
D.D. Tibuhwa
Department of Molecular Biology and Biotechnology, University of Dar es Salaam, P.O. Box 35179, Dar es Salaam, Tanzania
M.N. Muchane
Department of Botany and Zoology, National Museums of Kenya, P.O. Box 40658, Nairobi, Kenya
C.W. Masiga
ASARECA P.O. Box 765, Entebbe, Uganda
C. Mugoya
ASARECA P.O. Box 765, Entebbe, Uganda
M. Muchai
Department of Botany and Zoology, National Museums of Kenya, P.O. Box 40658, Nairobi, Kenya
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 6 | Page No.: 399-410







ABSTRACT
The Serengeti-Mara ecosystem (SME) extends on both sides of Tanzania and Kenya. It comprises a unique and highly conserved ecosystem in the world. In spite of the importance of macro-fungi in maintaining and promoting productivity by enhancing nutrient cycling, researchers in the SME has been neglecting them. This work inventoried macrofungi in the drier and wetter side of SME based on 3 land use types: Crop land, woodland and grassland in protected and unprotected area, during both dry and wet season. The method based on fruit body recording which included fungi taxonomic field work and documents the macrofungi species presence. It also included analysis using diversity species indices for comparisons in terms of species diversity and richness across the habitats. A total of 92 species of macro-fungi distributed in 17 families and 33 genera were encountered. Most of these taxa (55.4%) were found in Tanzanian side while 44.5% were found in Kenyan side. Macro-fungi species of the family Lyophyllaceae (23%), Agaricaceae (21%) and Polyporaceae (12%) were the most commonly represented taxa in the ecosystems. Woodland habitat recorded the highest number of macro-fungi species (47%), followed by grassland (37%) while only few species were encountered in the agricultural farms (16%). The wet region recorded significantly high macro-fungi species compared to dry region. The Reyni diversity ordering showed tremendous decreases in species diversity in plots outside the park compared to those found inside the park. This result implies that disturbance affects myco-biota diversity which calls for the need of conservation and modification of agro-ecosystems. Introducing agro-forestry ectomycorrhiza tree species can transform the agro-ecosystems to mimic natural ecosystems and be an alternative source of mushroom resources.
PDF Abstract XML References Citation
Received: September 18, 2011;
Accepted: November 26, 2011;
Published: December 10, 2011
How to cite this article
D.D. Tibuhwa, M.N. Muchane, C.W. Masiga, C. Mugoya and M. Muchai, 2011. An Inventory of Macro-fungi and their Diversity in the Serengeti-Masai Mara Ecosystem, Tanzania and Kenya. Journal of Biological Sciences, 11: 399-410.
DOI: 10.3923/jbs.2011.399.410
URL: https://scialert.net/abstract/?doi=jbs.2011.399.410
DOI: 10.3923/jbs.2011.399.410
URL: https://scialert.net/abstract/?doi=jbs.2011.399.410
INTRODUCTION
Serengeti Mara ecosystem (SME) is the largest wildlife sanctuaries in the world covering about 25,000 km squares (9,700 sq mi) across the two countries Tanzania and Kenya (Thirgood et al., 2004). The SME is home for a wide range of biodiversity ranging from large mammals such as elephant to small microorganism. The interaction of these organisms makes a complex ecosystem which supports life for about one million pastoralists and agriculturalists (Sinclair and Norton-Griffiths, 1995). Plant and animal biodiversity in the rangelands surrounding the protected core of the SME has been negatively affected due to considerable changes in land cover and land use systems for over past 40 years now (Serneels et al., 2001). In the SME drudges for restoration and information on conservation of the ecosystems has concentrated on large organisms such as wild animals (Borner et al., 1996), birds (Fishpool and Evans, 2001), Amphibians and reptiles (Kreulen, 1975) and vegetations (Schmidt, 1975; Belsky, 1987). However, little attention has been given to macro-fungi, although they contribute greatly on balancing the ecosystem in terms of ecological niche, nutrient recycling as well as symbiotic associations. Recently, there have been global efforts to promote innovation/farming practices which sustain biological diversity without compromising food and income needs of the local community outside the protected areas (Buck et al., 2001). To accomplish this mission it is very crucial to have a wide knowledge of all biodiversity contributing to sustainable ecosystems, including macro-fungi.
Macro-fungi include fungi distinguished by forming fruit bodies commonly referred to as mushrooms visible to naked eye. Macro-fungal studies have long been of interest to scientists and community due to their significant roles. They form an integral part of the forest environment and perform significant roles in forest ecosystems such as forest flourishing through mycorrhizal associations and nutrient recycling through saprobic life (Boa, 2004). Macrofungi also are very useful in food industries (Lindequist et al., 2005) as well as enormous use of their fruit bodies for the general welfare in human life. Edible mushrooms provide a wide range of minerals and vitamins although the total nutrient contents vary significantly among species. It is amicable that a diverse fungal population contributes to a diverse diet for wildlife and humans. Mushroom cultivation provide an opportunity to improve local farmers’ livelihoods and reduce dependence on natural resources. The sustainability of macro-fungi is thus important to maintain and promote productivity of croplands, rangelands and forests and may be critical for maintenance of biodiversity (Allen et al., 1995) and livelihoods.
Despite the importance of macro-fungi in both natural and agro-ecosystems, little is known about their community structure and dynamics especially in the tropics (Hawksworth, 1991). Macro-fungi in the tropics are sparingly studied, relatively under-utilized and very little information on their diversity is known (Osemwegie and Okhuoya, 2009; Mueller et al., 2004; Osemwegie et al., 2006; Labarere and Menini, 2000). On the other hand, fungal diversity is usually overlooked during consideration of management of forest ecosystems (Amarathus, 1998), yet successful conservation efforts in any ecosystems may require understanding mushroom communities in terms of ecology and distribution.
This study aimed at inventorying the macro-fungi community across different land uses with different level of human disturbances in the SME. The study was conducted as a baseline for identifying land use practices with potential of conserving macro-fungi communities and identifying sustainable utilization of some fungal species with economic importance to the local community in this area and as one among the ways of conserving the remaining diversity while improving rural livelihood.
MATERIALS AND METHODS
Study site: Serengeti-Mara ecosystem is made up of protected land (Serengeti National Park, Ngorongoro and Conservation Area) in Tanzania and Maasai Mara National Reserve in Kenya (Fig. 1). The protected area in the east is surrounded by semi-arid rangelands characterized by less rainfall (c.800 mm per year) inhabited by Maasai agro-pastoralists while the west side is wet with high rainfall (c.1200 mm per year) comprising highly intensive agriculture. The land use system in this region comprises of natural woodland and grassland inside and outside the park. Natural woodland and grassland inside the park are characterized by minimal human disturbances while woodland and grassland outside the park are characterized by heavy human disturbances. Agricultural systems outside the protected core of SME comprise of small-scale subsistence farming of cassava, millet, maize and beans. Intensive agriculture with selected commercial maize and wheat varieties grown with heavy external inorganic fertilizer inputs is also practiced especially in Kenyan side.
Experimental design: A one year survey was conducted during the short rain of September-November 2009 and long rains of March-June 2010 in Kenyan and Tanzanian side of SME. The study was conducted in pre-established study land use/habitat categories containing uniform land use differing according to major landuse types viz., (i) indigenous woodland (ii) natural grasslands and (iii) Cropland with mixed subsistence cropping in both Kenya and Tanzania as well as maize and wheat monocropping in Kenyan side. For each land use, plots of 10x5 km were selected inside the park (protected) and outside the park (unprotected) and one crop land from both dry and wet region of the SME (Fig. 1). In Tanzania, the plots included (i) Kibeyo woodland outside the park (TSWWO), (ii) Tabora B woodland inside the park (TSWWI), (iii) Gibaso grassland outside the park (TSCPWO), (iv) Lamai grassland inside the park (TSGWI) and (v) Nyansurura crop land (TSCPWO) in the wet region of SME. In the dry side of SME the study plots included (i) Ololosokwan woodlands outside the park (TSWDO), (ii) Lobo woodland inside the park (TSWDI), (iii) Togoro Plains grassland outside the park (TSGDI), (iv)Wasso grassland inside the park (TSGDO) and (v) Mdito crop land (TSCPDO). In Kenya, the study plot comprised of (i) Isokon woodland outside the park (KMWWO1), (ii) Isokon grassland outside park (KMGWO1), (iii) Serena woodland inside the park (KMWWI1), (iv) Ololo gate grassland inside the park (KMGWI1), (v) Maize mono-cropping (KMCPWO1-1), (vi) large-scale maize-beans mixed cropping (KMCPWO1-2) and (vii) small-scale maize-beans mixed cropping (KMCPWO1-3) in the wet region of Mara. In the dry region the study plots included (i) Siana woodland outside the park (KMWDO1), (ii) Olongayanat grassland outside the park (KMGDO1), (iii) Nkama woodland outside the park (KMWDI1), (iv) Posse plains grassland inside park (KMGDI1), (v) Maize mono-cropping (KMCPDO1-1), (vi) wheat mono-cropping (KMCPDO1-2) and (vii) maize-beans mixed cropping (KMCPDO1-3).
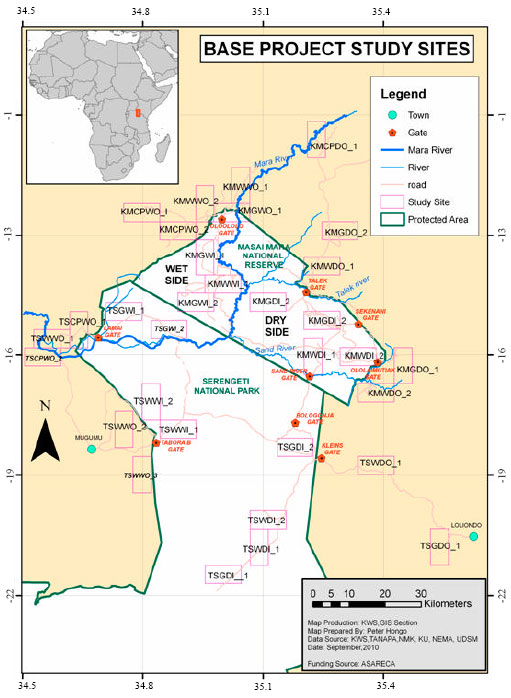 | |
| Fig. 1: | Map of Serengeti-Masai Mara showing the study sites |
In each sampling plot (10x5 km) in each land use types, four transects each measuring 1x 0.05 km were laid out, 1 km away from each other and 0.5 km away from the road. Macro-fungi fruiting bodies (mushrooms) were surveyed twice during dry season September-November, 2009 and wet season April-June 2010 in each transects.
Macro-fungi sampling: Mushrooms were collected randomly along 1x0.05 km transects. Observed mushrooms were photographed in situ (Fig. 2) and features such as smell, habitat, colour, nature of substrate and associations recorded. Few representative fruit bodies of each collection were dehydrated using silica gel in the air tight plastic bags for transportation to the laboratory for herbarium collection and microscopic analysis whenever necessary.
 | |
| Fig. 2(a-i): | Some macrofungi fruit bodies collected in the SME (a) Termitomyces eurhizus (b) Funalia polyzona (c) Coprinus comatus (d) Mycena sp. (e) Macrolepiota sp. (f) Auricularia polytrica (g) Termitomyces microcarpus (h) Pycnoporus sanguineus (i) Schizophyllum commune |
In the laboratory, the collected macro-fungi identification was based on macro-micromophological features by comparison using coloured field guide books, monographs of colored mushrooms such as (Largent and Their, 1984; Largent, 1986; Kirk et al., 2001; Harkonen et al., 1995, 2003; Mueller et al., 2004; Lincoff, 2005) and internet facility. Some of the mushrooms after freshly observation were dehydrated using silica gel and deposited at the mycological herbarium of the University of Dar es Salaam (UDSM) and National Museums of Kenya.
Data analysis: Every species of macro-fungi occurring at each site was recorded and the percentage of species occurring at one land use type was calculated as:
Species richness and diversity index was statistically done using the Shannon-Wiener index (Nolan and Callahan, 2006) because it favors neither rare nor common species disproportionately while counting all species according to their frequency thus considering weighting to each species exactly by its frequency in the sample. The Sorensen similarity Index and Reyni diversity of species ordering were both done according to Sorenson (1948) using PISCES Community analysis Package Version 1.50 all under license of PISCES Conservation Ltd (PISCES Conservation, 2001), (IRC House, Pennington, Lymington SO41 8GN UK).
RESULTS
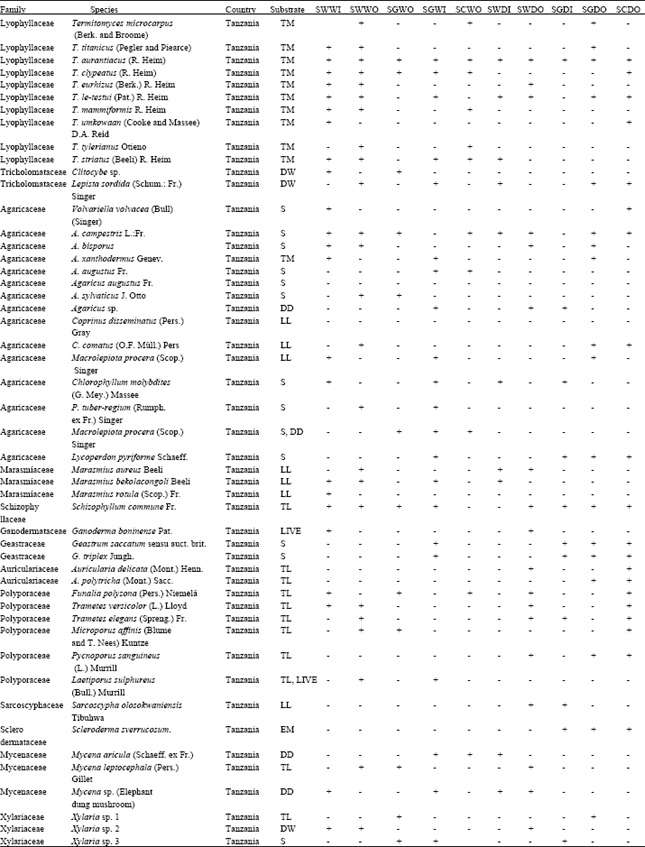
Macro-fungi representation in Tanzania and Kenya: A total number of 92 macro-fungi species were encountered throughout the SME (Table 1a and b). Out of the 92 species, 70.3% were identified to species level, 18% to genus level and 11.7% completely unidentified. The encountered taxa belonged to two sub-divisions; the Basidiomycota and Ascomycota were distributed in seventeen families and 33 genera. The macro-fungi families encountered in the two countries differed. Species belonging to families Schizophyllaceae, Ganodermataceae, Geastraceae, Auriculariaceae, Sarcoscyphaceae, Xylariaceae and Sclerodermataceae were encountered only in the Serengeti in Tanzania.
| Table 1a: | Macro-fungi species observed across the SME from Tanzania |
 | |
| Table 1b: | Macro-fungi species observed across the SME from Kenya |
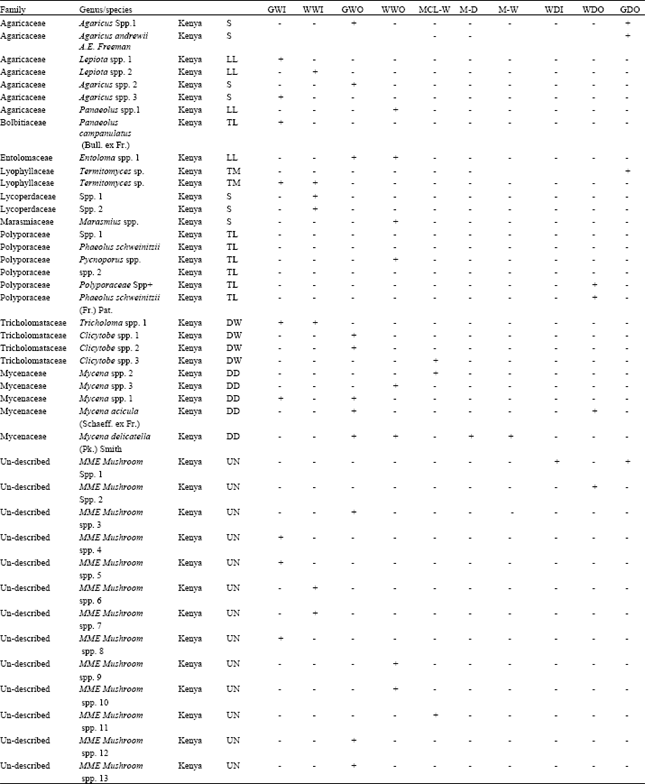 | |
| +’: Present; ‘-’: Absent; TM: Termite Mound; TL: Tree log; DW: Decaying wood; LL: Leaf Litter; WD: Wood Debris; S: Soil; EM: Ectomycorrhiza; LIVE: Live Tree; DD: Decomposing dung. Kibeyo (TSWWO); Tabora B (TSWWI); Nyansurura (TSGWO); Lamai (TSGWI), Togoro Plains (TSGDI), Wasso (TSGDO), Lobo (TSWDI), Ololosokwan (TSWDO), Mdito (TSCPDO), Gibaso (TSCPWO), Isokon woodland outside the park (WWO), Isokon grassland outside park (GWO), Serena woodland inside the park (WWI), Ololo gate grassland inside the park (GWI), Maize mono-cropping (M-W), large-scale maize-beans intercropping (MCL-W), Siana woodland outside the park (WDO), Olongayanat grassland outside the park (GDO), Nkama woodland outside the park (WDI), Posse plains grassland inside park (GDI), Maize mono-cropping (M-D), wheat mono-cropping (W-D), maize-beans mixed cropping (MCL-D) | |
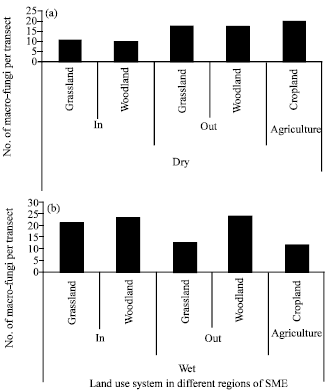 | |
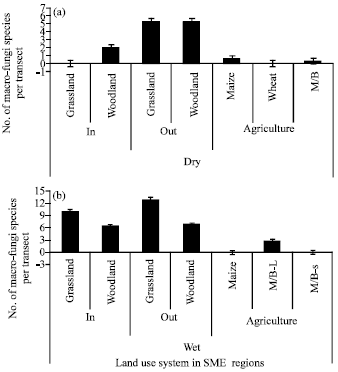
| Fig. 3(a-b): | Number of macro-fungi species per transect in different land uses systems: woodland and grassland inside and outside the park and agricultural farms in dry (a) and wet (b) side of Serengeti Mara Ecosystems (SME), Tanzania side |
Like wise species belonging to family Bolbitiaceae, Entolomataceae and Lycoperdaceae were only encountered in Maasai Mara in Kenyan (Table 1a). However species belonging to Agaricaceae, Lyophyllaceae, Marasmiaceae, Polyporaceae, Mycenaceae and Tricholomataceae families were found in both Tanzania and Kenya (Table 1b). In total thirteen families were encountered in Tanzania while nine families were encountered in Kenya.
The total number of macro-fungi species were statistically different in the two countries (p = 0.02), with high number of species encountered in Tanzania. Total of fifty macro-fungi species comprising of 54% of the total species were collected from Tanzanian side compared to 42 species comprising of 46% of the total collected from Kenya (Fig. 3, 4). However, the total number of species represented in Agaricaceae, Lyophyllaceae, Mycenaceae and Tricholomataceae (p<0.05 in all cases) differed significantly in the two countries. Species belonging to Agaricaceae and Lyophyllaceae were high in Serengeti in Tanzania than in Kenya while species belonging to Mycenaceae and Tricholomataceae were high in Maasai Mara in Kenya than in Tanzania.
 | |
| Fig. 4(a-b): | Number of macro-fungi species per transect in different land systems: woodland and grassland inside and outside the park and agricultural farms in dry (a) and wet (b) side of Serengeti Mara Ecosystems (SME), Kenya side |
The species belonging to the family Agaricaceae and Lyophyllaceae were the best represented taxa (30 and 20% representation, respectively) in Tanzania while the other families recorded less than 10% of species representation with exception of Polyporaceae family which had 12% representation. Alternatively, species belonging to the family Agaricaceae (17% representation), Polyporaceae (14% representation) and Mycenaceae (12% representation) were the best represented taxa in Kenya while the other families recorded less than 10% of species representation with exception of Tricholomataceae family which had 10% representation (Fig. 5).
Most of the macro-fungi recorded in SME were saprophytic, mostly colonizing the wood and litter-based substrates. Wood and litter supported the highest macro-fungi species (24%), followed by Termites cultivated mushrooms (23%) while decomposing dung (11%) and ecto-mycorrhiza fungi (fungi that grow with association plant roots) supported the least macro-fungi taxa (only 1%; Fig. 6a). The distribution of mushroom species in the three land use habitats showed the woodland leading with a total of 47.0% followed by grassland with 37% and the crop land recorded the least with 16.0% (Fig. 6b).
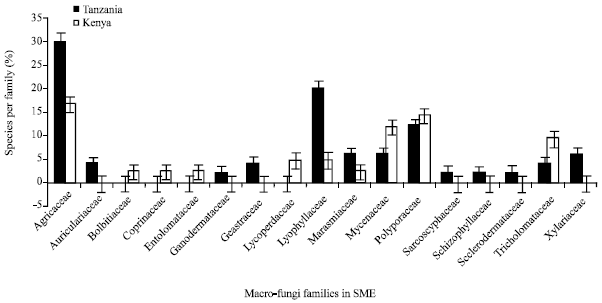 | |
| Fig. 5: | Number of species in different families encountered in the study area from both sides Tanzania and Kenya |
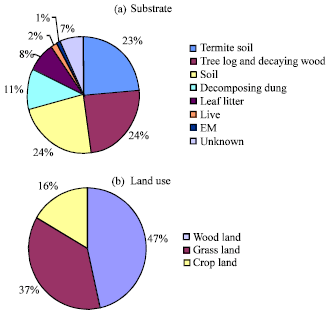 | |
| Fig. 6(a-b): | Quantitative distribution of macrofungi in the SME based on (a) substrate (b) Land use types from both sides Tanzania and Kenya |
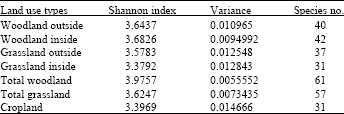
Effect of land use on macro-fungi species: The wet region of SME recorded significantly high macro-fungi species compared to dry region of SME (p = 0.007; average of 5.52±2.36 vs. 2.00±19 species per land use). In general woodland habitat represented highest species diversity with Shannon index (3.9757) followed by Grass land (3.9247) while crop land had the least diversity (3.3969). In Tanzania the dry region of SME, woodland and grassland outside the park recorded the highest species richness compared to woodland and grassland inside the park.
| Table 2: | Shannon diversity indices of different land use types in the SME |
 | |
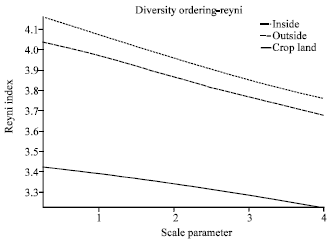 | |
| Fig. 7: | Reyni species diversity ordering across the three area: Protected area inside the park, unprotected area outside the park and the highly disturbed cropland |
On the other hand the crop land in the dry region recorded high species richness (20 species) comparable to those in woodland and grassland outside the park (Fig. 3a). In wet region of SME different land use systems affected the species richness but not the species diversity. The macro-fungi species richness was not different in woodland inside (23 species) and outside the park (24 species), however high number of species was observed in grassland inside the park (21 species) than in grassland outside the park (12 species; Fig. 3b). In the cropland, macro-fungi species richness (11 species) was low compared to woodland and grassland inside the park but was not different from grassland outside the park. Unlike species richness, the comparatively macro-fungi species diversity in the two countries was higher in protected core of SME (inside the park) than unprotected core of SME (outside the park; Table 2). This has been depicted by Reyni species diversity ordering which showed relatively higher and stable diversity inside the park but generally poor and progressive decreasing in the crop land (Fig. 7).
DISCUSSION
Mushrooms have short life cycles (1-20 days) and different species appear at different times a year and in some cases individual species may not appear every year. The results of this study is based on two seasons of mushrooms fruiting and since the observed species number increases nonlinearly with sampling effort (Gotelli and Colwell, 2001), mushroom in this study may be under estimate of the total mushroom species in this region. However, the result of this study is very vital since it provides baseline for further research in this area and increases our knowledge of macro-fungi diversity in SME. In addition this is the first report of macro-fungi in SME which provides an important plan to monitor the effectiveness and needs for macro-fungi conservation in SME and related savannah ecosystems.
Contrary to our expectations, many mushroom species were encountered during the dry season than in the wet season. This was as a result of changing climate which resulted in to unexpected rainy season in November, 2009 which extended through wet season (March-May 2010). Mushrooms growth depends so much on rainfall conditions and macro-fungi are known to appear when the conditions for growth are favorable (Packham et al., 2002). Mushroom that fruited between the sampling seasons or not at all during the study period could have been missed. Detailed monitoring studies are thus necessary to document all fungi species which could have been missed during the two surveys and at the same time document the sprouting period of each fungal species after the on-set of rainy season.
About 30% of collected macrofungi in this study were not identified, 11.7% completely unidentified while 18% identified to genus level. This might probably be due to deterioration of some mushrooms before proper identification was carried out on one hand but also on the other hand the fact SME ecosystem is very complicated with special unique habitats which might favor the diversified range of macro fungi and that to the best of our knowledge no any study has ever been done on the macrofungi from this unique habitats, suggesting that probably many taxa could be new to science. In fact one species of Ascomycetes (Sarcoscypha ololosokwaniensis sp. nov.) has been recently found in the Serengeti side of SME (Tibuhwa, 2011). This necessitates more detailed taxonomic studies using both morphological and molecular studies to confirm this.
High numbers of mushroom species were recorded in wet region of SME than the dry region. This result was expected due to variations in the climate (macro and micro) between the two regions, litter fall dynamics and composition, differences in diversity of tree species among others. The dry region of SME is characterized by less rainfall and high temperatures as well as low plant diversity hence low wood substrates. This study also suggests that, the nature of substrate chemistry and micro-environment may also differ in the two regions affecting the distribution of mushrooms species, however this merits further studies.
High numbers of mushroom taxa were found in Tanzania side of SME than in the Kenya side. Taking into considerations the climatic conditions in the two countries are similar, the differences could have been as a result of different managements systems between the two countries. Managements inside the protected and unprotected core of SME differs between the two countries and disturbances levels outside the park as well as intensive agriculture have been indicated as more pronounced in Kenyan side than in the Tanzania side (Serneels et al., 2001).
The Serengeti ecosystems recorded rich diversity of mushroom in the subdivision Basidiomycota with the genus Termitomyces in the family Lyophyllaceae leading in Serengeti Park while Agaricus and Mycena were leading in the Maasai Mara. This may be due to the fact that the members of Termitomyces live symbiotically with termites of the family Termitidae thence their distribution are synchronized (Tibuhwa et al., 2010). Apart from the host distribution, inside parks is a highly protected area experiencing very little human disturbance which might attribute to good condition that favor flourishing of Termitidae which cultivate them in their gardens. In fact it is a kind of unplanned enrichment for termite to cultivate Termitomyces in their gardens. Interestingly in protected woodland inside Serengeti National Park (Tabora B plot) Termitomyces mushrooms were fruiting out abundantly like flowers in the garden (Fig. 2). Noticeably low numbers of Termitomyces specie were recorded in the Masai Mara despite the high numbers of termites in this region. This suggests that the termite species in this region were non-fungi cultivators or the sampling season may have missed the proliferation period of the mushroom species. The high representation of the Agaricus and Polypore fungi in the SME could be attributed to the fact that most of these species are saprophyte, capable of biodegrading many recalcitrant organic-based substrates (Lynch and Thorn, 2006), present in some of the land use systems like woodland and grassland. In addition species from Agaricaceae are known to not to associate with a given habitat, they are able to grow anywhere provided the conditions are favourable (Uzun, 2010). In line with observation from other studies, the high abundance of mushrooms in wood habitats with lots of decaying woody debris may be related to their high moisture retention (Edmonds, 1991; Graham et al., 1994). Wood-based substrates also have been shown elsewhere to be a major determinant of mushroom diversity in woodland vegetations (forest and agro-forests) in both temperate and tropical regions (Osemwegie et al., 2010). Mycena species were largely found growing in wild animal dung in the grasslands and woodlands as well as farmyard manure in the farms, showing the contribution of animals to macro-fungi diversity in this region. Ectomycorrhiza fungi (ECM) species were recorded in small proportion in this study. Low species of ECM suggest the lack of ECM plant host species and need to introduce ECM host (plant) species in this area to diversify the mushroom diversity.
Natural ecosystems especially the forests ecosystems have been shown to have a wide range of mushroom assemblage, diversity, abundance and species richness due to abundance of wood substrates from different plant species. This was also noted in this study where by woodland and grassland were the best sources of mushroom compared to agro-ecosystems (Fig. 3, 4). On the other hand agro-ecosystems are usually poor in mushroom diversity due to low or lack of wood substrates. However, the mushroom diversity in these systems may be increased through use of organic inputs or through intercropping of agro-forestry tree species with the food crop. We have shown in this study that application of farmyard manure in some of the cropping systems supported the growth of some Mycena species. Protected areas did not support high species richness than unprotected areas. However, Reyni species diversity ordering (Fig. 7) showed that protected areas (inside park) have higher species diversity and are relatively stable compared to unprotected areas (outside the park) and noticeably were very low diversity in the crop land which was shown to be progressively decreasing. This result demonstrated that disturbances affect mycobiota diversity and distribution which consequently may lead to loss of biodiversity. Lack of many species in protected areas suggest less disturbances in park resulting to less wood substrate turnover compared to unprotected areas outside the park as well as presence of termites mounds.
CONCLUSION
Based on the data presented in this study, it appears that the SME with unique complex ecosystem comprises very high diversity of macro-fungi which are not yet studied and probably many new to science. It also shows that protected areas have high diversity of macrofungi than unprotected and the wood land is the best habitat that provides suitable substrate which supports much macrofungi growth. This calls for conservation of natural ecosystems and the need to modify agro-ecosystems to mimic natural ecosystems through introduction of agro-forestry tree species and the use of organic farming which may both increase the proliferation of some mushroom species in agricultural systems and be an alternative source of mushroom resources. Although, studies of utilization of mushroom by the local communities in this region are still under way we expect an increase in collection of wild mushroom following the increasing awareness of mushroom resources as food and medicine. This increase necessitates the need to cultivate some selected mushroom species as an alternative source of food, medicine and finance to the local community to reduce human pressure on the natural ecosystems.
ACKNOWLEDGMENTS
Author are grateful to the Association of Strengthening Agricultural Research in Eastern and Central Africa the (ASARECA) project that sponsored the field work, the armed guard from Tanzanian and Kenya side wildlife division who escorted us in the park during mushroom gathering.
REFERENCES
- Allen, E.B., M.F. Allen, D.J. Helm, J.M. Trappe, R. Molina and E. Rincon, 1995. Patterns and regulation of mycorrhizal plant and fungal diversity. Plant Soil, 170: 47-62.
Direct Link - Belsky, A.J., 1987. Revegetation of natural and human-caused disturbances in the Serengeti National Park, Tanzania. Vegetation, 70: 51-60.
Direct Link - Hawksworth, D.L., 1991. The fungal dimension of biodiversity: Magnitude, significance and conservation. Mycol. Res., 95: 641-655.
Direct Link - Gotelli, N.J. and R.K. Colwell, 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett., 4: 379-391.
CrossRef - Labarere, J. and G.U. Menini, 2000. Collection, characterization, conservation and utilization of mushrooms, germplasm resources in Africa. Proceedings of the 1st International Congress for the Characterization, Conservation, Evaluation and Utilization of Mushroom Genetic Resources for Food and Agriculture, March 9-13, 1998, FAO, Bordeaux, France, pp: 17-63.
- Lindequist, U., T.H.J. Niedermeyer and W.D. Julich, 2005. The pharmacological potential of mushroom. Evidence-based Complement. Evidence-Based Complement. Aletern. Med., 2: 285-299.
CrossRefDirect Link - Lynch, M.D.J. and R.G. Thorn, 2006. Diversity of basidiomycetes in Michigan agricultural soils. Applied Environ. Microbiol., 72: 7050-7056.
CrossRefDirect Link - Osemwegie, O.O., E.G. Eriyaremu and J. Abdulmalik, 2006. A survey of macrofungi in Edo/Delta region of Nigeria, their morphology and uses. Global J. Pure Applied Sci., 12: 149-157.
Direct Link - Osemwegie, O.O. and J.A. Okhuoya, 2009. Diversity of macrofungi in oilpalm agroforest of Edo State Nigeria. J. Biol. Sci., 9: 584-593.
CrossRef - Osemwegie, O.O., J.A. Okhuoya, A.O. Oghenekaro and G.A. Evueh, 2010. Macrofungi community in rubber plantations and a forest of Edo State, Nigeria. J. Applied Sci., 10: 391-398.
CrossRef - Packham, J.M., T.W. May, M.J. Brown, T.J. Wardlaw and A.K. Mills, 2002. Macrofungal diversity and community ecology in mature and regrowth wet eucalypt forest in Tasmania: A multivariate study. Aust. Ecol., 27: 149-161.
CrossRef - Serneels S., M.Y. Said and E.F. Lambin, 2001. Land cover changes around a major African wildlife reserve: The mara ecosystem (Kenya). Int. J. Remote Sensing, 22: 3397-3420.
Direct Link - Sorenson, T., 1948. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Biol. Skrifter, 5: 1-34.
Direct Link - Tibuhwa, D.D., 2011. Morphology and taxonomy of Sarcoscypha ololosokwaniensis sp. Nov: A new Ascomycota species from Serengeti National Park-Tanzania. J. Yeast Fungal Res., 2: 1-6.
Direct Link - Uzun, Y., 2010. Macrofungal diversity of Ardahan and Igdir province (Turkey). Int. J. Botany, 6: 11-20.
CrossRefDirect Link