus nigricans (BITRS-1004) showing halo diameter significantly different from the other isolates at p<0.05. Lipase activity was recorded in 90% of the isolates with Aspergillus flavus (BITRS-1008) and Penicillum italicum(BITRS-1005) presenting the highest lipase activity with a halo diameter of 6.50 cm and 6.43 cm while Articulospora inflata (BITRS-1010) had no lipase activity.
Poly Galacturonase (PG) and Pectin Methyl Esterase (PME) activity were recorded in 60% of the isolates respectively (Table 2). Trichoderma viride (BITRS-1001), Aspergillus flavus (BITRS-1008), Articulospora inflata (BITRS-1010), Mycotypha microsporium (BITRS-1009), Paecilomyces spp. (BITRS-1007) and Aspergillus fumigatus (BITRS-1003) ranked highest in PG activity with halo diameters not significantly different at p<0.05 from each other, while Spondylocladiella botrytioides (BITRS-1006), Penicillum italicum (BITRS-1005), Rhizopus nigricans (BITRS-1004) and Candida albicans (BITRS-1002) showed no detectable polygalacturonase production. In the case of pectin methylesterase, Penicillum italicum (BITRS-1005) exhibited the highest PME activity at levels significantly different from all other isolates at p<0.05. Candida albicans (BITRS-1002), Aspergillus fumigatus (BITRS-1003), Rhizopus nigricans (BITRS-1004) and Aspergillus flavus (1008) showed no signs of PME production (Table 2).
DISCUSSION
The results obtained showed that fungi constitute part of the normal resident of the tar sand soil samples under investigation and that they can be isolated and cultivated for industrial exploitation. Although, the presence of toxic metals and harmful chemical substances in the tar sand sample present an unfavourable environment to these fungi (Adebiyi et al., 2005; Oguntimehin and Ipinmoroti, 2007). Their isolation from this environment shows that these fungi have evolved strategies of adapting to the environment and/or utilizing these substances as energy sources. Fungi as a result of their competitive saprophytic ability expressed by fast mycelia growth, spores production, presence of efficient and extensive systems of powerful enzymes are able to utilize complex chemical substances as energy sources. This ability of fungi make them potentially important in nature (Akinyele and Adetuyi, 2005; Da Silva et al., 2005; Khalid et al., 2006).
The results of this investigation clearly showed that different genera and species of fungi are associated with or found in the tar sand samples. A sum of ten (10) fungi was isolated from the tar sand as well as the control plots. The fungal species isolated were Articulospora inflata BITRS-1010, Aspergillus flavus BITRS-1008, Aspergillus fumigatus BITRS-1003, Candida albicans BITRS-1002, Mycotypha microsporium BITRS-1009, Paecilomyces spp. BITRS-1007, Penicillium italicum BITRS-1005, Rhizopus nigricans BITRS-1004, Spondylocladiella botrytioides BITRS-1006 and Trichoderma viride BITRS-1001. Close observation revealed T. viride BITRS-1001 to be dominant and the most frequently encountered fungi in contrast with the findings of Oboh et al. (2006) who reported the dominance of Aspergillus spp. of the quadri-Trichoderma, Aspergillus, Penicillum and Rhodotorula spp. isolated from tar sand samples from riverine areas of the state. However, the isolation of these fungi from the soil shows that these fungi are physiologically and genetically equipped to cope with the harshness in the environment and that the tar soil constitutes a suitable substratum for the growth of the fungi.
In the screening of the fungal isolates for the production of the hydrolytic enzymes, the method of radial diffusion in solid media indicated the activity of the various enzymes in qualitative form (Carrim et al., 2006). Although, extracellular enzymes may be produced in liquid or solid media, the use of solid media however permits a fast screening of large populations of fungi, allowing the detection of specific enzymes and helping in the chemo-taxonomical differentiation of many microorganisms (Alves et al., 2002). The observed result as shown in Table 2 indicates that all the isolates hydrolysed starch and gelatin in the medium, signalling the production of amylase and protease, respectively. This ability of the fungi to rapidly utilise starch have been attributed to the fact that starch is the most abundant organic carbon source in the environment serving as the major reserve carbohydrate for higher plants (Aiyer, 2004).
However, the ability of the several fungi to selectively produce the various hydrolytic enzymes of interest confirmed that microorganisms (fungi inclusive) are a rich source of new biocatalysts in nature, that have been endowed with vast potentials to produce arrays of enzymes (Akpan, 2004; Jayani et al., 2005). These have great potentials of being used as biotechnological sources of industrially relevant enzymes (Alva et al., 2007). Studies on the isolation, identification and screening of fungi associated with tar sand deposit in Ondo State Nigeria are rare. This study therefore, has provided information on the types of fungi associated with the tar sand deposit in Ondo State, Nigeria. The isolates are also capable of producing potentially useful hydrolytic enzymes. However, attention should not be centred on the exploration of the tar sand at the expense of the associated organisms (fungi) that possibly played an important role in the balancing of the economy of nature.
REFERENCES
- Adebayo, E.A., J.K. Oloke and D.A. Aina, 2009. Effects of culture parameters of a bac-terial consortium on biodegradation of bitumen. Adv. Environ. Biol., 3: 46-52.
Direct Link - Adebiyi, F.M., F.S. Olise, O.K. Owoade, I.B. Obioh, T.R. Ajayi, H.B. Olaniyi and O.I. Asubiojo, 2005. TXRF characterization of EDTA extractable metals from soils of bituminous sands occurrence area, agbabu, South Western Nigeria. J. Applied Sci., 5: 1151-1157.
CrossRefDirect Link - Aiyer, P.V.D., 2004. Effect of C.N ratio on alpha amylase production by Bacillus licheniformis SPT 27. Afr. J. Biotechnol., 3: 519-522.
Direct Link - Akinyele, B.J. and F.C. Adetuyi, 2005. Effect of agrowastes, pH and temperature variation on the growth of Volvariella volvacea. Afr. J. Biotechnol., 4: 1390-1395.
Direct Link - Alves, M.H., G.M. Campos-Takaki, A.L.F. Porto and A.I. Milanez, 2002. Screening of Mucor spp. for the production of amylase, lipase, polygalacturonase and protease. Braz. J. Microbiol., 33: 325-330.
CrossRef - Arotupin, D.J., F.A. Akinyosoye and A.K. Onifade, 2008. Purification and characterization of pectinmethylesterase from Aspergillus repens isolated from cultivated soil. Afr. J. Biotechnol., 7: 1991-1998.
Direct Link - Damaso, M.C.T., M.A. Passianoto, S.C. de Freitas, D.M.G. Freire, R.C.A. Lago and S. Couri, 2008. Utilization of agroindustrial residues for lipase production for solid-state fermentation. Braz. J. Microbiol., 39: 676-681.
Direct Link - Jayani, R.S., S. Saxena and R. Gupta, 2005. Microbial pectinolytic enzymes: A review. Process Biochem., 40: 2931-2944.
CrossRefDirect Link - Mahmood, K., Y. Wei-Jun, K. Nazir, R.Z. Iqbal and A.G. Arijo, 2006. Study of cellulolytic soil fungi and two nova species and new medium. J. Zhejiang Univ. Sci. B, 7: 459-466.
CrossRefPubMedDirect Link - Oguntimehin, I.I. and K.O. Ipinmoroti, 2007. Solvent extraction of vanadium from nigerian bitumen using tri-butylphosphate. J. Applied Sci., 7: 4028-4031.
CrossRefDirect Link - Da Silva, R., E.S. Lago, C.W. Merheb, M.M. Macchione, Y.K. Park and E. Gomes, 2005. Production of xylanase and cmcase on solid substrate fermentation in different residues by Thermoascus aurantiacus miehe. Braz. J. Microbiol., 36: 235-241.
Direct Link - Alva, S., J. Anupama, J. Savla, Y.Y. Chiu and P. Vyshali et al., 2007. Production and characterization of fungal amylase enzyme isolated from Aspergillus sp. JGI 12 in solid state culture. Afr. J. Biotechnol., 6: 576-581.
Direct Link - Carrim, A.J.I., E.C. Barbosa and J.D.G. Vieira, 2006. Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham. (Carobinha-do-campo). Braz. Arch. Biol. Technol., 49: 353-359.
CrossRefDirect Link - Gokalp, Z., M. Basaran, O. Uzun and Y. Serin, 2010. Spatial analysis of some physical soil properties in a saline and alkaline grassland soil of Kayseri, Turkey. Afr. J. Agric. Res., 5: 1127-1137.
Direct Link