
Masoud Soltanialvar
Division of Avian Diseases,Islamic Azad University, Shoushtar Branch, Khozestan, Iran
Ali Bagherpour
Division of Microbiology, Islamic Azad University, Shoushtar Branch, Khozestan, Iran
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 4 | Page No.: 340-344







ABSTRACT
H9N2 influenza viruses have been circulating in Iran since 1998 but a systematic investigation of H9N2 in Iran has not been undertaken since 2002. The present study aimed to analyze and evolution of H9N2 influenza viruses in Iran from 2008 to 2009. To determine the genetic relationship of Iranian viruses, the PB2 genes from five isolates of H9N2 viruses isolated from commercial chickens in Iran during 2008-2009 were amplified and sequenced. The Iranian isolates did not exhibit insertions or deletions within (PB2) gene as compare with their prototype A/turkey/winconsin/66 but rather numerous point mutations were registered. The PB2 gene of Iranian viruses showed great genetic diversity and shared a high level of similarity with PB2 genes from H7 subtype rather than with established H9N2 Eurasian lineages. The results of this study indicated that during recent years the Iranian H9N2 viruses have undergone extensive genetic reassortment which led to the generation of H9N2 viruses with novel genotypes.
PDF Abstract XML References Citation
Received: May 20, 2011;
Accepted: September 14, 2011;
Published: September 28, 2011
How to cite this article
Masoud Soltanialvar and Ali Bagherpour, 2011. Characterization of PB2 Gene of H9N2 Avian Influenza Viruses from Iran, 2008 to 2009. Journal of Biological Sciences, 11: 340-344.
DOI: 10.3923/jbs.2011.340.344
URL: https://scialert.net/abstract/?doi=jbs.2011.340.344
DOI: 10.3923/jbs.2011.340.344
URL: https://scialert.net/abstract/?doi=jbs.2011.340.344
INTRODUCTION
Influenza viruses are members of the family Orthomyxoviridae with a genome of single-stranded negative-sense RNA composed of 8 gene segments encoding at least 10 proteins (Lee and Saif, 2009). These viruses are classified into three major types A, B, C based on the antigenic differences in their nucleoprotein and matrix protein. The type A virus is pleomorphic and spherical (approximately 120 nm in diameter) and can be further classified into subtypes according to the antigenicity of two surface glycoproteins Hemagglutinin (HA) and Neuraminidase (NA) (Bouvier and Plese, 2008). To date, 16 HA subtypes and 9 NA subtypes of influenza A virus have been identified (Fouchier et al., 2005).The eight segments of the influenza virus RNA genome are packaged into Ribonucleoprotein Particles (RNPs) containing the Nucleoprotein (NP) and the trimeric RNA-dependent RNA polymerase complex which comprises subunits PA, PB1 and PB2. The polymerase operates in two distinct modes: in the first, it transcribes virally encoded genes, using a cap-snatching mechanism to prime transcription and ensure proper 5' capping of viral messenger RNA; in the second, it replicates full-length viral RNA to produce first positive-strand complementary RNA and then progeny viral RNA. The PB1 subunit binds the conserved 5' and 3' ends of the viral RNA and carries both the polymerase active site and the endonuclease activity that cleaves hostcell mRNA bound by the cap-binding PB2 subunit. PB1 residues implicated in the endonuclease and polymerase active sites have been identified, although the location of the cap-binding site of PB2 remains controversial (Bouvier and Plese, 2008).
The H9N2 subtype that has spread worldwide in poultry could infect humans and is considered to be one of the potential public health risks with possible emergence of a pandemic virus (Butt et al., 2005). In this study, we characterized PB2 genes and proteins of five Iranian isolates which have been isolated from commercial broiler chicken in the Iran between 2008 and 2009. These isolates resulted in rapid mortality due to tracheitis and respiratory congestion. We delineated the PB2 gene of these field isolates and we also established their phylogenetic relationship to the other Asian H9N2 viruses.
MATERIALS AND METHODS
Samples and virus isolation: Samples from various parts of the country (30 farms) were collected between April 2008 and February 2009.
Sample collection was performed according to the standard method for clinical poultry specimens (Swayne et al., 1998). Lung and trachea specimens were stored at -70°C until used.
They were treated with 2xphosphate buffer solution (PBS, pH 7.4) containing antibiotics and antifungals (Penicillin 10000 unit mL-1, Streptomycin 10000 unit mL-1 and Nystatin 20000 unit mL-1). Initial viral isolation was performed in 10-day-old SPF (Specific Pathogen Free) Embryonated Chicken Eggs (ECEs). Eggs were candled daily and embryos dying within 24 h Post Inoculation (PI) were discarded. Allantoic fluids were collected from the eggs and the presence of viruses was determined by haemagglutination. Subtype identification of the viruses was determined by standard Haemagglutination-inhibition (HI) and Neuraminidase-inhibition (NI) tests using polyclonal chicken antisera as described by Alexander and Spackman (1981).
Five virus isolates obtained in this study were named as follows:
| • | A/chicken/Iran/RZ28/2008 |
| • | A/chicken/Iran/RZ36/2008 |
| • | A/chicken/Iran/RZ37/2008 |
| • | A/chicken/Iran/RZ42/2009 |
| • | A/chicken/Iran/RZ53/2008 |
RT-PCR and sequence analysis: The viral RNA was extracted directly from the allantoic fluid by means of the High pure viral Nucleic Acid Kit (Roche Germany). Purified genomic RNA was used to generate cDNA clones by (RT-PCR) according to the standard procedure (Lee et al., 2001). RT-PCR was performed by using specific primers for 2262 bp fragment of PB2.
Primers used for PB2 amplification were:
| • | Forward primer (2262 bp): 5'- GCAAAAGCAGGAG TGAAAATG-3' |
| • | Reverse primer (2262 bp): 5'- AGTCCTGAGCACAA ATAACTGG-3' |
The PCR products were purified by using High pure product purification kit (Roche Germany). PCR products were applied to low melting point agarose (LMP) and the distinct bands were purified from gel for sequencing (MWG co, Germany).
Nucleotide and deduced amino acid sequences of the PB2 gene were edited with the Editseq (DNASTER Software package Version 5.2 (Nucleotide and deduced amino acid sequences were aligned by ClustalW, Version 1.4.
Nucleotide sequences of the PB2 gene were used for phylogenetic tree construction. The phylogenetic analysis was performed with the MegAlign program 2.8.
RESULTS
In this study, 2262 base pairs of the PB2 genes were sequenced and 754 amino acid sequences from five isolates of the PB2 genes were deduced from the nucleotide sequences.
The Iranian isolates did not exhibit insertions or deletions within (PB2) gene as compare with their prototype A/turkey/winconsin/66 but rather numerous point mutations were registered.
These viruses characterized by 8 amino acid substitutions at the following positions: 7 (V to I), 270 (S to C), 271 (T to A) 280 (N to S), 285 (R to K)3 301) (H to k), 416 (M to L) compared with other viruses reported.
Phylogenetic analysis of the PB2 gene showed that all the PB2 genes of the Iranian H9N2 virus fell in two groups, unknown avian and Dk1 (Fig. 1). All of the H9N2 viruses isolated in 2008-2009 except A/chicken/Iran/RZ53/2008 belonged to the unknown avian sublineage which grouped with the 2004 Pakistani H7N3 viruses. A/chicken/Iran/RZ53/2008 clustered with Dk2 sublineage which are most closely related to Dk/ST/163/04, it isolated possibly from migratory duck.
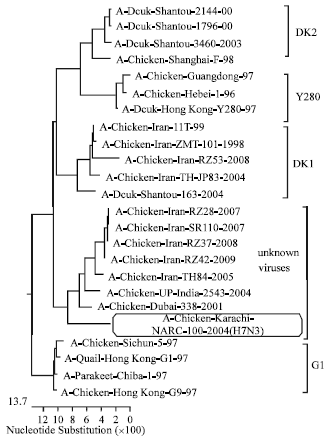 | |
| Fig. 1: | Phylogenetic analysis of the PB2 gene showed that all the PB2 genes of the Iranian H9N2 viruse fell in two groups, unknown avian and Dk1 |
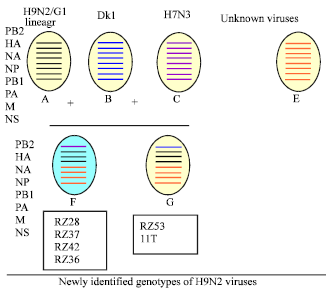 | |
| Fig. 2: | Phylogenetic analysis of the Iranian PB2 gene revealed two different genotypes |
Based on sequence comparison and our previous studies (Soltanialvar et al., 2010, 2011), we identified at least two different genotypes, designated F and G, among these five viruses (Fig. 2).
The PB2 polymerase genes of the recent H9N2 viruses originated from 2 sublineages. The PB2 gene of A/chicken/Iran/RZ53/2008 (genotype G) belonged to the Dk2 sublineage. However, most PB2 genes of the H9N2 viruses isolated in 2008-2009 (genotypes F) were similar to those of the H7N3 viruses isolated in the Pakistan.
A number of residues in the polymerase proteins (PB1, PB2 and PA) are known to play a key role in the host range of avian influenza viruses to increase virulence or replication in the mammalian host. All Iranian isolates carry amino acid substitution A717L in the PB2 gene, a mutation that correlates with the adaptation of H9N2 avian influenza virus to mice and humans.
DISCUSSION
H9N2 viruses circulated widely in the Middle East (Iran, United Arab Emirates Israel)and were associated with serious disease in poultry (Aamir et al., 2007; Alexander, 2007; Mosleh et al., 2009; Perk et al., 2009). In this study, we have reported the first genetic analysis of PB2 gene of H9N2 avian influenza viruses and found that Iranian viruses had undergone genetic reassortment.
The molecular basis of host-range restriction and adaptation of influenza A viruses to a new host species has not yet been defined. Previous studies suggested that mutation of the polymerase complex is required for adaptation to a new host and may enhance replication and transcription of the adapted virus in a mammalian host (Gabriel et al., 2005; Wu et al., 2009).
Earlier work focused on the role of the HA and M1 proteins in host-range restriction and adaptation (Govorkova et al., 2000; Wan and Perez, 2007). But numerous studies now indicate that the virulence of influenza viruses is likely to be a multigenic trait (Chen et al., 2007; Gabriel et al., 2005). Amino acid 717 of PB2 is a known determinant of pathogenicity and host specificity. Shinya et al. (2007) indicated that the PB2 A717L substitution may be crucial for adaptation to mammalian hosts. We suggested that the substitution PB2 A717L observed here in Iranian viruses may also lead to increase pathogenicity. The PB2 substitution A717L was located between the second Nuclear Localization Signal (NLS2) at 736-739 and the cap-binding motif at 634-650: it is possible that the mutation affects the activity of one or both of these signals.
Previous studies have shown that the Eurasian lineage consists of at least three sublineages represented by their prototype strains: A/chicken/Korea/38349-p96323/96 (Korean-like), A/duck/Hong Kong/Y280/97 (Y280-like) and A/quail/Hong Kong/G1/97 (G1-like) (Guan et al., 2000; Matrosovich et al., 2001). As reported by Xu et al. (2007), our result also showed that PB2 gene of H9N2 viruses formed different sublineages including G1-like, Ck/Bei-like, three duck lineages (Dk1, Dk2, Dk3) and unknown avian.
Our previous studies (Soltanialvar et al., 2010, 2011) indicated that Iranian surface glycoprotein genes (HA and NA) and one internal genes (NP) are similar to G1-like virus represented by Qa/HK/G1/97, whereas the PB2 genes of the Iranian H9N2 viruses, formed a distinct group compared to G1-, Korean- and Y280-like sublineage.
Comparison of the extent of PB2 gene sequence homologies of the Iranian isolates showed more similarity with a H7N3 chicken isolate from Pakistan (A/Chicken/ Karachi/NARC-100/2004; 91.4-93.1%) compared to Qa/HK/G1/97 (85.3-86.6%), Dk/HK/Y280/97 (84.7-86.9%) and Ck/Korea/323/96 (88.2-89.9%). Based on the genetic similarities and phylogenetic analysis, our findings suggested that the Iranian viruses had undergone genetic reassortment with other influenza subtypes including H7 viruses.
Like the Iranian isolates, reassortment between H9N2 and the highly pathogenic avian influenza virus H7N3 subtype was reported in Pakistan (Abbas et al., 2010). It is also noted that the viruses from Dubai and India shared an out group relationship with the Iranian viruses in the PB2 gene tree suggesting that these viruses are derived from the same gene pool.
Phylogenetic analysis of the Iranian PB2 gene revealed two different genotypes. Our identification of novel genotypes of H9N2 viruses in 2008-2009 was markedly similar to those of a recent study conducted by Iqbal et al. (2009) in Pakistan. This finding suggested that a high degree of diversity among the H9N2 viruses in the regions of Middle East and Indian sub-continent.
In subsequent years, multiple reassortment variant genotypes of H9N2 avian influenza viruses from domestic poultry in China and India have been identified and well characterized (Wu et al., 2008; Tosh et al., 2008; Sun et al., 2010). The association of high mortality in recent years and report of H5N1 and H9N2 in wild birds in Iran (Shoushtari et al., 2008) raised the probability of a new genetic modified avian influenza virus.
Homayounimehr et al. (2010) and Langeroudi et al. (2008) have shown that the Iranian isolates possessed amino acid Leucine (L) at position 226 instead of glutamine (Q) at the receptor binding site of haemagglutinins (HA) which is similar to A/Quail/HongKong/G1/97 and the 2 human isolates A/HK/1073/99, A/HK/1074/99 (Homayounimehr et al., 2010; Langeroudi et al., 2008). Amino acid differences in the receptor binding sites of HAs have been shown to be associated with differences in receptor binding specificity (Wan and Perez, 2007). So, Iranian H9N2 isolates can bind to α(2, 6) receptors, this feature suggested the pandemic potential of the H9N2 avian influenza virus and emphasizes the need for continuous surveillance in Iran which has been continuing since 2000.
CONCLUSION
Our findings demonstrated that the H9N2 viruses in Iran underwent extensive reassortment which led to generation with novel genotypes. The emergence of these novel genotypes of H9N2 viruses and the sustained prevalence of these viruses in poultry warrant further surveillance of H9N2 viruses by complete genomic analysis.
ACKNOWLEDGMENTS
This study was supported by Islamic Azad University, Shoushtar Branch. The authors thank the excellent technical support provided by Miss Eshratabadi.
REFERENCES
- Aamir, U.B., U. Wernery, N. Ilyushina and R.G. Webster, 2007. Characterization of avian H9N2 influenza viruses from United Arab Emirates 2000 to 2003. Virology, 361: 45-55.
CrossRefPubMedDirect Link - Abbas, M.A., E. Spackman, D.E. Swayne, Z. Ahmed and L. Sarmento et al., 2010. Sequence and phylogenetic analysis of H7N3 avian influenza viruses isolated from poultry in Pakistan 1995-2004. Virol. J., 7: 137-146.
CrossRef - Alexander, D.J., 2007. An overview of the epidemiology of avian influenza. Vaccine, 25: 5637-5644.
CrossRef - Alexander, D.J. and D. Spackman, 1981. Characterization of influenza A viruses isolated from turkeys during March-May 1979. Avian Pathol., 10: 281-293.
PubMed - Bouvier, N.M. and P. Palese, 2008. The biology of Influenza viruses. Vaccine, 56: D49-D53.
Direct Link - Butt, K.M., G.J. Smith, H. Chen, L.J. Zhang and Y.H. Leung et al., 2005. Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. J. Clin. Microbiol., 43: 5760-5767.
PubMed - Chen, H., R.A. Bright, K. Subbarao, C. Smith, N.J. Cox, J.M. Katz and Y. Matsuoka, 2007. Polygenic virulence factors involved in pathogenesis of 1997 Hong Kong H5N1 Influenza viruses in mice. Virus Res., 128: 159-163.
CrossRefPubMedDirect Link - Fouchier, R.A.M., V. Munster, A. Wallensten, T.M. Bestebroer and S. Herfst et al., 2005. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J. Virol., 79: 2814-2822.
CrossRefDirect Link - Gabriel, G., B. Dauber, T. Wolff, O. Planz, H.D. Klenk and J. Stech, 2005. The viral polymerase mediates adaptation of an Avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci., 102: 18590-18595.
CrossRef - Govorkova, E.A., A.S. Gambaryan, E.C. Claas and Y.A. Smirnov, 2000. Amino acid changes in the hemagglutinin and matrix proteins of influenza a (H2) viruses adapted to mice. Acta Virol., 44: 241-248.
PubMed - Guan, Y., K.F. Shortridge, S. Krauss, P.S. Chin and K.C. Dyrting et al., 2000. H9N2 influenza viruses possessing H5N1-like internal genomes continue to circulate in poultry in Southeastern China. J. Virol., 74: 9372-9380.
CrossRefDirect Link - Homayounimehr, A.R., H. Dadras, A. Shoushtari and S.A. Pourbakhsh, 2010. Sequence and phylogenetic analysis of the haemagglutinin genes of H9N2 avian influenza viruses isolated from commercial chickens in Iran. Trop. Anim. Health Prod., 42: 1291-1297.
CrossRef - Iqbal, M., T. Yaqub, K. Reddy and J.W. McCauley, 2009. Novel genotypes of H9N2 influenza A viruses isolated from poultry in Pakistan containing NS genes similar to highly pathogenic H7N3 and H5N1 viruses. PLoS ONE, Vol. 4.
CrossRefDirect Link - Langeroudi, A.G., V. Karimi, M.T. Kheiri, F. Mahboudi and A. Barin, 2008. Nucleotide and amino acid sequence analysis of hemagglutinin protein in cleavage site region of H9N2 isolated from broilers in Tehran province during 1998-2007. J. Anim. Vet. Adv., 7: 529-534.
Direct Link - Lee, C.W. and Y.M. Saif, 2009. Avian influenza virus. Comp. Immunol. Microbiol. Infec. Dis., 32: 301-310.
CrossRef - Lee, M.S., P.C. Chang, J.H. Shien, M.C. Cheng and S.K. Shieh, 2001. Identification and subtyping of avian influenza viruses by reverse transcription PCR. J. Virol. Methods, 97: 13-22.
Direct Link - Matrosovich, M.N., S. Krauss and R.G. Webster, 2001. H9N2 influenza A viruses from poultry in Asia have human virus-like receptor specificity. Virology, 281: 156-162.
CrossRefPubMedDirect Link - Mosleh, N., H. Dadras and A. Mohammadi, 2009. Molecular quantitation of H9N2 avian influenza virus in various organs of broiler chickens using TaqMan real time PCR. J. Mol. Genet Med., 3: 152-157.
Direct Link - Perk, S., N. Golender, C. Banet-Noach, E. Shihmanter and S. Pokamunsky et al., 2009. Phylogenetic analysis of hemagglutinin, neuraminidase, and nucleoprotein genes of H9N2 avian influenza viruses isolated In Israel during the 2000-2005 epizootic. Comp. Immunol. Microbiol. Infec. Dis., 32: 221-238.
PubMed - Shinya, K., S. Watanabe, T. Ito, N. Kasai and Y. Kawaoka, 2007. Adaptation of an H7N7 equine influenza A virus in mice. J. Gen. Virol., 88: 547-553.
CrossRef - Shoushtari, A., M.H. Hablolvarid, M. Vascellari and A. Hedayati, 2008. Mortality of wild swans associated with naturally infection with highly pathogenic H5N1 avian influenza virus in Iran. Arch Razi Inst., 62: 207-213.
Direct Link - Soltanialvar, M., H. Shoushtari, M. Bozorgmehrifard, S. Charkhkar and F. Eshratabadi, 2010. Molecular characterization of hemagglutinin and neuraminidase genes of H9N2 avian influenza viruses isolated from commercial broiler chicken in Iran. J. Biol. Sci., 10: 145-150.
CrossRef - Sun, Y., J. Pu, Z. Jiang, T. Guan and Y. Xia et al., 2010. Genotypic evolution and antigenic drift of H9N2 influenza viruses in China from 1994 to 2008. Vet Microbio., 146: 215-225.
PubMed - Swayne, D.E., J.R. Glisson, M.W. Jackwood, J.E. Pearsson and W.M. Reed, 1998. A Laboratory Manual for the Isolation and identification of Avian Pathogens. 4th Edn., American Association of Avian Pathologist, Pennsylvania, USA., ISBN-13: 9780915538072, Pages: 311.
Direct Link - Wan, H. and D.R. Perez, 2007. Amino Acid 226 in the Hemagglutinin of H9N2 Influenza Viruses Determines Cell Tropism and Replication in Human Airway Epithelial Cells. J. Virol., 81: 5181-5191.
PubMed - Wu, R., H. Zhang, K. Yang, W. Liang and Z. Xiong et al., 2009. Multiple amino acid substitutions are involved in the adaptation of H9N2 vian influenza virus to mice. Vet Microbiol., 138: 85-91.
PubMed - Wu, R., Z.W. Sui, H.B. Zhang, Q.J. Chen and W.W. Liang et al., 2008. Characterization of a pathogenic H9N2 influenza A virus isolated from central China in 2007. Arch. Virol., 153: 1549-1555.
PubMed - Xu, K.M., G.J.D. Smith, J. Bahl, L. Duan and H. Tai et al., 2007. The genesis and evolution of H9N2 influenza viruses in poultry from Southern China, 2000 to 2005. J. Virol., 81: 10389-10401.
CrossRefPubMedDirect Link