
Mahdi Eskandarian Boroujeni
Department of Microbiology, Genohelix Biolabs,
A Division of Centre for Advanced Studies in Biosciences, Jain University,
127/2, Bull Temple Road, Chamarajpet, Bangalore-560019, India
Arijit Das
Department of Microbiology, Genohelix Biolabs,
A Division of Centre for Advanced Studies in Biosciences, Jain University,
127/2, Bull Temple Road, Chamarajpet, Bangalore-560019, India
K. Prashanthi
Department of Molecular Biology, Genohelix Biolabs,
A Division of Centre for Advanced Studies in Biosciences, Jain University, 127/2,
Bull Temple Road, Chamarajpet, Bangalore-560019, Karnataka, India
Sandeep Suryan
Department of Molecular Biology, Genohelix Biolabs,
A Division of Centre for Advanced Studies in Biosciences, Jain University, 127/2,
Bull Temple Road, Chamarajpet, Bangalore-560019, Karnataka, India
Sourav Bhattacharya
Department of Microbiology, Genohelix Biolabs,
A Division of Centre for Advanced Studies in Biosciences, Jain University,
127/2, Bull Temple Road, Chamarajpet, Bangalore-560019, India
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 1 | Page No.: 43-50







ABSTRACT
Actinomycetes constitute an economically important class of bacteria for their ability to produce various degradative enzymes and secondary metabolites. The present study was conducted with the objectives to isolate some soil actinomycetes, screen them for production of hydrolytic enzymes and characterize them. A total of 30 isolates of actinomycetes were obtained from six soil samples collected from Kuruva Island and the banks of Pookot Lake from Wayanad district in Kerala, India. Screening for enzyme production among 18 randomly selected isolates revealed that 13 of them were able to synthesize either cellulase, pectinase and/or xylanase. The results indicated that 44.4% of the total isolates produced all the three enzymes. Based on morphological and biochemical characterization, the active isolates were found to belong to the genus Streptomyces. The relatedness between the 13 active isolates was determined by RAPD fingerprinting using 20 different primers. Only amplification with primer OPAB-9 resulted in good scorable bands and revealed a prominent band at 600 bp in 54% of the isolates. The dendrogram showed that the active isolates fell into one super cluster which further consisted of three clusters and one out group. The presence of identical cultural characteristics was detected in most of the clusters. With respect to the enzyme activity, the similarity was only noticed among majority of the members of cluster 1. This molecular study may be employed as an effective tool to characterize streptomycete isolates capable of producing various industrially important enzymes.
PDF Abstract XML References Citation
Received: August 30, 2011;
Accepted: December 27, 2011;
Published: January 20, 2012
How to cite this article
Mahdi Eskandarian Boroujeni, Arijit Das, K. Prashanthi, Sandeep Suryan and Sourav Bhattacharya, 2012. Enzymatic Screening and Random Amplified Polymorphic DNA Fingerprinting of Soil Streptomycetes Isolated from Wayanad District in Kerala, India. Journal of Biological Sciences, 12: 43-50.
DOI: 10.3923/jbs.2012.43.50
URL: https://scialert.net/abstract/?doi=jbs.2012.43.50
DOI: 10.3923/jbs.2012.43.50
URL: https://scialert.net/abstract/?doi=jbs.2012.43.50
INTRODUCTION
Actinomycetes are classified as vast groups of gram positive bacteria which are filamentous in appearance and ubiquitously present in both terrestrial and aquatic environments. Streptomycetes constitute a major class of soil actinomycetes which produce numerous natural metabolites such as antibiotics and enzymes (Lo et al., 2002; Chiani et al., 2010).
Streptomycetes are able to utilize a wide range of organic compounds as carbon sources, including complex biological polymers such as cellulose and lignin (Lignell, 2008). Cellulolytic potential of these bacteria has been extensively studied and implemented in various commercial processes (Alam et al., 2004).
Bacterial enzymes such as cellulases, pectinases and xylanases have evoked scientific interest for their practical applicability in textile, beverage, food and feed industries (Coughlan and Hazlewood, 1993; Blanco et al., 1995; Saadoun et al., 2007). Microbial pectinases are regarded as one of the economically significant food enzymes, particularly involved in the juice recovery, clarification and enhancement of nutritional quality of foods (Tochi et al., 2009; Murad and Azzaz, 2011).
There has been rapid increase in xylanase application due to its necessity for the products produced from dissolving pulp such as rayon, cellophane and chemicals (Beg et al., 2001; Subramaniyan and Prema, 2002). Such enzymes provide eco-friendly alternatives for effective bleaching of paper pulp without the use of toxic chlorine compounds and without adversely affecting the quality of the paper pulp. Moreover, microbial cellulases have gained significant attention due to their wide applicability in various industrial processes including pulp and paper, textile, laundry, biofuel production, food and feed industry, brewing and agriculture (Kuhad et al., 2011).
Molecular characterization based on RAPD fingerprinting is frequently used nowadays to study the ecological distribution and diversity of microorganisms. It is considered as a convenient and cost-effective typing method employed in identification of bacterial species (Gillespie and Oliver, 2004; Kannan and Vincent, 2011).
In this study, various streptomycetes isolated from the forest soils of Wayanad district in Kerala, India were screened for their potential to produce the fiber hydrolytic enzymes (cellulases, pectinases and xylanases). These isolates were further characterized by both traditional (morphological and biochemical) and molecular methods.
MATERIALS AND METHODS
The present study was conducted during the period from 20.12. 2010 to 11.07. 2011 at Genohelix Biolabs, A Division of Centre for Advanced Studies in Biosciences, Jain University, Chamarajpet, Bangalore, Karnataka, India.
Collection of soil samples: Soil samples (dry and wet) were collected from 6 different locations of Kuruva Island (11°49'18" N, 76°5'32" E) and the banks of Pookot Lake (11°32'30" N, 76°1'37" E) of Wayanad district in Kerala, India, using pre-sterilized zip lock covers and sterile spatula. Precautionary measures were taken to minimize the contamination. All the samples were stored at 4°C and processed within 24 h.
Sample processing: Each soil sample was air dried at room temperature, then mixed thoroughly and sieved through a 2 mm pore size sieve (Retsch, Haan, Germany) to get rid of large debris. The sieved soil was used for the isolation processes.
Isolation of actinomycetes: One gram each of the treated soil samples was suspended in 100 mL sterile distilled water and then incubated in an orbital shaker incubator (Orbitek) at 28°C with shaking at 200 rpm for 1 h (Oskay et al., 2004). Mixtures were allowed to settle and then serial dilutions of the spore suspensions were prepared up to 10-4. From each dilution, 0.1 mL was taken and spread evenly over the surface of starch casein nitrate agar (SCNA) (El-Nakeeb and Lechevalier, 1963; Kuster and Williams, 1964) plates (supplemented with cycloheximide 50 μg mL-1) (in triplicate) with sterile L-shaped glass rod, then incubated at 28°C for 10 days. Dilutions that gave 30-300 colonies were chosen for further isolation. Actinomycete isolates were purified by streak-plate technique and the pure cultures were maintained on SCNA slants at 4°C for further use.
Screening for xylanase-producing actinomycetes: Pure isolates of actinomycetes were cultured on xylan agar containing (g L-1): yeast extract, 1; xylan, 10; KH2PO4, 4; NaCl, 2; MgSO4.7H2O, 1; MnSO4, 0.05; FeSO4.7H2O, 0.05; CaCl2.2H2O, 2; NH4Cl, 2 and agar, 20; pH 7.3 and then incubated at 28°C for 5 days (Nanmori et al., 1990). The plates were then flooded with absolute ethanol (99%) and left for 1 h at room temperature. Colonies producing xylanase enzyme showed clear zones against an opaque colour of non-hydrolyzed medium. Positive isolates were tested again for confirmation.
Screening for pectinase-producing actinomycetes: Pure isolates of actinomycetes were cultured on pectin agar containing (g L-1): yeast extract, 1; pectin, 5; KH2PO4, 4; NaCl, 2; MgSO4.7H2O, 1; MnSO4, 0.05; FeSO4.7H2O, 0.05; CaCl2.2H2O, 2; NH4Cl, 2 and agar, 20; pH 7.3 and then incubated at 28°C for 5 days. Plates were then flooded with 1% solution of polysaccharide precipitant CTAB (cetyl trimethyl ammonium bromide), dissolved in a 15% alcoholic solution and then used to detect pectinase production. After 1 h of exposure, colonies producing pectinase showed a clear zone against an opaque colour of the non-hydrolyzed medium.
Screening for cellulase-producing actinomycetes: Pure isolates of actinomycetes were cultured on cellulose agar containing (g L-1): yeast extract, 1; CMC, 10; KH2PO4, 4; NaCl, 2; MgSO4.7H2O, 1; MnSO4, 0.05; FeSO4.7H2O, 0.05; CaCl2.2H2O, 2; NH4Cl, 2 and agar, 20; pH 7.3 and then incubated at 28°C for 5 days. The plates were then flooded with 0.1% Congo Red and left for 30 min, washed with 1 mL NaCl (1 M) and left for 15 min. Colonies producing cellulase showed clear zones against red colour of non-hydrolyzed medium. Positive isolates were tested again for confirmation (Carder, 1986).
Morphological characterization of the isolates: Only actinomycete isolates that gave a positive result for cellulase, pectinase and/or xylanase were characterized morphologically and physiologically following the directions given by the International Streptomyces Project (ISP) according to Shirling and Gottlieb (1966). Morphological characters of isolates were observed by smears from colonies up to 10 days, stained by Gram’s method as described by Hucker and Conn (1923).The spore chain morphology was determined by direct microscopic examination using the 10 days old cultures under a compound light microscope (Nikon, Japan) using 1000X magnification power. The observed structures were compared with Bergey’s Manual of Determinative Bacteriology, ninth edition (Holt et al., 1994) and the organisms were identified. Color of aerial mycelia was determined from mature, sporulating aerial mycelia of the actinomycetes colonies on SCNA (Pridham, 1965) and the isolates were grouped into separate color series according to the system proposed by Nonomura (1974). Color of the substrate mycelia (reverse of the plate) was also observed along with diffusible pigments if any (Padmadhas, 2010).
Biochemical characterization of the isolates: Various biochemical tests performed for the identification of the potential isolates were indole production, methyl red, Voges Proskauer, citrate utilization, casein hydrolysis, starch hydrolysis, urea hydrolysis, gelatin hydrolysis, H2S production, acid production from sugar, NaCl resistance and temperature tolerance (APHA, AWWA and WEF, 2005).
Growth conditions: All Streptomyces isolates that showed unusual fiber hydrolytic enzyme profile were cultured on tryptic soy broth (TSB) (30 g L-1) (Hopwood et al., 1985) at 28°C with shaking at 140 rpm for 48 h. Purity of the cultures was confirmed by plating 0.1 mL from the broth on starch casein nitrate agar (SCNA) (Kuster and William, 1964) plates and incubated at 28°C for 72 h.
Extraction of genomic DNA from the actinomycete isolates: The potent actinomycete isolates were cultured in SCN broth and incubated at 28°C for 7 days in an orbital shaker at 130 rpm. Genomic DNA were extracted using Bacterial Genomic DNA Isolation Kit RKT09 (Chromous Biotech Pvt. Ltd., Bangalore, India) according to the manufacturer instructions and visualized using 0.8% (w/v) agarose gel electrophoresis.
Characterization of the actinomycete isolates using random amplified polymorphic DNA (RAPD): The potent isolates of actinomycetes were subjected to molecular characterization by the RAPD fingerprinting technique using 20 different random primers. The primer sequences have been listed out in Table 1. Each primer was 10-mer long (Sigma Aldrich, USA). In order to determine the type-ability, reproducibility and discrimination of each primer, separate amplifications of each primer were conducted (two trials for each primer). The output of each experiment was compared to the previous one (Arbeit, 1994). The PCR reactions were performed in volumes of 25 μL containing 10 picomoles of primer, 2.5 μL of 10X PCR Buffer (Chromous Biotech, India), 2.5 mM dNTP mix (Chromous Biotech, India), 1 U Taq DNA polymerase (Chromous Biotech, India), 10-15 μg mL-1 of template DNA.
| Table 1: | Sequences of RAPD primers used in the study |
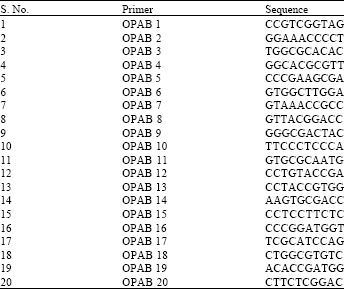 | |
Nuclease free water (Chromous Biotech, India) was used to bring the reaction mixture to 25 μL.
PCR amplification was carried out in 0.2 mL thin walled, nuclease free PCR tubes (Molecular Bio Products, USA) using thermocycler (MJ Research PTC-200, USA) programmed as follows: initial denaturation at 95°C for 5 min, followed by 40 cycles of denaturation at 94°C for 1 min, annealing at 30°C for 1 min, extension at 72°C for 2 min. Finally, after an extension at 72°C for 5 min, the tubes were held at 4°C for direct use or stored at -20°C until used.
Electrophoresis and photography: The amplified PCR products were separated on 1.2% (w/v) agarose gels stained with ethidium bromide. Gels were viewed and documented using gel documentation unit (Herolabs, Germany).
Data analysis of RAPD profiles: The RAPD gel photographs were analyzed by Easy Win 32 software supplied with horizontal and vertical rulers. Data were then normalized and further processed by the Gel Compare 3.1 Program (Applied Maths, Kortrijk, Belgium) (Vauterin and Vauterin, 1992). Patterns were analyzed by band based Dice method and clustered using the unweighted pair group method with arithmetic averages (UPGMA) (Sokal and Michener, 1958).
RESULTS
Isolation of streptomycetes: After processing of soil samples, a total of 30 different actinomycetes (streptomycete-like colonies) were isolated. All of these isolates were selected based on their colony morphology, resembling that of Streptomyces species. Randomly selected 18 streptomycete isolates were screened for production of cellulase, pectinase and xylanase enzymes.
Screening for the fiber-hydrolytic enzymes production by streptomycetes: Among the 18 isolates, 13 isolates showed promising enzyme activity (cellulases, pectinases and xylanases) and they were subjected to further characterization by conventional and molecular methods (Table 2).
| Table 2: | Screening for hydrolytic enzymes produced by different Streptomyces isolates |
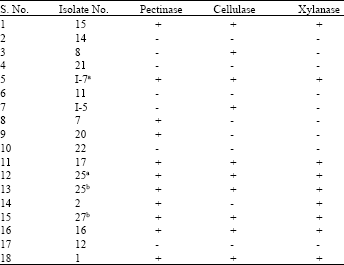 | |
| aThe production of pectinase and xylanase was found to be highest in Isolate I-7 with zones of 20 and 15 mm, respectively. bThe production of cellulase was maximum in Isolate 27 with a zone of 21 mm | |
Data showed that 13 (72.2%) of 18 isolates were active and could produce hydrolytic enzymes. The enzyme-producing ability of the isolates revealed, 22.2% were able to produce one enzyme, 5.5% produced two different enzymes and 44.4% of the 18 isolates were able to produce all the three enzymes. Results indicated that 61.1% of the isolates were able to produce pectinase, whereas 55.5% produced cellulase enzyme and only 50% produced xylanase (Table 3). The relative activities of the isolates to produce the enzymes ranged from weak, moderate to strong depending on the diameter of the clear zone formed in the screening process as seen in Fig. 1.
Morphological characterization of the selected streptomycete isolates: After 10 days of incubation on SCNA, the colonies with characteristic color of actinomycetes such as white, grey, black, brown, yellow, tan, pink and green were observed, as illustrated in Table 4.
| Table 3: | The percentage of Streptomyces isolates producing the fiber-hydrolytic enzymes |
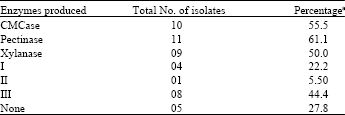 | |
| I, II and III demonstrate the ability of Streptomyces isolates to produce one, two and three fiber-hydrolytic enzymes, respectively. aThe values have been calculated considering the total of 18 isolates | |
 | |
| Fig. 1: | Plate diffusion method showing the fiber hydrolytic enzymes activity. (a) Detection of CMCase production, (b) Detection of pectinase production and (c) Detection of xylanase production |
| Table 4: | Characteristics of 13 active isolates of Streptomyces |
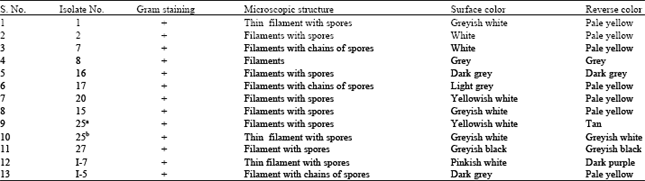 | |
| +: Positive | |
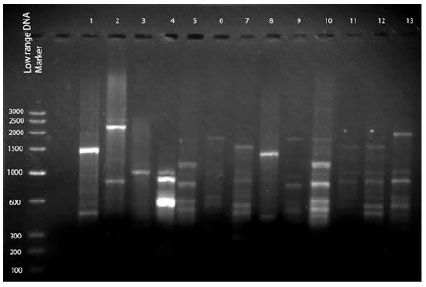 | |
| Fig. 2: | RAPD fingerprints profile of Streptomyces isolates on 1.2% agarose gel electrophoresis, amplified using primer OPAB 09. Lane Marker: 100 bp DNA ladder molecular weight marker; lane 0: PCR negative control; lanes 1-13 correspond to the Streptomyces isolates; lane 1: 1, lane 2: 16, lane 3: 27, lane 4: 25b, lane 5: 17, lane 6: I-7, lane 7: 15, lane 8: 8, lane 9: I-5, lane 10: 7, lane 11: 20, lane 12: 25a, lane 13: 2 |
The 13 fiber-hydrolytic enzymes producing-streptomycete isolates were categorized into 3 general colour series depending on the colour of mature sporulated aerial mycelia: white (including greyish, yellowish and pinkish) (61.5%), grey (dark and light) (30.7%) and greyish black (7.6%). Moreover, some of the isolates demonstrated pigmentation on the reverse of the colonies. Regarding the colour of the substrate mycelium (reverse colour), 53.8 and 15.3% of the isolates showed pale yellow and grey color, respectively. However, the lowest occurrence was for tan, black and purple colour series and was represented by 7.6% for each, shown in Table 5.
Biochemical characterization of the isolates: Results of the biochemical tests revealed all the selected actinomycete isolates belonged to the genus Streptomyces (data not shown).
Extraction of genomic DNA from Streptomyces isolates: Genomic DNA were extracted using commercial DNA isolation kit. The extracted DNA were of good quality as checked by agarose gel electrophoresis. The extracted DNA were intact, clean and free of RNA.
RAPD study: The extracted genomic DNA from 13 isolates were used to perform RAPD-PCR. The PCR conditions were optimized and screened for reproducible amplification with 20 different RAPD primers. Only amplification with primer OPAB-9 resulted in good scorable bands, as depicted in Fig. 2.
| Table 5: | The percentage of 13 enzyme- producing Streptomyces isolates exhibiting different pigmentation |
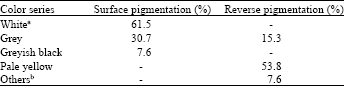 | |
| aIncludes greyish, yellowish and pinkish white. bRepresents tan, black and purple color series | |
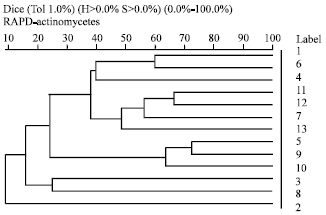 | |
| Fig. 3: | Dendrogram using average linkage between groups, numbers 1-13 represent the isolates (1: 1; 6: I-7; 4: 25b; 11: 20; 12: 25a; 7: 15; 13: 2; 5: 17; 9: I-5; 10: 7; 3: 27; 8: 8; 2: 16) |
The RAPD-fingerprints of the various strains differed in fragments number, size and intensity depending on which primer being used in each run. Each RAPD experiment was repeated thrice with each primer and similar results were obtained.
The number of polymorphic bands observed for each isolate was between 3 and 10 with a size ranging from 100 to 2000 bp. Amplification with primer OPAB 9 revealed a prominent band at 600 bp in 54% of the isolates. Furthermore, many sets of monomorphic bands were observed in the range of 500 and 1000 bp.
The dendrogram presented in Fig. 3 shows that the tested isolates fell into one super cluster. This super cluster consists of three clusters and one out group. The first cluster includes 7 isolates (1, I-7, 25b, 20, 25a, 15 and 2). The second cluster contains 3 isolates (17, I-5 and 7). The third cluster includes only 2 isolates (27 and 8). Isolate 16 was considered as an out group to cluster 3.
DISCUSSION
Actinomycetes are frequently observed in soil with high organic matter and low moisture contents (Lee and Hwang, 2002). In the present study a high count of actinomycetes was obtained from the island soil and lake sediments. This may be attributed to the presence of thick vegetation on the island and partly saline soil rich in humus content. Abundance of this organic matter in soil and sediments is essential for the enrichment of the favorable niche of saprophytic actinomycetes. The actinobacterial diversity has been extensively investigated in different regions of the southern states of India (Sivakumar et al., 2007).
Selection of a potent actinomycete strain capable of producing three fiber hydrolytic enzymes (cellulase, xylanase and pectinase) is vital to the industrial and biotechnological applications (Beg et al., 2000).
The results of morphological characterization indicate that the white and the grey color series were the most recurrent. Similar results were reported for Streptomyces isolates that have been recovered from other locations in Kerala (Arunachalam et al., 2010). Also, Remya and Vijayakumar (2008) reported, among 173 strains of actinomycetes obtained from marine and mangrove ecosystems, 65.6% of the isolates belonged to white and grey color series. These studies were virtually in line with our findings.
Dendrogram analysis is frequently applied in computational biology to elucidate the relatedness among microorganisms (Khucharoenphaisan et al., 2011). Results of the dendrogram revealed identical cultural and microscopic characteristics among all the members in each cluster except cluster 3. As observed, all the isolates within cluster 1 and 2 exhibited filaments with spores and filaments with chains of spores, respectively. These results may also be correlated with the banding patterns as evident in the RAPD profile.
Evaluation of colony pigmentation is one of the efficient approaches for characterization of Streptomyces isolates (Pridham and Tresner, 1974). Subsequently, it was obvious that surface color appeared to be white with a very slight variability in all the isolates of cluster 1. However, as to reverse color, only 4 out of 7 showed pale yellow. In the case of cluster 2, all the isolates had pale yellow reverse color. Moreover, the isolates 27 and 8 in cluster 3, with respect to the color of aerial and substrate mycelia, exhibited grey and greyish black, respectively.
With respect to the enzyme activity, most of the isolates within cluster 1 were able to produce all the three fiber hydrolytic enzymes (5 out of 7). Also, I-7 with the highest pectinase and xylanase activities fell into this cluster, whereas, isolate 27 with the highest cellulase production was seen in cluster 3.
RAPD profiles for the 13 fiber-hydrolytic enzymes producing isolates were analyzed by UPGMA dendrograms. This analysis enabled a simultaneous comparison between these isolates. In the current study, the RAPD-PCR with primer OPAB 9 resulted in a common band at 600 bp in nearly majority of the isolates, particularly in both of the most active isolates (I-7 and 27). The size and the number of bands were not in agreement with the results reported by Saadoun et al. (2007). This dissimilarity in results may be caused by the employment of various PCR reaction conditions such as different primers and PCR programs. The reproducibility of the generated randomly amplified polymorphic DNA fragments is considered as a major obstacle due to the effect of countless experimental variables on the results of fingerprinting. Thus, it is recommended to develop suitable RAPD protocols along with sequence-based analysis for identification and phylogenetic studies.
CONCLUSION
Eighteen different streptomycete isolates were obtained from the forest and lake soils of Wayanad district of Western Ghats. Among these, 13 isolates demonstrated the potential to produce different fiber-hydrolytic enzymes such as cellulase, pectinase and xylanase. The dendrogram analysis based on RAPD-fingerprinting revealed the presence of identical cultural characteristics in most of the clusters. However, with respect to the enzyme activity, the similarity was only noticed among majority of the members of cluster 1. This molecular study may be employed as an effective tool to characterize the active isolates of streptomycetes capable of producing various industrially important enzymes.
ACKNOWLEDGMENT
We wish to extend our sincere gratitude to Dr. Chenraj Jain, Chairman, Jain Group of Institutions, Bangalore, for providing us with the financial and laboratory facilities required for this research work. We also wish to thank all the faculties and the entire supporting staff of the laboratory whose help has been invaluable for the successful completion of our research work. We are thankful to Prof. V.R. Padmawar of Indian Statistical Institute, Bangalore for his technical advice and valuable suggestions.
REFERENCES
- Alam, M.Z., M.A. Manchur and M.N. Anwar, 2004. Isolation, purification, characterization of cellulolytic enzymes produced by the isolate Streptomyces omiyaensis Pak. J. Biol. Sci., 10: 1647-1653.
CrossRefDirect Link - APHA, A.D. Eaton, AWWA and WEF, 2005. Standard Methods for the Examination of Water and Wastewater. 21st Edn., APHA-AWWA-WEF, Washington, DC., USA., ISBN: 9780875530475,.
Direct Link - Arunachalam, R., E.G. Wesely, J. George and G. Annadurai, 2010. Novel approaches for identification of Streptomyces noboritoensis TBG-V20 with cellulase production. Curr. Res. Bacteriol., 3: 15-26.
CrossRefDirect Link - Beg, Q.K., B. Bhushan, M. Kapoor and G.S. Hoondal, 2000. Effect of amino acids on production of xylanase and pectinase from Streptomyces sp. QG-11-3. World J. Microbiol. Biotechnol., 16: 211-213.
CrossRefDirect Link - Beg, Q.K., M. Kapoor, L. Mahajan and G.S. Hoondal, 2001. Microbial xylanases and their industrial applications: A review. Applied Microbiol. Biotechnol., 56: 326-338.
CrossRefDirect Link - Tochi, B.N., Z. Wang, S.Y. Xu and W. Zhang, 2009. The influence of a pectinase and pectinase/hemicellulases enzyme preparations on percentage pineapple juice recovery, particulates and sensory attributes. Pak. J. Nutr., 8: 1184-1189.
CrossRefDirect Link - Blanco, A., T. Vidal, J.F. Colom and F.I.J. Pastor, 1995. Purification and properties of xylanase A from alkali-tolerant Bacillus sp. strain Bp-23. Applied Enviorn. Microbiol., 61: 4468-4470.
PubMedDirect Link - Carder, J.H., 1986. Detection and quantitation of cellulase by congo red staining of substrates in a cup-plate diffusion assay. Anal. Biochem., 153: 75-79.
CrossRefPubMedDirect Link - Chiani, M., A. Akbarzadeh, A. Farhangi, M. Mazinani, Z. Saffari, K. Emadzadeh and M.R. Mehrabi, 2010. Optimization of culture medium to increase the production of desferrioxamine B (desferal) in Streptomyces pilosus. Pak. J. Biol. Sci., 13: 546-550.
CrossRefDirect Link - Coughlan, M. and G.P. Hazlewood, 1993. β-1,4-D-Xylan-degrading enzyme systems: Biochemistry, molecular biology and applications. Biotechnol. Applied Biochem., 17: 259-289.
PubMed - El-Nakeeb, M.A. and H.A. Lechevalier, 1963. Selective isolation of aerobic actinomycetes. Applied Environ. Microbiol., 11: 75-77.
Direct Link - Gillespie, B.E. and S.P. Oliver, 2004. Comparison of an automated ribotyping system, pulsed-field gel electrophoresis and randomly amplified polymorphic DNA fingerprinting for differentiation of Streptococcus uberis strains. Biotechnology, 3: 165-172.
CrossRefDirect Link - Kannan, R.R. and S.G.P. Vincent, 2011. Molecular characterization of antagonistic Streptomyces isolated from a mangrove swamp. Asian J. Biotechnol., 3: 237-245.
CrossRefDirect Link - Khucharoenphaisan, K., U. Puangpetch, K. Puttaraksa and K. Sinma, 2011. Grouping of actinomycetes isolated from termites using biochemical character. J. Biol. Sci.
Direct Link - Kuhad, R.C., R. Gupta and A. Singh, 2011. Microbial cellulases and their industrial applications. Enzyme Res., 2011: 1-10.
CrossRefDirect Link - Kuster, E. and S.T. Williams, 1964. Selection of media for isolation of Streptomycetes. Nature, 202: 928-929.
CrossRefDirect Link - Lee, J.Y. and B.K. Hwang, 2002. Diversity of antifungal actinomycetes in various vegetative soils of Korea. Can. J. Microbiol., 48: 407-417.
Direct Link - Lo, C.W., N.S. Lai, H.Y. Cheah, N.K.I. Wong and C.C. Ho, 2002. Actinomycetes isolated from soil samples from the Crocker range Sabah. ASEAN Rev. Biodivers. Environ. Conserv., 9: 1-7.
Direct Link - Murad, H.A. and H.H. Azzaz, 2011. Microbial pectinases and ruminant nutrition. Res. J. Microbiol., 6: 246-269.
CrossRefDirect Link - Nanmori, T., T. Watanabe, R. Shinke, A. Kohno and Y. Kawamura, 1990. Purification and properties of thermostable xylanase and beta-xylosidase produced by a newly isolated Bacillus stearothermophilus strain. J. Bacteriol., 172: 6669-6672.
PubMedDirect Link - Oskay, A.M., T. Usame and A. Cem, 2004. Antibacterial activity of some actinomycetes isolated from farming soils of Turkey. Afr. J. Biotechnol., 3: 441-446.
Direct Link - Padmadhas, R.R., 2010. Isolation, characterisation and identification of novel actinomycetes collected from Western Ghats region of India. JPBMS, 1: 1-7.
Direct Link - Pridham, T.G., 1965. Color and streptomycetes: Report of an international workshop on determination of color of streptomycetes. Applied Microbiol., 13: 43-61.
Direct Link - Remya, M. and R. Vijayakumar, 2008. Isolation and characterization of marine antagonistic actinomycetes from west coast India. FACTA Univ. Ser. Med. Biol., 15: 13-19.
Direct Link - Saadoun, I., R. Rawashdeh, T. Dayeh, Q. Ababneh and A. Mahasneh, 2007. Isolation, characterization and screening for fiber hydrolytic enzymes-producing streptomycetes of Jordanian forest soils. Biotechnology, 6: 120-128.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Sivakumar, K., M.K. Sahu, T. Thangaradjou and L. Kannan, 2007. Research on marine actinobacteria in India. Indian J. Microbiol., 47: 186-196.
CrossRef - Sokal, R.R. and C.D. Michener, 1958. A statistical method for evaluating systematic relationships. Univ. Kansas Sci. Bull., 38: 1409-1438.
Direct Link - Subramaniyan, S. and P. Prema, 2002. Biotechnology of microbial xylanases: Enzymology, molecular biology and application. Crit. Rev. Biotechnol., 22: 33-64.
CrossRefDirect Link