
Norah A. Al-Enezi
Department of Biology, Science College, Dammam University, Dammam 31113, Saudi Arabia
Jameel M. Al-Khayri
Department of Agricultural Biotechnology, College of Agricultural and Food Sciences, King Faisal University, Al-Hassa 31982, Saudi Arabia
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 3 | Page No.: 146-153







ABSTRACT
Ionizing radiation induces physiological changes which activate stress-reaction mechanisms inciting proline accumulation in plants. The effects of radiation differ according to plant and dose parameters. The present study was conducted to determine the effects of different doses of X-rays on seedlings of date palm (Phoenix dactylifera L.). Seedlings collected from cv. Khalas were exposed to 0-1500 rad using a medical X-rays device. After exposure, the seedlings were grown in soil and growth was measured in terms of fresh and dry weight of roots and leaves. Accordingly, water content of root and leaf tissues was calculated. Proline content of leaf tissue was spectrophotometrically determined. The results showed that the X-rays dose affected proline accumulation and growth parameters of date palm seedlings. A direct relationship between the radiation dose and proline content was observed. Based on measurements of proline content, the minimum dose of X-rays that causes a significant stress reaction in date palm seedling is 100 rad. Exposure to X-rays caused noticeable increases in fresh and weight as well as water content of both leaf and root tissues. Significant changes in fresh and dry weights as well as water content of root and leaf tissues were observed with low doses ranging from 5 to 10 rad. Higher doses caused no additional significant changes in weights and water content. This study has shown that X-rays induce stress in date palm seedlings, as evident by proline accumulation. A noticeable increase in weight and water content occurred without noticeable phenotypic changes. Ionizing irradiation technology can facilitate stress-related physiological studies. Further exploration of this technology may prove beneficial for genetic improvement strategies of date palm.
PDF Abstract XML References Citation
Received: December 12, 2011;
Accepted: February 16, 2012;
Published: June 12, 2012
How to cite this article
Norah A. Al-Enezi and Jameel M. Al-Khayri, 2012. Effect of X-irradiation on Proline Accumulation, Growth and Water Content of Date Palm (Phoenix dactylifera L.) Seedlings. Journal of Biological Sciences, 12: 146-153.
DOI: 10.3923/jbs.2012.146.153
URL: https://scialert.net/abstract/?doi=jbs.2012.146.153
DOI: 10.3923/jbs.2012.146.153
URL: https://scialert.net/abstract/?doi=jbs.2012.146.153
INTRODUCTION
The impact of ionizing radiation on plant growth is largely deleterious and at high doses is lethal. Although at low doses growth stimulation may occur, the sensitivity of various plant species to different doses of ionizing radiation varies greatly (Sax, 1955). The behavior of numerous plant species under exposure to Gamma radiation have been extensively studied (Irfaq and Nawab, 2001, 2003; Ellafa et al., 2007; Norfadzrin et al., 2007; Songsri et al., 2011; Tuncer and Yanmaz, 2011). In comparison, studies on the effect of X-rays are relatively limited. Nonetheless, X-rays irradiation was shown to induce biological modifications that affected plant growth, cellular mechanisms and metabolism functions (Lea, 1947; Younis et al., 1962). Francis (1934) demonstrated retardation of fresh and dry weight yield of wheat (Triticum aestivum) irradiated seeds. Heavy doses of X-rays were shown to reduce seed viability, germination and growth in wheat and barley (Hordeum vulgare) (Froier and Gustafsson, 1941). Moreover, X-radiation can cause cytological changes and induce mutations (Goodspeed and Uber, 1939; Gustafsson, 1947; Shima and Ichikawa, 1994; El Araqi et al., 1997). Roy et al. (1972) reported reduced synthesis of DNA, RNA and protein was detected in X-radiation of cotyledons of stone pine (Pinus pinea). In broad bean (Vicia faba) transpiration rate and stomata opening were affected following irradiation resulting in decreased fresh and dry weights as well as water content of roots and leaves (Roy, 1974). Similarly, X-rays decreased fresh and dry weight and affected nucleic acid and protein metabolism in barley (Joshi and Ledoux, 1970). In addition, irradiation was found to induce changes in the concentration of cellular constituents like flavonoids in tissue extracts of several medicinal plant species (Al-Nimer Marwan and Abdul Lateef, 2009).
Ionizing radiations, including X-ray, are known to produce free radicals which can induce oxidative stress (Kumagai et al., 2003; Risom et al., 2003; Gudkov et al., 2010). This activates stress-reaction mechanisms in plant cells in an attempt to effectively scavenge the free radicals to relieve stress. It is well known that some soluble agents accumulate inside plants cells following exposure to stress factors. Proline is the most studied amino acid as a stress indicator (Claussen, 2005). It plays an essential role in free radicals resistance and contributes to cellular stress adjustments, protein stability and protection of enzymes, membranes and cytoplasm (Martinez et al., 1995; Gzik, 1996; Oncel et al., 2000; Patakas et al., 2002; Ozden et al., 2009). In addition, proline is a store for carbon and nitrogen which are necessary for growth during stress conditions (Singh et al., 1995). The amount of accumulated proline differs according to plant species and the intensity of stress factors (Delauney and Verma, 1993; Trotel et al., 1996; Ashraf and Orooj, 2006). Proline accumulation was studied as a metabolic response to salt stress and drought stress in several plant species (Kandpal et al., 1981; Hasegawa et al., 1984; Roy et al., 1993; Ismail et al., 1994; Hoque et al., 2007; Ueda, et al., 2007; Pongprayoon et al., 2008; Tatar and Gevrek, 2008; Amirjani, 2010; Ghotb Abadi et al., 2010). In date palm (Phoenix dactylifera L.) in vitro cultures, proline accumulation was observed to accumulate in response to stress conditions including salinity (Al-Khayri, 2002; Al-Bahrany and Al-Khayri, 2012) and drought (Al-Khayri and Al-Bahrany, 2004; Helaly and El-Hosieny, 2011).
In response to radiation, proline accumulation after exposure to ultraviolet (UV) rays was observed in cotton (Gossypium hirsutum) (Zhao et al., 2003), pea (Pisum sativum) (Katerova et al., 2009) and wheat (Balouchi et al., 2009). In date palm, treatments of seedlings with non-ionizing radiation like magnetic field were found to induce changes in proline content (Dhawi and Al-Khayri, 2008) as well as growth and water content (Dhawi and Al-Khayri, 2009) in addition to other physiological parameters (Dhawi and Al-Khayri, 2011).
In a recent study, Al-Enezi and Al-Khayri (2012) have shown that X-irradiation induced changes in the content of DNA, photosynthetic pigments and ions in date palm seedlings. No other studies addressing the effect of X-rays on date palm were encountered, despite the potential applications of X-irradiation in mutagenesis studies towards the genetic improvement of date palm. Moreover, literatures correlating proline accumulation in plants with X-rays dose appear lacking. Our objective was to evaluate the effect of various doses of X-rays on proline accumulation, water content and fresh and dry weights of date palm seedlings.
MATERIALS AND METHODS
Plant material: Seeds collected from female date palm trees of cv. Khalas were surface sterilized with 1% sodium hypochlorite for 5 min, soaked in water for 24 h at 37°C, then germinated over moist filter paper at the same temperature. Irradiation was applied and the seedlings were grown individually in a 20-cm plastic pot containing potting mix (1 soil:1 peat moss:1 vermiculite) and maintained in a greenhouse under natural light at 30-40°C with a relative humidity of approximately 50%. The seedlings were watered as needed to ensure that they were not stressed for moisture.
Irradiation treatments: At the age of 15 days, seedlings were placed in 9 cm Petri dishes and submerged in 10 mL of distilled water prior to exposure to X-rays. Using a therapeutic medical X-ray device (Clinac 23EX Linear Accelerator, Varian Medical Systems, USA), samples were exposed to different treatments (0, 5, 10, 25, 50, 100, 250, 500, 750, 1250 and 1500 rad) which are equivalent to (0, 0.05, 0.1, 0.5, 1, 2.5, 5, 7.5, 12.5 and 15 Gy). Each Petri dish contained 10 seedlings and 7 dishes were treated with each dose.
Estimation of proline: Amino acid proline was evaluated 3 months after exposure to X-rays according to the method described by Bates et al. (1973). Proline content was recorded at 520 nm using a UV/VIS spectrophotometer (Model V530, Jasco Inc., USA).
Plant measurements: The fresh weight of root and leaf samples was recorded after 3 months of irradiation. The dry weight was recorded after drying at 70 °C for 24 h in a drying vacuum oven (Model 3620, Lab Line Instruments Inc., USA). The percentage of water content for roots and leaves were calculated using the following equation:
Statistical analysis: The experiment was randomly designed with a single factor consisting of 11 levels with 7 replications. Data were subjected to analysis of variance (ANOVA) and the means were separated, where appropriate, using the Least Significant Difference (LSD) at 5%. Standard deviation for each treatment was also calculated.
RESULTS
Proline accumulation: Analysis of variance (ANOVA) revealed that the proline concentration in date palm leaves was significantly affected by the X-ray dose (Table 1).
| Table 1: | Analysis of variance of the proline accumulation in the leaves of date palm seedlings irradiated with various doses of X-ray |
| p-values less than 0.05 are significant | |
| Table 2: | Analysis of variance of tissue fresh and dry weights of leaves and roots of date palm seedlings irradiated with various doses of X-ray |
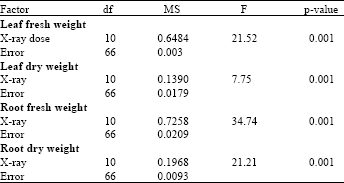 | |
| p-values less than 0.05 are significant | |
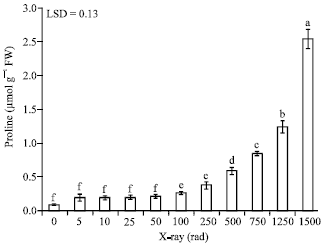 | |
| Fig. 1: | The impact of different doses of X-rays on proline accumulation in leaf tissue of date palm seedlings. Bars followed by different letters are significantly different |
Data indicated that proline accumulation increased in response to dose increases (Fig. 1). Incremental increases of X-ray dose ranging from 5 to 50 rad caused changes in proline content but the differences were not significant. In comparison to the control, proline accumulation increased significantly starting at 100 rad where proline content was almost 3 times that of the control. Increasing the dose to 250 rad, resulted in a concentration of proline 4 times that of the control; however, this treatment was not significantly different than the 100 rad treatment. Further increase of X-ray dose to 500, 750 and 1250 rad resulted in significantly more proline accumulation reaching 6, 8 and 12 times, respectively, that of the non-treated seedlings. A marked accumulation of proline occurred at 1500 rad which produced the highest concentration of proline, 2.55 μmol g-1, an equivalent to 28 times of that produced by the control seedlings, 0.09 μmol g-1. Based on these observed changes in proline content, it can be concluded that the minimum dose of X-ray that can incite a significant stress reaction in date palm seedling is 100 rad.
Weights of root and leaf tissues: Growth of date palm seedling expressed in fresh and dry weights of leaves and roots were significantly affected by X-ray dose, according to ANOVA (Table 2). Seedlings treated with as low as 5 rad of X-radiation exhibited a significant increase in root fresh weight (Fig. 2). Increasing the dose to 10 rad caused further significant increase in root fresh weight as compared to the preceding dose. Increasing the dose to 25 rad resulted in higher weight but this effect was not significantly different from the previous dose. Starting at 50 rad, increasing the X-ray dose caused no further 1.7 to 1.8 g, was obtained from seedlings treated with 10 to 1500 rad where dry weights reached 1.7 times that of the control seedlings, 1.3 g.
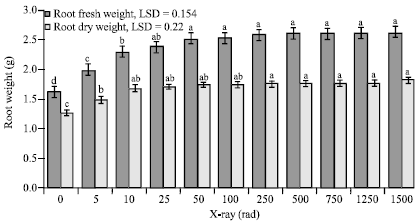 | |
| Fig. 2: | The impact of different doses of X-rays on root fresh and dry weights of date palm seedlings. Bars within a group followed by different letters are significantly different |
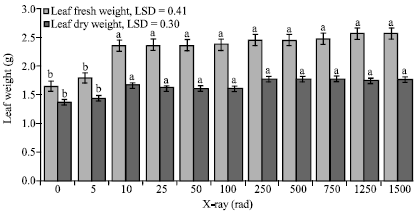 | |
| Fig. 3: | The impact of different doses of X-rays on leaf fresh and dry weights of date palm seedlings, Bars within a group followed by different letters are significantly different |
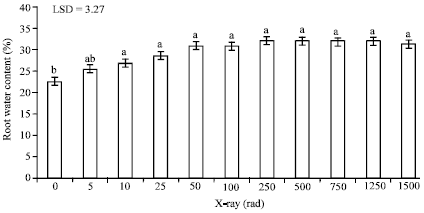 | |
| Fig. 4: | The impact of different doses of X-rays on root water content of date palm seedlings, Bars followed by different letters are significantly different |
In addition, fresh and dry weights of date palm leaf tissue were significantly affected by the X-ray dose. Generally, increasing the dose resulted in increased weights. At 5 rad, leaf fresh was slightly higher than the control but the difference was not significant (Fig. 3). Seedlings irradiated with 10 rad showed a significant increase in leaf fresh weight reaching 1.5 times that of the control. Increasing the dose beyond this level caused no further increase in weight.
Similarly, the minimum dose that induced a significant increase in dry weight was 10 rad (Fig. 3). No significant changes in weights were noticed at higher doses of radiation. The leaf dry weight increased from 1.4 g associated with the control to 1.7 g when a dose of 10 rad was applied. This increase is equivalent to 1.2 times that of the control.
Water content of root and leaf tissue: According to ANOVA (Table 3), water content of both leaf and root tissues was significantly affected by the radiation dose. Root water content of irradiated date palm seedlings showed incremental increase in response to increase the radiation doses.
| Table 3: | Analysis of variance of water content of leaves and roots of date palm seedlings irradiated with various doses of X-ray |
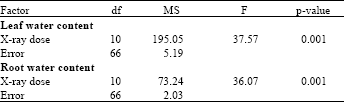 | |
| p-values less than 0.05 are significant | |
However, these increases ceased when doses higher than 50 rad were used (Fig. 4). The minimum X-radiation dose that elicited a significant increase in water content of roots was 10 rad. Water content of roots increased from 22.5% in the control seedlings to 30.9% obtained from seedlings irradiated with 50 rad. Doses higher than 50 rad gave a slight increase in water content but the difference was not significant.
Moreover, leaf water content of irradiated seedlings increased significantly with respect to the control. Exposure to 5 rad caused only a slight increase in water content; however, 10 rad resulted in a significant increase as compared to the control (Fig. 5).
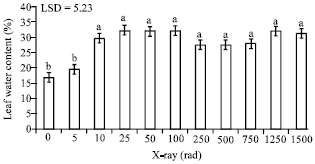 | |
| Fig. 5: | The impact of different doses of X-rays on leaf water content of date palm seedlings. Bars followed by different letters are significantly different |
Increasing the radiation dose to 25 rad resulted in the highest leaf water content, 32.2%, as compared to 16.8% associated with the control. This X-ray dose, however, gave a statistically similar response as 10 rad. When dose higher than 25 rad were used, changes in leaf water content were not significant.
DISCUSSION
In the present study date palm seedling were subjected to X-rays in the active phase (sprouting) as determined by Shull and Mitchell (1933), who reported that the effects of X-rays were more pronounced when seeds are subjected to radiation when in a sprouted condition. The most favorable effect on root fresh and dry weight and water content occurred with 50 rad. Whereas, in leaves the most favorable effect was obtained with 25 rad, This indicated that X-ray treatment effects differ among plant organs.
Treatments with X-rays doses from 5 to 1500 rad induced proline accumulation which became greater with dose increases. Weight decrease caused by X-ray exposures could be due to the accumulation of proline which was significantly higher and increased with increasing dose rates (5-155 rad). The accumulation of amino acid causes a decrease of water content and dry-fresh weight (Gzik, 1996; Claussen, 2005). Possibly proline accumulation in date palm exposed to X-rays was due to the role of proline in the resistance of free radicals that occur in plants exposed to X-rays. We conclude that proline accumulation in date palm leaves increased significantly, affected by oxidative stress resulting from X-ray stress. This effect resembles proline accumulation in date palm in vitro cultures subjected to salt stress (Al-Khayri, 2002) and drought stress (Al-Khayri and Al-Bahrany, 2004).
Likewise, Dhawi and Al-Khayri (2008) demonstrated that at the lowest intensity, 10 mT, of Static Magnetic Field (SMF), proline concentration increased in response to longer exposure durations, reaching a maximum at 240 min. Beyond that duration, a reduction in proline concentration occurred. In contrast, at higher doses, 50 and 100 mT, proline concentration decreased as the exposure duration was increased. Alternating Magnetic Field (AMF) significantly reduced proline concentration after as little as a 1 min. exposure.
In a previous study with date palm seedlings, Al-Enezi and Al-Khayri (2012) have demonstrated a significant increase in DNA content in response to 5 rad reaching the highest concentration at 25 rad. Similarly, the highest ions content was observed at 25 rad. A dose of 5 rad caused a significant increased sodium, potassium and phosphorus ions. A higher dose, 10 rad, was necessary to increase the content of calcium and magnesium ions. Conversely, total photosynthetic pigments started to diminish at 5 rad and continued to decrease with dose reaching the lowest level at 1500 rad.
Growth parameters studied were also influenced by radiation. In a previous study with magnetic fields, Dhawi and Al-Khayri (2009) observed that leaf fresh and dry weight and water content increased significantly in response to SMF treatments. Similarly, root fresh weight and water increased significantly; however root dry weight increases were insignificant. The same study showed that treatments with AMF significantly increased all the plant growth parameters measured.
The current study has shown that low doses of X-ray affect date palm seedlings positively. In a similar study, low doses of 150 rad X-ray stimulated growth of two cultivars of winter wheat (Triticum aestivum) and increased yield, with increasing doses ranging from 150 to 2000 rad (Erickson et al., 1979). A study on salad rocket plant (Eruca vesicaria ssp. sativa) showed that X-ray leads to higher growth with respect to control samples (Sakuma and Nakayama, 2006).
Moreover, Wort (1941) reported that the greatest fresh and dry weights resulted at 76 rad. Doses of X-radiation from 76 to 114 rad accelerated heading and flowering of plants. Height and weight of winter wheat seedlings were increased considerably by all doses of X-rays used; the maximum occurring when the dosage was 114 rad. An increase in weight of okra (Abelmoschus esculentus) was observed in samples treated with X-rays doses above 8 kR (Rao and Rao, 1978). Similarly, X-ray treatments stimulated plant growth in wheat, corn (Zea mays), oats (Avena sativa) and sunflower (Helianthus annuus) (Shull and Mitchell, 1933). High doses of X-rays affected growth of date palm seedlings negatively. This is analogous to Van Huystee and Cherry (1967) finding that radiation levels of 750 krad and higher completely inhibited peanut (Arachis hypogaea) seed growth and germination. A study by Roy (1974) showed that Vicia faba leaf growth was reduced; whereas, water loss was reduced through stomata openings under X-ray treatment resulting in a higher water content. In addition, the treatment of barley plants by X-ray led to decrease fresh and dry weight (Joshi and Ledoux, 1970). A study by Francis (1934), on wheat seedlings showed that high doses of X-rays (565-13560 rad) caused retardation of fresh weight and dry weight production of the growing parts of the seedlings and retardation of linear growth of the coleoptile, leaf, primary root and the lateral roots. On the contrary, a high dose has a stimulatory effect on some plants. Younis et al. (1962), observed increase of growth, flowering and fruiting in Egyptian cotton seeds with X-radiation doses of 250, 500 and 10000 rad.
CONCLUSION
Irradiating date palm seedlings with X-ray caused a significant accumulation of proline and increased water content and weight of both roots and leaves. Despite the doses used, the seedlings exhibited normal growth appearance. The accumulation of proline is an indication of cellular activity to counter the effect of radiation-induced stress which is known to be a consequence of free radicals activity. Irradiation is a powerful tool for genetic improvement of agricultural plants. In date palm, irradiating in vitro cultures with Gamma radiation proved extremely effective to induce mutants resistant to Bayoud disease, a major disease of date palm, caused by the soil fungus Fusarium oxysporum f. sp. albedinis (Jain, 2007, 2010). Nonetheless, the utilization of irradiation technology in date palm improvement remains limited because of the lack of relevant studies. The current study gives the first insight into the response of date palm to X-radiation. Information presented here should facilitate future studies aimed at developing strategies for date palm genetic improvement through mutation induction and selection techniques.
REFERENCES
- Al-Khayri, J.M., 2002. Growth, proline accumulation and ion content in sodium chloride-stressed callus of date palm. In Vitro Cell. Dev. Biol.-Plant, 38: 79-82.
CrossRefDirect Link - Al-Khayri, J.M. and A.M. Al-Bahrany, 2004. Growth, water content and proline accumulation in drought-stressed callus of date palm. Biol. Plant, 48: 105-108.
CrossRefDirect Link - Al-Nimer Marwan, S.M. and Z.W. Abdul Lateef, 2009. X-rays radiation directly produced favorable and harmful effects on the constituents of different medicinal plants. Pharmacogn. Res., 1: 331-335.
Direct Link - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - Ashraf, M. and A. Orooj, 2006. Salt stress effects on growth, ion accumulation and seed oil concentration in an arid zone traditional medicinal plant ajwain (Trachyspermum ammi [L.] Sprague). J. Arid Environ., 64: 209-220.
CrossRefDirect Link - Balouchi, H.R., S. Sanavy, Y. Emam and A. Dolatabadian, 2009. UV radiation, elevated CO2 and water stress effect on growth and photosynthetic characteristics in durum wheat. Plant Soil Environ., 55: 443-453.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Claussen, W., 2005. Proline as a measure of stress in tomato plants. Plant Sci., 168: 241-248.
CrossRefDirect Link - Delauney, A.J. and D.P.S. Verma, 1993. Proline biosynthesis and osmoregulation in plants. Plant J., 4: 215-223.
CrossRef - El Araqi, K.T., J. Gilot-Delhalle, I. Roelandts, G. Weber, R. Deltova and J. Moutschen, 1997. Modified fast rejoining of chromosomes after fractionated X-ray exposures with iron and copper sulfate pretreatments related to uptake detected by PIXE analysis in Nigella damascena seeds. Environ. Exp. Bot., 38: 87-97.
CrossRefDirect Link - Ellafa, K., O.H. Ahmed, S. Shaharudin and D.A. Rahman, 2007. Gamma radio sensitivity study on snap bean (Phaseolus vulgaris). Int. J. Agric. Res., 2: 844-848.
Direct Link - Erickson, P.I., M.B. Kirkham and G.B. Adjei, 1979. Water relations, growth and yield of tall and short wheat cultivars irradiated with X-rays. Environ. Exp. Bot., 19: 349-356.
CrossRefDirect Link - Francis, D.S., 1934. The effects of X-rays on growth and respiration of wheat seedlings. Bull. Torrey Bot. Club, 61: 119-153.
Direct Link - Abadi, F.S.G., M. Mostafavi, A. Eboutalebi, S. Samavat and A. Ebadi, 2010. Biomass accumulation and proline content of six citrus rootstocks as influenced by long-term salinity. Res. J. Environ. Sci., 4: 158-165.
CrossRefDirect Link - Goodspeed, T.H. and F.M. Uber, 1939. Radiation and plant cytogenetics. Bot. Rev., 5: 1-48.
Direct Link - Gudkov, S.V., S.A. Garmash, I.N. Shtarkman, A.V. Chernikov, O.E. Karp and V.I. Bruskov, 2010. Long-lived protein radicals induced by X-ray irradiation are the source of reactive oxygen species in aqueous medium. Dokl. Biochem. Biophys., 430: 1-4.
CrossRefDirect Link - Gzik, A., 1996. Accumulation of proline and pattern of α-amino acids in sugar beet plants in response to osmotic, water and salt stress. Environ. Exp. Bot., 36: 29-38.
CrossRefDirect Link - Helaly, M.N.M. and A.M.H. EL-Hosieny, 2011. In vitro selection and photosynthetic characterization of date palm regenerated seedling as affected by water stress. Am. J. Plant Physiol., 6: 126-143.
CrossRef - Hoque, M.A., E. Okuma, M.N.A. Banu, Y. Nakamura, Y. Shimoishi and Y. Murata, 2007. Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities. J. Plant Physiol., 164: 553-561.
CrossRefPubMedDirect Link - Irfaq, M. and K. Nawab, 2001. Effect of γ irradiation on some morphological characteristics of three wheat (Triticum aestivum L.) cultivars. J. Biol. Sci., 1: 935-937.
CrossRefDirect Link - Irfaq, M. and K. Nawab, 2003. A study to determine the proper dose of gamma radiation for inducing beneficial genetic variability in bread wheat (Triticum aestivum L.). Asian J. Plant Sci., 2: 999-1003.
CrossRefDirect Link - Ismail, M.R., S.W. Burrage and M.A. Aziz, 1994. Growth, plant water relations, photosynthesis rate and accumulation of proline in young carambola plants in relations to water stress. Sci. Hortic., 60: 101-114.
Direct Link - Jain, S.M., 2007. Recent advances in date palm tissue culture and mutagenesis. Acta Hortic., 736: 205-211.
Direct Link - Jain, S.M., 2010. Mutagenesis in crop improvement under the climate change. Romania Biotech. Lett., 15: 88-106.
Direct Link - Joshi, R.K. and L. Ledoux, 1970. Influence of X-irradiation and seed-moisture on nucleic-acid and protein metabolism in barley. Rad. Bot., 10: 437-443.
CrossRefDirect Link - Kandpal, R.P., C.S. Vaidyanathan, M.U. Kumar, K.S.K. Sastry and N.A. Rao, 1981. Alterations in the activities of the enzymes of proline metabolism in Ragi (Eleusine coracana) leaves during water stress. J. Biosci., 3: 361-370.
CrossRefDirect Link - Katerova, Z., S. Ivanov, S. Mapelli and V. Alexieva, 2009. Phenols, proline and low-molecular thiol levels in pea (Pisum sativum) plants respond differently toward prolonged exposure to ultraviolet-B and ultraviolet-C radiations. Acta Physiol. Plant., 31: 111-117.
CrossRefDirect Link - Kumagai, J., K. Masui, Y. Itagaki, M. Shiotani, S. Kodama, M. Watanabe and T. Miyazaki, 2003. Long-lived mutagenic radicals induced in mammalian cells by ionizing radiation are mainly localized to proteins. Radiat. Res., 160: 95-102.
PubMed - Martinez, C.A., C. Guerrero and U. Moreno, 1995. Diurnal fluctuations of carbon exchange rate, proline content and osmotic potential in two water-stress potato hybrids. R. Bras. Fisiol. Veg., 7: 27-33.
Direct Link - Norfadzrin, F., O.H. Ahmed, S. Shaharudin and D.A. Rahman, 2007. A preliminary study on γ radiosensitivity of tomato (Lycopersicon esculentum) and Okra (Abelmoschus esculentus). Int. J. Agric. Res., 2: 620-625.
CrossRefDirect Link - Oncel, I., Y. Keles and A.S. Ustun, 2000. Interactive effects of temperature and heavy metals stress on the growth and some biochemical compounds in wheat seedlings. Environ. Pollut., 107: 315-320.
Direct Link - Ozden, M., U. Demirel and A. Kahraman, 2009. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Hort., 119: 163-168.
CrossRef - Patakas, A., N. Nikolaou, E. Zioziou, K. Radoglou and B. Noitsakis, 2002. The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines. Plant Sci., 163: 361-367.
CrossRefDirect Link - Pongprayoon, W., S. Cha-Um, A. Pichakum and C. Kirdmanee, 2008. Proline profiles in aromatic rice cultivars photoautotrophically grown in responses to salt stress. Int. J. Bot., 4: 276-282.
CrossRefDirect Link - Rao, S. and D. Rao, 1978. Effect of irradiation on physiological and morphological variability's in Abelmoschus esculentus (L.) Moench. Proc.: Plant Sci., 87: 129-133.
Direct Link - Risom, L., P. Moller, U. Vogel, P.E. Kristjansen and S. Loft, 2003. X-ray-induced oxidative stress: DNA damage and gene expression of HO-1, ERCC1 and OGG1 in mouse lung. Free Radical. Res., 37: 957-966.
PubMedDirect Link - Roy, D., N. Basu, A. Bhunia and S.K. Banerjee, 1993. Counteraction of exogenous L-proline with NaCl in salt-sensitive cultivar of rice. Biol. Plant., 35: 69-72.
CrossRefDirect Link - Roy, R.M., 1974. Transpiration and stomatal opening of X-irradiated broad bean seedlings. Rad. Bot., 14: 179-184.
Direct Link - Roy, R.M., B. Donini and A. Brunori, 1972. Biochemical and cytological studies on developing cotyledons of Pinus pinea following X-ray irradiation of dry seeds. Radiation Bot., 12: 249-260.
CrossRef - Sakuma, A. and M. Nakayama, 2006. Radiation hormesis using an X-ray radiography device (the fourth report): Radiation hormesis of salad rocket. Radioisotopes, 55: 687-690.
Direct Link - Sax, K., 1955. The effect of ionizing radiation on plant growth. Am. J. Bot., 42: 360-364.
Direct Link - Shima, N. and S. Ichikawa, 1994. Synergisms detected among methyl methanesulfonate, ethyl methanesulfonate and X-rays in inducing somatic mutations in the stamen hairs of Tradescantia clone BNL 4430. Environ. Exp. Bot., 34: 393-408.
Direct Link - Shull, C.A. and J.W. Mitchell, 1933. Simulative effects of X-rays on plant growth. Plant. Physiol., 8: 287-296.
Direct Link - Singh, V.P., S.R. Voleti and C.N. Rao, 1995. Distribution pattern of free proline in rice (Oryza sativa L.) grown under low light irradiance. J. Agron. Crop Sci., 175: 207-209.
Direct Link - Songsri, P., B. Suriharn, J. Sanitchon, S. Srisawangwong and T. Kesmala, 2011. Effects of gamma radiation on germination and growth characteristics of physic nut (Jatropha curcas L.). J. Biol. Sci., 11: 268-274.
CrossRefDirect Link - Tatar, O. and M.N. Gevrek, 2008. Influence of water stress on proline accumulation, lipid peroxidation and water content of wheat. Asian J. Plant Sci., 7: 409-412.
CrossRefDirect Link - Trotel, P., A. Bouchereau, M.F. Niogret and F. Larher, 1996. The fate of osmo-accumulated proline in leaf discs of rape (Brassica napus L.) incubated in a medium of low osmolarity. Plant Sci., 118: 31-45.
CrossRef - Ueda, A., Y. Yamamoto-Yamane and T. Takabe, 2007. Salt stress enhances proline utilization in the apical region of barley roots. Biochem. Biophys. Res. Commun., 335: 61-66.
Direct Link - Van Huystee, R. and J.H. Cherry, 1967. Effect of X-irradiation and post irradiation storage of peanut seed on nucleic acid metabolism in cotyledons. Rad. Bot., 7: 217-223.
Direct Link - Wort, J.D., 1941. X-ray effects on the growth and reproduction of wheat. Plant Physiol., 16: 373-383.
PubMedDirect Link - Younis, A.E., M.A. Hammouda and A.T. Hegazi, 1962. Effect of X-radiation of soaked cotton seeds upon growth, fruiting and yield. Plant Soil, 17: 131-133.
CrossRefDirect Link - Zhao, D., K.R. Reddy, V.G. Kakani, J.J. Read and J.H. Sullivan, 2003. Growth and physiological responses of cotton (Gossypium hirsutum L.) to elevated carbondioxide and ultraviolet-B radiation under controlled environmental conditions. Plant Cell Environ., 26: 771-782.
CrossRefDirect Link