
Satadal Chakrabarty
Central Sericultural Research and Training Institute, Berhampore (W.B.), 742101, India
B. Manna
Parasitology Research Unit, The University of Calcutta, 35, Ballygunge Circular Road, Kolkata 700 019, India
A.K. Saha
Central Sericultural Research and Training Institute, Berhampore (W.B.), 742101, India
B.B. Bindroo
Central Sericultural Research and Training Institute, Berhampore (W.B.), 742101, India
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 1 | Page No.: 1-14







ABSTRACT
The spore production of Nosema sp. in mulberry silkworm, Bombyx mori and non-mulberry silkworms, tasar, Antheraea mylitta D.; Eri, Philosamia ricini B. and muga, Antheraea assama Ww. were examined and found that spore multiplication in the host followed a logistic pattern of development. Inoculum concentrations of Nosema sp. during various seasons is the most important factor for the development of pebrine disease in mulberry and non-mulberry silkworms. Mean larval mortality in mulberry silkworm was high (90.00%) when larvae inoculated with low concentration (1.52x106 spores mL-1) of N. bombycis and it was low (66.48%) during March-April and gradually decreased during December-January (28.85%) and May-June (25.21%), respectively. The spore multiplication reaches its peak maximally in moth stage (2.67x108 spores mL-1). However, the rate of multiplication of pathogen decreased (2.0x108 and 1.4x108 spores mL-1) with the higher inoculum concentrations of pathogen (1.52x108 and 1.52x107 spores mL-1) respectively. The high larval mortality (85.00%) was recorded in A. mylitta during cross-infection with high dose (1.52x108 spores mL-1) of N. assamensis as compared to same dose of N. bombycis and N. mylitta and mean seasonal mortality was high during August-September (73.95%) followed by October-November (63.55%). Generally, larval mortality in A.mylitta was recorded high (61.44-85.00%) irrespective of variation in pathogen, inoculum concentrations and seasons. Maximum larval mortality was recorded in 3rd instar (39.44%) and 4th instar (20.55%), respectively. Multiplication of N. mylitta in moth stage was high (4.0x109 spores mL-1) in comparison to any stage of larva or pupa, when inoculation was done at 1st instar. Mortality in eri larvae with the infection of N. ricini and cross-infection with N. bombycis, N. mylitta, N. assamensis were not observed even after 3rd successive inoculation with high doses (1.52x108 spores mL-1) of above pathogens. Mortality percentage was found always high (52.00-100%) in muga larvae infected by N. bombycis, N. mylitta, N. ricini and N. assamensis during August-September in comparison to October-November.
PDF Abstract XML References Citation
Received: July 11, 2011;
Accepted: October 04, 2011;
Published: January 20, 2012
How to cite this article
Satadal Chakrabarty, B. Manna, A.K. Saha and B.B. Bindroo, 2012. Comparative Study on Effect Different Types of Nosema sp.
(Microsporidia: Nosematidae) on Mulberry and Vanya Silkworms. Journal of Biological Sciences, 12: 1-14.
DOI: 10.3923/jbs.2012.1.14
URL: https://scialert.net/abstract/?doi=jbs.2012.1.14
DOI: 10.3923/jbs.2012.1.14
URL: https://scialert.net/abstract/?doi=jbs.2012.1.14
INTRODUCTION
Silk and silk products have been associated with luxury and traditionally been expensive. Natural silk is produced in very few countries of world. China and India are the major silk producers among the 58 silk producing countries in the world. China contributed (1, 30, 000 MT) nearly 85% of the total silk production in the world during 2006-07 (1, 53, 942 MT). India produced 13.1% of silk. India has a unique distinction of being the only country in the world producing four varieties of silk and produced 16,525 MT of mulberry, 350 MT of tasar, 1485 MT of eri and 115 MT of muga silk during 2006-07 (Anonymous, 2008).
Nevertheless, the production is comparatively low due to various reasons of which disease is noteworthy. All silkworms suffer from various diseases like Pebrine (Protozoa: Microsporida), Flacherie (Bacteria), Grasserie (Virus) and Muscardine (Fungus). Thus these pathogens have a great role in deterioration of sericulture industry. Further, most of the commercially silkworm races exploited in India are prone to microsporidian infection and silkworm races are reared in India since decades without any major change. As a result, they loose their tolerance against diseases (Veeranna, 1999). B. mori is highly susceptible to diseases. Crop loss due to infection with various diseases was seen up to 20-40% (Janakiraman, 1961; Hanumanappa, 1968; Chitra et al., 1975; Veeranna, 1999). About 30% crop loss is attributed due to pebrine disease with occasional crop failure (Nataraju et al., 2005).
Deviah and Krishnaswamy (1975) observed the seasonal incidence of pebrine disease in B. mori. Sengupta and Griyaghey (1981) found that mortality rate is directly proportional to the amount of spore administer. Patil (1993) opined that 3rd and 4th stage tasar larvae are more susceptible to pathogen infection and maximum mortality was recorded during late age and spinning time. Only 100 spores of N. bombycis is required by 5th instar larvae of B. mori to cause disease (Iwano and Ishihara, 1981).
Mariswamy (1977) reported that temperature and humidity plays a vital role in development of disease of muga and eri silkworms. The incidence of pebrine is more during summer season followed by autumn and spring in temperate countries (Jolly, 1986). Among environmental factors, temperature plays a major role on growth and productivity in silkworm (Benchamin and Jolly, 1986). Late age silkworm prefers relatively lower temperature than young age (Krishnaswami, 1994). Slight fluctuation of temperature during different stages of larval development is more favourable for growth and development of pathogen than constant temperature (Anonymous, 1975). Mishra et al. (1992) reported the comparative effect of larval mortality on tasar silkworm, A. mylitta due to pebrine infection during three rearing seasons, rain, autumn and winter. Griyaghey and Krishnan (1976) studied on the thermo-control of microsporidiosis of the tropical tasar silkworm, A. mylitta. Sing et al. (1976) described the influence of altitude and climate on rearing of the oak tasar silkworm, A. proylei, in Manipur. Eri silkworm is comparatively more resistant to disease and tolerant of high fluctuations of temperature and humidity (Thangavelu, 1989). Eri silkworm is hardy and can withstand the extreme climatic conditions (Prasad and Saha, 1992).The muga silkworm is susceptible to various diseases resulting in heavy crop loss (Thangavelu and Sahu, 1983).
Multiplication of pathogen depends upon the age of silkworm and time dependent and other indirect factors (Sloter et al., 1989). Rainfall, temperature and relative humidity are other abiotic factors responsible for multiplication of pathogen (Baruah et al., 1998; Chakrabarti and Manna, 2008a). Transmission of disease is dependent on the intensity of mother moth suffers (Griyaghey and Sengupta, 1989). Yasonga et al. (1991) has studies infection and development of Nosema sp. NIS M11 in a lepidopteran cell line and they found that life cycle and spore production is different in unsuitable medium. Hayasaka et al. (1993) studied on infection and proliferation of microsporidians pathogenic to the silkworm, Antheraea pernyi in lepidopteran cell lines. Wilson and Sohi (1977) studied the effect of temperature on healthy and microsporidia infected continuous cultures of hemocytes of Malacosoma distriae.
Chakrabarti and Manna (2006) described three species of Nosema infecting vanya silkworms prevalent in Eastern and North-Eastern India and named them as Nosema ricini from P. ricini, N. assamensis from A. assamensis and N. mylitta from A. mylitta based on ten major morphological characters of the spores under electron microscopy. N. mylitta is cross-infected to B. mori. N. bombycis and N. assamensis are cross-infected to A. mylitta. All the pathogens, N. bombycis, N. mylitta and N. ricini are cross-infected to A. assamensis. But, N. bombycis, N. mylitta and N. assamensis are not cross-infected to P. ricini (Chakrabarti and Manna, 2008b, 2009). Nosema sp. which affecting Philosamia cyanthia ricini and Antheraea pernyi do not infect mulberry silkworm (Jolly, 1986).
From the foregoing literature, it is observed that most of the workers studied in their respective field but a concrete picture of relationship between multiplication of pebrine spores with silkworm mortality in respect of seasons, spore concentration, infection, cross-infection and transovarian transmission in mulberry and vanya silk in eastern and north eastern region is not available.
Therefore, we have undertaken detail study on the relationship of inoculum concentration of Nosema sp. with larval mortality and multiplication of pebrine spore during infection, cross-infection and transovarian transmission in mulberry and vanya silkworm in respect of various seasons for pebrine disease development in eastern and north eastern region.
MATERIALS AND METHODS
Mulberry silkworm: The silkworm (Nistari breed) rearing was conducted as per standard procedure (Dandin et al., 2001). The period, Nov-Dec (season-1) and Feb-Mar (season-2) were considered as favourable seasons and period May (season-3) as unfavourable. Temperature, 25-28, 28.5-34.5 and 20-40.5°C and Relative Humidity of 65-72, 55-81 and 64-90.5% were maintained during season-1, 2 and 3, respectively with 12L+12D photoperiodic condition. Twelve replications each with 3rd stage ‘0’ h 60 larvae were considered for experiment.
Infected mulberry, tasar, eri and muga silkworm pupae were obtained from the silk growers of Berhampore (West Bengal), Baripada (Odissa), Boko (Assam) and Sibsagar (Assam) respectively and used in the present study. The spores were isolated from the infected pupae and purified by centrifugation at 3000 rpm for 10 min using percoll cushions (PVP coated silica particles, Sigma chemicals Co., USA) following standard procedure (Sato and Watanabe, 1986). After centrifugation, the spores were suspended in 0.65% NaCl and stored at 4°C. To obtain fresh spores, 3rd instar ‘0’ h mulberry, tasar, eri and muga larvae were per orally inoculated through the leaf with respective Nosema sp. Spores were counted using a Neubauer haemocytometer under light microscope (x600) and determined the inoculums concentration according to Undeen (1997).
N. bombycis, N. mylitia, N. ricini and N. assamensis were propagated in their respective primary hosts. Spores were isolated from pool sample and purified from moths and made in three different concentrations (1.5x106, 1.5x107 and 1.5x108 spore mL-1). Three replications each with 60 larvae were maintained for each concentration. Each dose of isolated Nosema in aqueous suspension (1.5 mL) was thoroughly mixed with freshly prepared three mulberry leaf dishes (28.3 cm2) and dried and allowed to feed to the larvae of each replication for a period of 6 h. The mulberry leaf dishes smeared with distilled water were fed to the larvae of healthy control group.
Mortality due to pebrine was confirmed in dead larvae by microscopic examination. Spores were isolated from pooled sample and purified and were counted. All the data were statistically analyzed by using in INDOSTAT software package version (Indostat Services, Hydrabad, India).
For study of transovarian transmission study, infected male and female moths of first generation were allowed for coupling. Eggs laid by infected female moths were allowed for hatching. Newly hatched larvae were reared on mulberry leaves following standard rearing techniques (Dandin et al., 2001). Dead larvae of different stages, pupae and moths were collected. Pebrine spores were isolated from pool sample and counted.
Tasar silkworm: Ten disease free layings of tasar silkworm (A. mylitta D., Race-Daba) were collected from Basic Seed Multiplication and Training Centre, Central Silk Board, Patelnagar, Dist-Birbhum, West Bengal, India. Few eggs were crushed with mortar and pestle in sterilized water and examined under microscope to confirm further that the eggs were free from disease. Healthy control larvae were reared separately.
Trivoltine tasar silkworm, A. mylitta were reared in indoor under laboratory condition on a diet of fresh Arjuna twigs (Terminalea arjuna) dipped into water in mud pitcher at 26-33°C and 93.5-71.0% R.H and 12L+12D photoperiodic condition and reared during Aug-Sep, 2001. Freshly collected 1st instar larvae at ‘0’ h (1st day 1st instar) were considered for experiment and were fed on fresh Arjuna leaves smeared with Nosema sp. of mulberry, tasar, eri and muga silkworms. The procedure involves, dipping of leaf in 1.5 mL of spore suspension, drying and then allowing the larvae to feed through leaf for a period of 6 h. The Arjuna leaves dipped with distilled water were fed to the tasar silkworm of healthy control group. For the next season, procedure involved as the in case of previous rearing, inoculation, purification etc. was followed. Rearing was conducted during Sep-Oct, 02 under 19-33°C and 62-94% R.H. Mortality was recorded and dead larvae were examined under light microscope to confirm mortality due to pebrine disease. Spores were isolated from pooled sample and purified and were counted.
For study of transovarian transmission study, infected male and female moths were allowed for coupling and grainage was conducted. Eggs laid by infected female moths were allowed for hatching. Newly hatched larvae were allowed for feeding Arjuna leaves following standard techniques. The dead larvae of different stages in second generation were collected for study of transovarian transmission and pebrine spores were counted.
Eri silkworm: Ten disease free layings of Philosamia ricini collected from Eri Silkworm Seed Project Office, Central Silk Board, Mirza, Kamrup district, Assam, India. Fifty eggs were crushed in morter and pestle and tested as a sample under compound microscope to confirm that the eggs were free from disease.
After hatching, the batches of 180 larvae were again divided into 3 of 60 larvae in each replication. 1st instar ‘0’ h larvae (1st day 1st instar) were fed fresh Castor (Ricinus communis) leaves, smeared with pathogens, N. bombycis, N. mylitta, N. ricini and N. assamensis suspension containing 1.52x108 spore. mL-1. Briefly, the procedure involves as in other cases. All the larvae were reared in indoor by dala/rearing tray. Rearing was conducted during Mar-Apr, 2003 (20 days). All dead larvae after 4 days of post inoculation were examined under microscope for confirmation of mortality due to pebrine infection.
Muga silkworm: Fifteen disease free layings of muga silkworm (A. assamensis Ww., Multivoltine, Semi-domesticated) were collected from Regional Muga Research Station, Central Silk Board, Boko, Dist-Kamrup, Assam, India and Asstt. Director, Research Extension Centre, Coochbehar. Few eggs were crushed with mortar and pestle in sterilized water and examined under microscope to confirm further that the eggs were free from disease. All the larvae were reared in indoor on som and soalu leaves. Rearing was conducted during Aug-Sep, 2002 (32 days) (season-1) and Sep-Oct, 2002 (34 days) (season-2). Eggs were hatched and rearing was conducted in between 19-33°C and 62-94% R.H.
All dead larvae after 4 days of post inoculation were examined under microscope for confirmation of mortality due to pebrine infection. One hundred selected larvae were reared as healthy control. Briefly, the procedure involves as in other cases.
Comparative study of effect of different concentrations of pathogens responsible for larval mortality and multiplication of pathogens during different stages of host and transovarial transmission in different seasons.
B. mori, A. mylitta, P. ricini and A. assamensis were taken as host and three different concentrations, 1.52x108, 1.52x107 and 1.52x106 spore mL-1. of N. bombycis, N. mylitta, N. ricini and N. assamensis were taken as pathogens. The procedure involves for inoculation of pathogens, recording mortality %, spore multiplication, counting of spores was followed as in previous cases. All dead larvae after 4 days of post inoculation were examined under microscope for confirmation of mortality due to pebrine infection and mortality % of larva were recorded. The experiment was conducted in different seasons. Then spores were purified and isolated from all stages of dead larva, pupae and moth and counted under haemocytometer over Thoma-Zaiss counting slides.
RESULTS
Mulberry silkworm: Range of larval mortality was highly significant (p<0.01) in all cross-infected larvae. Mortality was recorded significantly higher (76.11±0.61%) in larvae cross-infected with high inoculum concentration, 1.5x108 spore mL-1 of N. mylitta during season-1 in first generation. However, N. mylitta with higher inoculum concentration (1.52x108 spore mL-1) was very effective for higher mortality (89-98.6%) in all the three seasons in second generation. It was observed that at least minimum inoculum load of 1.5x106 spore mL-1 was required for significantly high larval mortality in first generation but below that there was no effect on mortality in all batches. In the present observation, multiplication of spore was highly significant (p<0.01) in all cross-infected larvae in second generation. Multiplications of spore were observed highest in moth (2.0x108-2.1x108 spore mL-1) and pupal stage (1.76x108-1.83x108 spore mL-1) of first and second generation respectively, when N. mylitta infected with higher inoculum concentration (1.5x108 spore mL-1). N. mylitta transovarially transmitted with 100 % efficiency during second generation.
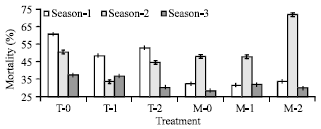 | |
| Fig. 1: | Mortality % of Bombyx mori in different concentrations of Nosema sp. (T = N. mylitta, M= N. bombycis, Treatment 0 = 1.52x108 spore mL-1. Treatment 1 = 1.52x107 spore mL-1, Treatment 2 = 1.52x106 spore mL-1, S1, S2 and S3= Season-1 (Nov-Dec-Winter ), 2 ( Feb-Mar-Spring) and 3 (May-Summer), Bar = Standard error of mean |
More than 98% mortality were recorded when larvae transovarially infected with higher inoculum load (1.5x108 spore mL-1). Mortality of 5th stage larva was recorded in treated batches and no mortality was recorded in control batches. In December to January (Season-1) mortality rate of 5th stage larva was 76.106% in mulberry larva inoculated by Nosema mylitta in the highest inoculums (1.52x108 spore mL-1) concentration (T-O). Where as mortality was 90.00 % in mulberry larva inoculated by the lowest dose (1.52x106 spores mL-1) of N. bombycis (M-2),during March-April (Season-2) (Fig. 1). However, highest dose (1.52x108 spores mL-1) of N. mylitta (T-0) caused to highest mean larval mortality (57.582 %). Where as, lower doses (1.52x106 and 1.52x107 spore mL-1) of N. mylitta (T-1 and T-2) caused to lower mean mortality (46.11 and 40.92 %), respectively. The highest mortality was recorded 965.636%) during December-January (Season-1) in all the batches of mulberry silkworm infected by Nosema mylitta and mean larval mortality% was gradually decreasing from December-January (Season-1) (65.636 %) to February-March (Season-2) (46.29%) to 3 May (Season-3) (32.95%) (Fig. 1).
Maximum multiplication of pathogen was observed in later days of 5th stage larva and in pupa and adult moth. Maximum multiplication (2 spore mL-1) of pathogen was observed in moth stages of mulberry silkworm, when it was cross-infected by highest dosage (1.52x108 spore mL-1) of N. mylitta collected from tasar silkworm [T-O]. Gradually the rate of multiplication of spore decreased (1.35x108 and 1.20x108 spore mL-1) with the decreasing inoculum concentrations of pathogen (T-1, 1.52x107 and T-2, 1.52x106 spore mL-1), respectively. However, when mulberry silkworm was infected by Nosema bombycis with lower inoculums concentration (M-2, 1.52x106 spores mL-1) multiplied maximally (2.67x108 spore mL-1) in moth stage.
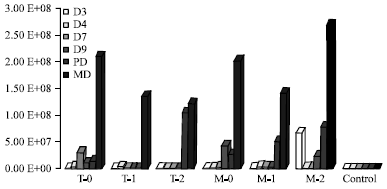 | |
| Fig. 2: | Multiplication of N. bombycis in 5th stage larva, pupa and moth of B. mori T = N. mylitta, M = N. bombycis, Treatment 0 = 1.52x108 spore mL-1 Treatment 1 = 1.52x107 spore mL-1 Treatment 2 = 1.52x106 spore mL-1. Control = No treatment, D = Day of 5th stage larva, PD = pupa and MD = Moth |
| Table 1: | Mortality % of Bombyx mori in different concentrations of N. bombycis and N. mylitta in different seasons |
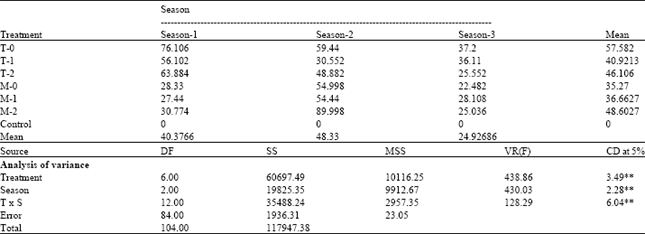 | |
| T = N. mylitta, M = N. bombycis, Treatment 0 = 1.52x108 spore mL-1, Treatment 1 = 1.52x107 spore mL-1, Treatment 2 = 1.52x106 spore mL-1, S1,S2 and S3 = Season-1 (Nov-Dec-Winter ), 2 ( Feb-Mar-Spring) and 3 (May-Summer), ** = significant at 1% level | |
Gradually the rate of multiplication of pathogen decreased (2.0x108 and 1.4x108 spore mL-1) with the higher inoculum concentrations of pathogen (M-0, 1.52x108 and M-1, 1.52x107 spore mL-1), respectively (Fig. 2).
The highest mean larval mortality (57.58%) was recorded in mulberry silkworm cross-infected by highest concentration (1.52x108 spore mL-1) of N. mylitta from tasar silkworm, (T-0). The highest mean seasonal mortality (48.33%) was recorded during March to April (Season-2). Lower mean mortality (35.27%) in larva of mulberry silkworm is recorded when it was infected by highest concentration (1.52x108 spore mL-1) of N. bombycis (M-0) and lower mean seasonal mortality(24.93%) was recorded during May (Season-3). The mean larval mortality % was always higher (66.48%) in mulberry silkworm during March-April (Season-2) than December-January (Season-1) (28.85%) and May-June (Season-3) (25.21%) due to infection of N. bombycis to mulberry silkworm (Table 1).
There is significant (p<0.01) difference among different seasons. Similarly, there is a significant interaction between seasons and treatments. This indicates the significant difference in impact of treatments in various seasons.
Transovarian transmission: N. bombycis and N. mylitta are transovarially transmitted. However, in both the cases highest inoculum concentration, M-0 (1.52x108 spore mL-1) and T-0 (1.52x108 spore mL-1) were very much effective for transovarial transmission. Where as, lower inoculum dosage T-1(1.52x107 spore mL) and T-2 (1.52x106 spore mL-1) as well as M-1 (1.52x107 spore mL-1) and M-2 (1.52x106 spore mL-1) slowly affected for transovarian transmission in later stages of development .In mulberry larva, N. bombycis multiplied maximum in 4th stage (9.3x108 spore mL-1) and while, N. mylitta was multiplied maximum in pupa stages (1.27x109 spore mL-1), during transovarial transmission (Fig. 3).
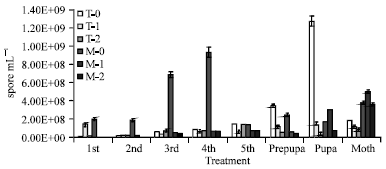 | |
| Fig. 3: | Multiplication of pathogens in different stage of B. mori during transovarial transmission T = N. mylitta, M = N. bombycis, Treatment 0 = 1.52x108 spore mL-1 Treatment 1 = 1.52x107 spore mL-1, Treatment 2 = 1.52x106 spore mL-1 Control = No treatment, 1st - 5th = Day of 1st-5th stage larva, Pre pupa, Pupa and Moth stage, Bar = Standard error of mean |
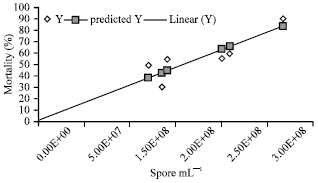 | |
| Fig. 4: | Regression analysis between the mortality % and multiplication of pathogens in B. mori |
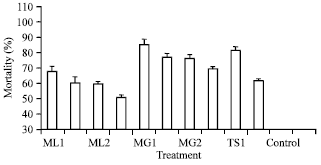 | |
| Fig. 5: | Mortality % in Antheraea mylitta in different concentrations of Nosema sp. generations (ML = N. bombycis, MG = N. assamensis, TS =N. mylitta, Treatment 1 = 1.52x108 spore mL-1, Treatment 2 = 1.52 x 107 spore mL-1, CON = No treatment, Bar = Standard error of mean) |
Interpretation of regression: Mortality % of larvae increased with the multiplication of pathogens. The positive correlation (R square = 0.890) effects between these two characters supports this findings. The regression equation between two parameters, Y = 1.311+3.078E-07x indicates that there positive increase of mortality % with the proportionate increase of multiplication of pathogens (Fig. 4).
Tasar silkworm: Rate of infection was low in 1st and 5th stage tasar larvae infected with N. bombycis, N. mylitta and N. assamensis. However, 2nd, 3rd and 4th stage infected larvae showed clear symptoms of microsporidian disease. The appearance of disease symptoms at 1st and early 2nd stage infected larvae was very slow though the feeding activity was normal. The black spots over the surface of the larval integument appeared from 3rd stage onwards. Most of the infected larvae were still in 3rd stage for a quite long period. Such larvae died in the 3rd stage itself or just after reaching 4th stage and were smaller in size than those, which did not carry black spots. Very few larvae could reach the 5th stage.
Range of larval mortality % was significantly (p>0.01) high (50.66-85.00%) in all infected batches. The larval mortality % was always higher in all batches of tasar silkworm infected and cross-infected by highest doses (1.52x108 spore mL-1.) of N. bombycis (ML-1), N. mylitta (TS-1) and N. assamensis (MG-1) in comparison to lower doses (1.52x106 or 1.52x107 spore mL-1) The highest larval mortality (85.00%) was recorded in tasar silkworm when cross-infected by highest dose (1.52x108 spore mL-1.) of Nosema assamensis from muga silkworm (MG-1) in comparison to same doses of N. bombycis and N. mylitta. But N. bombycis effected less mortality (67.77%) when cross-infected in the same host and same season. Average mortality was less (63.55%) during season-2 than in season-1. Maximum larval mortality was noticed in 3rd stage (39.44%) (Fig. 5).
On comparison, the different dose of pathogen among TS-1 (1.52x108 spore mL-1) MG-1 (1.52x108 spore mL-1) and ML-1 (1.52x108 spore mL-1), of which TS-1 was most virulent and MG-1 was more virulent in comparison to ML-1. Multiplication of spore of N. mylitta in tasar silkworm (TS-1) was always high in moth stages than in any larval stages and sharply increased from 5th larval stage (1.10x108 spore mL-1) to moth stage (4.0x109 spore mL-1). N. bombycis (ML-1) multiplied maximally in moth stage (1.39x108 spore mL-1) than 5th stage larva of tasar silkworm (6.88x105 spore mL-1).
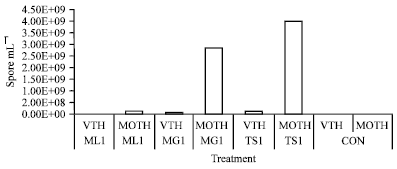 | |
| Fig. 6: | Multiplication of different concentrations of Nosema sp. in A.mylitta (ML = N. bombycis, MG = N. assamensis, TS =N. mylitta, Treatment 1 = 1.52x108 spore mL-1, Treatment 2 = 1.52x107 spore mL-1, CON = No treatment, VTH = 5th stage larva ) |
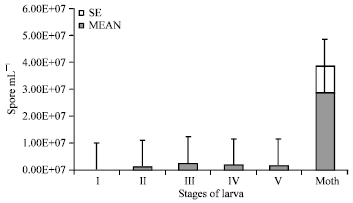 | |
| Fig. 7: | Multiplication of N. bombycis in tasar silkworm during transovarial transmission( I-V = 1st to 5th stage larvae, Bar = Standard error of mean ) |
The multiplication of N. assamensis (MG-1) in tasar silkworm was also increased in moth stage (2.84x109 spore mL-1) than 5th stage (5.1x107spore mL-1). Maximum multiplication of spore was observed in moth stage of tasar silkworm when N. assamensis (2.86x109 spore mL-1) and N. mylitta (4.02x109 spore mL-1) infected to tasar silkworm with higher inoculum concentration (1.52x108 spore mL-1) during 1st generation. The development and multiplication of microsporidia continued with the advancement of larval stage and it reached maximum in 3rd stage (1.60x106 spore mL-1) than any other larval stages in 1st generation when N. bombycis cross-infected to the tasar silkworm but maximum spores was harvested in moth stage in 1st generation (2.71x107 spore mL-1) and 2nd generation (2.63x107 spore mL-1) (Fig. 6).
Transovarial transmission: N. bombycis transovarially transmitted in tasar silkworm and higher inoculums concentration (1.52x108 spore mL-1) (M-0) was most effective for it. Maximum multiplication of spore (2.84x107 spore mL-1) was observed in the moth stage. The multiplication of spore was observed at a lower level (8.1x106-1.1x107 spore mL-1) during larval period. However, multiplication of spore in the moth stage in first generation (2.8x107 spore mL-1) was at par at the moth stage of next successive generation 2.86x107 spore mL-1 (Fig. 7).
Eri silkworm: Mortality in larval stages by Nosema sp. from mulberry, muga and tasar silkworm was not noticed even after 3rd successive inoculation with even high doses of pathogen. However, about 95% mortality was recorded when N. ricini was inoculated to eri silkworm in brushing stage, 2nd stage and 4th stage. Further, 95% of eri larvae were died by other diseases, like viral and bacterial after 3rd successive inoculation (1.52x108 spore mL-1) at 4th stage of silkworm infected by N. bombycis. 90 and only 5% worms died by other diseases after 3rd successive inoculation at 4th stage of silkworm infected by Nosema sp. from tasar and muga silkworm, respectively. Higher spore dose (1.52x108 spore mL-1) of N. ricini was effective for higher mortality in eri silkworm during August-September (Fig. 8).
The effective dose, 1.52x108 spore mL-1, for N. ricini was responsible for mortality of larvae as well as moths. The minimum number of spores required to initiate a disease in silkworm has been estimated. It appears to vary with the age of the host. In the present findings a minimum dose 1.52x106 spore mL-1 of N. ricini was responsible for mortality to muga silkworms, when infected to ‘0’ h 1st stage silkworm.
Eri silkworm is maximum resistant than other silkworms and only after repeated inoculation of pathogens larvae become weak and easily infected by other disease. Eri silkworm can withstand the extreme climatic condition. N. ricini is highly species specific.
Muga silkworm: Mortality % was always higher (52-100%) in all batches of muga silkworm infected by Nosema bombycis, N. mylitta, N. ricini and N. assamensis (Fig. 9). All the above pathogens were highly virulent and multiply very rapidly in muga silk worm irrespective of inoculum concentrations and seasons. A little quantity of pathogen was capable to produce disease and cause mortality in muga silkworm .This indicates the significant difference in impact of treatments in various seasons.
Larval mortality started during 4-5 days of post inoculation. N. bombycis was highly cross-infective to muga silkworm and infection was very rapid in comparison to infected control, when N. assamensis infected to muga silkworm. Mortality % was recorded always higher (73.67-99.17%) in all cross-infected batches. However, muga silkworms that cross-infected with N. bombycis was recorded highest mortality % (98.00) than infected control (95.00%) during August-September (Season-1) when inoculum concentration was higher (1.52x108 spore mL-1). But, N. mylitta effected less mortality (67.00%) during September-October (Season-2) when inoculum concentration was lower (1.52x107 spore mL-1). Mortality % was recorded lowest (52.00%) during Season-2, but it was very high in Season-1 (95%) in infected control batches. Average mortality % of all cross-infected and infected control batches were recorded less (71.96%) during Season-2 than Season-1 (84.00%) (Fig. 9).
The development and multiplication of microsporidia was continued with the advancement of larval stages and it was increased maximum (1.70x106 spore mL-1) in infected control batches during September-October (Season-2) when inoculated with higher inoculum concentration (1.52x108 spore mL-1) and minimum (1.85x105 spore mL-1) when cross-infected with N. mylitta during August-September (Season-1) with lower inoculum concentration (1.52x107 spore mL-1). Surprisingly, it was observed that the average multiplication of microsporidian were less (3.11x105 spore mL-1) in season-1 than in season-2 (5.60x105 spore mL-1) in all infected and cross-infected batches, but, average mortality % was higher (83.79%) in season-1 than season-2 (71.96%). Moreover, mature spores were found in the gut content of larva till survived. Generally, all above pathogen multiply rapidly during October-November (Season-2) in comparison to August-September (Season-1). Maximum multiplication of spores (1.70x106 spore mL-1) was observed from muga silkworm, when highest concentration (1.52x108 spore mL-1) N. assamensis (MG-1) infected to muga silk worm in season-2 (Fig. 10).
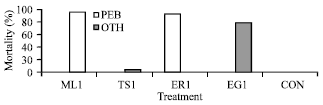 | |
| Fig. 8: | Mortality % in Philosamia ricini due to infection of N. ricini (ER-1) and other pathogens (ML = N. bombycis, MG = N. assamensis, TS = N. mylitta, Treatment 1 = 1.52x108 spore mL-1 CON = No treatment, PEB = Pebrine infected and OTH = Other pathogens) |
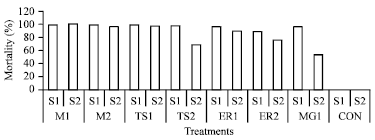 | |
| Fig. 9: | Mortality % in muga silkworm in different concentrations of Nosema sp. (ML = N. bombycis, MG = N. assamensis, TS = N. mylitta, ER = N. ricini, Treatment 1 = 1.52x108 spore mL-1, Treatment 2 = 1.52x107 spore mL-1, CON = No treatment, S = Season) |
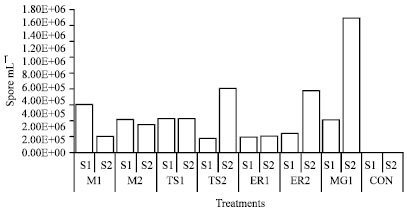 | |
| Fig. 10: | Multiplication of pathogens in muga silkworm in different seasons (ML = N. bombycis, MG = N. assamensis, TS = N. mylitta, ER = N. ricini, Treatment 1 = 1.52x108 spore mL-1, Treatment 2 = 1.52x107 spore mL-1, CON = No treatment, S = Season) |
DISCUSSION
This study reports first time the relationship of larval mortality with multiplication of Nosema sp. in relation with seasons, infection as well as cross-infection of mulberry and vanya silk in eastern and north eastern region.
In nature the pebrine disease occurred in mulberry, tasar, eri and muga silkworm (Rao et al., 1976). N. bombycis multiplied rapidly during season-2 as temperature and relative humidity were suitable for multiplication. Season-1 was the second favourable season for N. bombycis infecting B. mori. N. bombycis could not multiply rapidly during season-1 and 3 as temperature were lower and higher, respectively than the suitable for pathogen multiplication. It indicates that pathogen multiply in its suitable environment not favourable season of the host.
The temperature and humidity were moderate during season-2 regime coincides with the highest mortality, which might have acted as stimulatory factors of parasite (Dash and Nayak, 1998). Dasgupta (1950) made similar observation in B. mori. But the findings of Deviah and Krishnaswamy (1975) in B. mori stated minimum incidence of pebrine at high temperature and humidity which is deviated statement from the present findings. However, Dash and Nayak, 1998) observed lowest mortality in winter. Nomani et al. (1971) made contradictory statement and stated that temperature and humidity has no role with incidence of pebrine. However, such experiment of cross-infection of N. mylitta. to B. mori is lacking.
However, the mortality % was in decreasing trend in season-1 to season-2 to season-3 when N. mylitta cross-infected to B. mori. Mortality % was recorded significantly higher in B. mori larvae cross-infected with high inoculum concentration of N. mylitta during season-1 in 1st generation when temperature was favourable but relative humidity was low for development of pebrine spore. The parasitic effect on larval mortality is not directly related with the season but it is one of the factors in multiplication of pathogen and is related with the nature of parasite i.e., virulence, wild/domestic. The present findings corroborate the findings of Johnson (1989) and the infection rate that resulted from early application of Nosema locustae to grasshopper was initially higher than that caused by late application (Johnson, 1989). The pathogen develops more quickly at lower temperature and very slowly at higher temperature relative to development of the host. Slower rates and wider ranges of developmental times seen in infected insects may have implications for success in mating and in timely diapauses (Sloter et al., 1989). This also supports the present findings, where it was observed that N. mylitta multiplied maximally in lower temperature and more slowly at higher temperature.
In the present observation, multiplication of spore was highly significant (p<0.01) in all cross-infected larvae in second generation. However, N. mylitta with high inoculum concentration was very effective for high larval mortality in all the three seasons in second generation. Multiplications of spore were observed highest in moth and pupal stage of first and second generation respectively, when N. mylitta infected with high inoculum load. In general the mortality of insect is directly related to the size of the inoculums concentration of parasite received (Ghosh and Saha, 1995). In the present findings, it was observed that at least minimum inoculum load of pebrine spore was required for significantly high larval mortality in first generation but below that there was no effect on mortality. Weiser (1976) stated that time taken for establishment of the parasite for completion of its life cycle and production of spores depending on inoculum load and other environmental factors. The longevity of the instars and the amount of feeding is influenced the disease manifestation (Ishiwara and Fujiwara, 1965). Henry et al. (1973) obtained similar spore numbers of Nosema acridiophagus and N. cuneatum in Melanoplus sanguinipes at 20 days using 104 and 106 spore concentrations where spore production reaches the stationary phase but with a low concentration of 102 the spore production was lower when spore multiplication was still in progress at the time of harvest. If development is allowed to proceed to the stationary phase, multiplication of the parasite is greater with low concentrations than with high concentrations (Lai and Canning, 1983). But there is an apparent increase in multiplication with increased concentrations during an exponential phase and the difference between the multiplications of spore with the high and low initial spore concentration is narrow. Multiplications of pathogens depend upon the age of silkworm, time and other indirect factors (Sloter et al., 1989; Geetha Bai and Mahadevappa, 1995).
When a critical dose (1.52x107 spore mL-1) was inoculated then around 40% larval mortality was observed, in both the cases, when N. bombycis and N. mylitta cross-infected to B. mori. But 10 times above and lower that critical dose cause to higher (76%) and lower (25%) mortality, respectively, when N. mylitta infect to B. mori. Milner (1972) studied the effect of varying quantities of N. whitei spores on the mortality of larva of Tribolium castaneum, with a dose range from 1.6x103 to 1.0 x109 spores g-1 in flour medium. Milner reported a critical dose of 106 spores g-1 in medium above which the insects generally died as larvae, where as below that dose they generally emerged as adults. A particular concentration is effective for multiplication of spore and mortality (Windels et al., 1976) and below that level the concentration do not cause any larval mortality (Malone and Wigley, 1981).Therefore, the number of spores that a host can harbour and still function normally is important in determining the role of a microsporidia as parasite in nature (Windels et al., 1976).
It is clear from study that nosemosis develops more successfully in younger larvae and temperature can be considered as act ‘activator’ of the organism and no solely as a ‘depressor’ of the host, which is similar to the present findings, where the temperature 25-28°C is optimum for pathogen multiplications and early larval stage, 3rd and 4th stage are suitable for pathogen multiplication. The 3rd and 4th stages of tasar silkworm are more susceptible for microsporidian infection than 5th stage larva (Wilson, 1984; Patil, 1993). Decrease in mortality in the 5th instar pebrine infected larvae might be due to weak and inactive phase of parasite in 5th instar larvae and gradual development of immunity in the surviving larvae (Dash and Nayak, 1998).
In the present study N. mylitta transovarially transmitted with 100% efficiency during second generation. More than 98% mortality were recorded when larvae transovarially infected with higher inoculum load. It is assumed that higher rate of infection of the adult would increase the rate of transmission of the pathogen to the next generation thus ensure survival in nature Henry et al. (1973). Han and Watanabe (1988) found that N. bombycis was transovarially transmitted 100%, whereas, Nosema sp. M11 was transmitted only 1.2±0.41% and fifth stage larva was suitable for transovarial transmission. From the present results, it is obvious that N. mylitta is virulent to mulberry silkworm, B. mori (Chakrabarti and Manna, 2009).
The larval mortality of A. mylitta was higher in season-1 than season-2 in different inoculums concentration. Inoculums concentration of microsporidian is a major factor responsible for difference of mortality %. Temperature range of 26-28°C was optimum for multiplication of N. mylitta. The prevailing temperature was favourable in season-1 for pebrine diseases. Decrease in mortality in the 5th instar pebrine inoculated larvae might be due to gradual development of immunity in the surviving larvae (Nittono, 1960; Dash and Nayak, 1998). The longevity of the instars and the amount of feeding are some factors, which influence the disease manifestation (Ishiwara and Fujiwara, 1965). A lower inoculums concentration is responsible for lower mortality and higher inoculums concentration is responsible for higher mortality, when N. mylitta and N. assamensis infected to A. mylitta in the present findings. The time taken for establishment of the parasite for completion of its life cycle and production of spores varies considerably, depending on the age of host, species of the parasite, size of the inoculums concentration and other environmental factors (Weiser, 1976).
Maximum multiplications of spores were observed in moth stage than any larval stages of tasar silkworm in the present findings. This is also in agreement with the statement of Sloter et al. (1989) who stated that multiplication of pathogen depend upon the age of host when pathogen was inoculated. Season-1 was suitable for pathogen multiplication, where all the meteorological factors viz., temperature, relative humidity etc., stimulated for pathogen multiplication (Dash and Nayak, 1998). In the present finding, 3rd and 4th stages larvae of A. mylitta were suitable for multiplication of N. bombycis, N. mylitta and N. assamensis. This is in agreement with the observation of Hurpin (1959). But in case of N. bombycis infection, the multiplication of pathogen was recorded highest in 2nd instar infected larvae followed by 1st, 4th and 5th instar larvae (Geetha Bai and Mahadevappa, 1995). This is contradictory to the present findings. A particular concentration is effective for multiplication of spore and mortality Windels et al. (1976) and below that levels it do not cause any mortality (Malone and Wigley, 1981). Maximum spore was harvested, when larva were undergoing changes from pupa (6.88x105- 1.10x107 spore mL-1) to moth stage (1.86x107-4.02x109 spore mL-1) on 33-35 days in all infected batches in the present findings, which support the views of Lai and Canning (1983). The physiological changes in the insect could possibly have caused the development cycle of the parasite to switch on from a predominantly vegetative phage, merogony/schizogony to sporogony, resulting in an increasing spore production (Lai and Canning, 1983).
In the present findings, it was observed that the spore harvest was almost at the same level at moth stage in both the 1st and 2nd generations, when the spore production reached the stationary phase. This observation supports the view of Henry et al. (1973), Lai and Canning (1983) and Sloter et al. (1989). It was observed that N. bombycis was more efficient during transovarial transmission (100%) than N. mylitta (98-100%) and N. assamensis (98-100%). It supports the view of Han and Watanabe (1988). The female A. mylitta D. depending upon the intensity of infection can transmit the Nosema sp. with nearly 85-98% efficiency (Griyaghey and Sengupta, 1989).
With the advancement of larval age, the development and multiplication of pathogen continued. Further delaying the growth and development of larva was observed (Kobayashi and Kawase, 1981). Weight of 5th stage tasar larva increases slowly day by day due to gaining immunity slowly against the infection of pathogen (Nittono, 1960).
From the forgoing discussion, it is concluded that Nosema bombycis, N. assamensis and N. mylitta are virulent to A. mylitta and huge mortality in larval stage was occurred resulting in poor production of tasar silk.
Eri silkworm, Philosamia ricini is highly resistant to silkworm diseases. Nosema ricini n. sp. is able to produce pebrine disease in P. ricini, as this pathogen is highly species specific (Thangavelu, 1989). The number of spores (intensity of infection) that a host can harbour and still function normally is important in determining the role of a microsporidia as parasite in nature. The effective dose, 1.52x108 spore mL-1 for N. ricini is responsible for mortality of larvae as well as moths. This supports the view of Windels et al. (1976).
Both temperature at which the host was maintained and the size of the original inoculum were responsible for high mortality. Higher temperature (32°C) and higher spore dose (1.52x108 spore mL-1) of N. ricini were affective for higher mortality in eri silkworm during August-September. The minimum number of spores required to initiate a disease in silkworm has been estimated. It appears to vary with the age of the host. In the present findings a minimum dose 1.52x106 spore mL-1 of N. ricini is responsible for mortality to muga silkworms, when infected to ‘0’ h 1st stage silkworm. This supports the view of Milner (1972). Eri silkworm is maximum resistant than other silkworm and larvae become weak only after repeated inoculation of pathogens and easily infected by other disease. Eri silkworm can withstand the extreme climatic condition.
In the present study, N. bombycis N., N .mylitta and N. ricini were cross infected to muga silkworm and showed highly positive response causing development of severe pebrine disease. The previous study in this aspect is not available, except Patil (1989) got higher incidence of pebrine disease, when spores isolated from eri silkworm cross-infected to muga silkworm and Mistry (1979) cross-infected pebrine spore from eri to muga silkworm in natural condition and got 22.6% mortality. He suggested that such cross infection may be possible as the rearing of muga and eri silkworms are conducted side by side in villages in Assam.
Muga silkworm, Antheraea assamensis is very much susceptible to diseases, therefore, pathogen multiplies maximally in A. assamensis as per their virulence (Thangavelu et al., 1986). Muga silkworm looses its hybrid vigour and resistance against diseases due to continuous inbreeding depression (Gogoi and Goswami, 1998).
Teetor and Kramer (1976) informed that there was no discernible relationship between spore dose and subsequent infection of host, when he studied the quantitative pathogenicity of the microsporidia, Octospora muscaedomesticae in the adult black blowfly Phormia regina. They stated that lower than 103 spores per fly was able to overcome the host defenses plus the invasion of the host tissue with an increasing number of reingested new spores. But the present study with very lower dose (1.52x106 spore mL-1) of pathogen was effective to cause mortality, particularly in muga silkworm cross-infected by Nosema sp. from four different species of silkworms.
The magnitude of spore production is strongly age specific and thus, time dependent. In addition, under certain environmental conditions (24°C) infected larvae developed more slowly than healthy ones. These two effects of the disease lead to less synchrony between healthy and diseased larvae and even greater asynchrony between the production of spores and the presence of healthy larvae. In addition to these direct effects of the pathogen and host-parasite indirect effects may be occurred (Sloter et al., 1989). Due to presence of adjustment; the pathogen multiplies very rapidly become enormous in number within a short time in its primary host. However, pathogen multiplies slowly in its secondary host or accidental host to cope up with the new environment. The pathogen multiplies maximally to ensure certain percentage of pathogen gets entry in the next generation through transovarial transmission. It is reasonable to assume that higher rate of infection of the adult would increase the rate of transmission of the pathogen to the next generation thus ensure survival in nature (Henry et al., 1973).
It may be concluded that the larval mortality is directly proportional to the multiplication of Nosema sp. But the rate of multiplication of pathogen is different in mulberry and vanya silk depending upon the strain and critical concentration of pathogen, age of silkworm and seasons. Out of above factors critical concentration of N. bombycis is most important to initiate the disease development. Further time taking by the pathogen for disease development is depending upon the season of silkworm rearing. Less than 100 mature spores of N. bombycis in 5th stage of larvae and less than that quantity in earlier stage are able to cause pebrine disease in B. mori within 4-7 days during favourable environment of pathogen (Season-2). Whereas, at least 1.52x106 spore mL-1 of N. mylitta is required in 5th stage larvae reared under moderate temperature and relative humidity (season-2) are able to produce pebrine disease in B. mori during cross-infection. Either quantity of spore is required more or duration of life cycle of the pathogen will be increased when pathogen is passed through unfavourable environment. N. mylitta is virulent compare to N. assamensis and multiplication of N. mylitta is high than N. assamensis in A. mylitta but mortality was found less when tasar larvae infected with N. mylitta than N. assamensis irrespective of critical concentration due to host parasite adjustment since decades. But N. bombycis with higher critical dose can only effective for high mortality (98%) in muga silkworm than infected control. All the above pathogens with higher critical dose are able to produce pebrine disease to their secondary host (cross-infectivity) when coincides with the favourable environment of the pathogen only.
ACKNOWLEDGMENTS
The authors are grateful to late Dr. A.K. Bajpai, Director, Central Sericultural Research and Training Institute, Central Silk Board, Berhampore, West Bengal, India for their keen interest and kind help in this work. The authors are grateful to the Head of the Department of Zoology, The University of Calcutta, Kolkata for providing laboratory facilities to complete the piece of research work.
REFERENCES
- Han, M.S. and H. Watanabe, 1988. Transovarian transmission of two microsporidia in the silkworm, Bombyx mori and disease occurrence in the progeny population. J. Invertebr. Pathol., 51: 41-45.
CrossRef - Johnson, D.L., 1989. The effects of timing and frequency of application of Nosema locustae (Microspora: Microsporida) on the infection rate and activity of grasshoppers (Orthoptera: Acrididae). J. Invertebr. Pathol., 54: 353-362.
CrossRef - Thangavelu, K., A.K. Bhagowati and A.K. Chakraborty, 1986. Inbreeding depression of heterosis in muga silkworm, A. assama. Ins. Sci. Appl., 7: 723-725.
Direct Link