
K. Khucharoenphaisan
Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok 10220, Thailand
U. Puangpetch
Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok 10220, Thailand
K. Puttaraksa
Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok 10220, Thailand
K. Sinma
Department of Applied Biological Chemistry, Faculty of Agriculture, Shizuoka University, 836 Ohya, Shizuoka 422-8529, Japan
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 4 | Page No.: 314-319







ABSTRACT
Actinomycetes are a group of prokaryotic organisms belonging to Gram-positive bacteria and play an important ecological role in recycling substances in the nature. To determine possible established correlation between isolated actinomycetes and its biochemical degradation (xylan, soluble starch, cassava, protein, lipid, uric acid, carboxymethyl cellulose and avicel degradation) and oxidation property (guaiacol), the actinomycetes were isolated from Termes sp. The dendogram was generated from UPGMA analysis with FreeTree software. In this work, 45 strains of actinomycetes were isolated from guts of Termes sp. Among these, 44 isolated strains could degrade protein, soluble starch and cassava starch. Twenty-three strains degraded uric acid and xylan. The isolated strains that able to degrade avicel and oxidize lignin were rare. The morphological character showed the variety of aerial hyphae and spore forming in each strain. The dendogram was constructed based on biodegradation activity of tested strains. The tested strains were classified into 5 clusters. Cluster 1, cluster 2, cluster 3, cluster 4 and cluster 5 contained 14, 9, 9, 9 and 4 isolated strains, respectively. The actinomycetes strains showed a similar biodegration activity within a same cluster. This result indicated the relation between biodegradation activity and actinomycetes strains. Dendogram based on the biodegradation activity was found to be an efficiencient tool for grouping purpose.
PDF Abstract XML References Citation
Received: June 22, 2011;
Accepted: August 08, 2011;
Published: September 28, 2011
How to cite this article
K. Khucharoenphaisan, U. Puangpetch, K. Puttaraksa and K. Sinma, 2011. Grouping of Actinomycetes Isolated from Termites using Biochemical Character. Journal of Biological Sciences, 11: 314-319.
DOI: 10.3923/jbs.2011.314.319
URL: https://scialert.net/abstract/?doi=jbs.2011.314.319
DOI: 10.3923/jbs.2011.314.319
URL: https://scialert.net/abstract/?doi=jbs.2011.314.319
INTRODUCTION
Termites are important insects in terrestrial ecosystems, playing a role in the decomposition of wood or plant litter (Robert et al., 2007). They harbor an abundance and diversity of gut bacteria, which are thought to play essential roles in the carbon and nitrogen metabolism of their host termites (Ramin et al., 2008a). The dense microbial communities in their guts, consisting of both flagellated protists (eukaryotes) and prokaryotes, facilitated the digestion to their hosts (the termites). The relationship between termites and their gut microorganisms is a well-known example of symbiosis. Termite bacteria are also involved in the degradation of the cellulose, hemicelluloses, starch and aromatic compounds as well as nitrogen fixation and lignocellulose decomposition (Femi-Ola and Olowe, 2011; Ramin et al., 2008b; Ramin et al., 2008c; Varma et al., 1994; Konig et al., 2006; Li et al., 2006). The relationship between termites and the microorganisms enables termites to feed on lignocelluloses. However, culture-independent molecular sequence studies reveal that the majority of microbes in the gut are yet to be characterized (Leadbetter et al., 1999) which limits our understanding of the symbiosis. Filamentous prokaryotic organisms, putatively actinobacteria have been frequently described in termite guts since the detection of Micromonospora species. Since the late 1970s, the culture of Steptomyces followed other successful culturing attempted for Arthrobacter, Aureobacterium, Cellulosimicrobium, Cellulomonas, Kocuria, Microbacterium, Micrococcus, Rhodococcus and Norcadia species (Kuhnigk et al., 1994; Schafer et al., 1996; Kuhnigk and Konig, 1997; Bakalidou et al., 2002; Wenzel et al., 2002). Although, microscopically evidence suggests that actinobacteria clusters are not simply accidental occupants of the intestinal lumen, but intimately associated with the absorptive epithelia (Konig et al., 2006) still information on these organisms in culture is incomplete.
Dendogram is a tree diagram that frequently used to illustrate the arrangement of the clusters of genes or samples (Al-Soqeer, 2011; Chaveerach et al., 2008). The identification of a large number of isolated actinomycetes needed high cost and much time. Grouping method using dendogram is a choice to reduced and save processing cost. In this work, actinomycetes from termite has been isolated and studied on their biodegradation activities against various substrates. Among the isolated actinomycetes, a variety of biodegradation was found. One strain has ability to degrade more than one substance. To handle with large number of actinomycetes strains which have different biodegradation ability, grouping in cluster in order to reduce a number of samples was necessary. The dendogram was constructed by using its biochemical properties and statistical program for grouping purpose. Moreover, morphological properties of representative strains were also characterized.
MATERIALS AND METHODS
This research project was conducted from September 2010 to June 2011 at Faculty of Science and Technology, Phranakhon Rajabhat University, Thailand.
Isolation and screening of actinomycetes: Actinomycetes were isolated from gut of termites in genus of Termes that was collected from Sakaerat Environmental Research Station in Nakhon Ratchasima province, Thailand. Each sample was grinded and diluted to go on serial dilution and plate on humic acid vitamin agar (Hayakawa and Nonomura, 1987) supplemented with 50 mg L-1 of cyclohexamide and 20 mg L-1 of nalidixic acid. The isolation plates were incubated at room temperature for 30 days. The number of total actinomycetes was evaluated after the colonies that appeared on the plates. The colonies had been transferred to yeast extract-malt extract (ISP medium No. 2) agar (Shirling and Gottlieb, 1966; Khucharoenphaisan and Sinma, 2011) plates for purity check.
Degradation and oxidation properties: Xylan, carboxymethylcellulose (CMC) and avicel degradation were tested on agar medium containing 10 g L-1 beechwood xylan or CMC or avicel; 5 g L-1 peptone; 3 g L-1 yeast extract; 3 g L-1 malt extract; 2 g L-1 KH2PO4; 0.3 g L-1 MgSO4·7H2O; 0.3 g L-1 CaCl2 and 18 g L-1 agar. The degradation was evaluated from the clear zone formation around colony after 30 days of inoculation. Uric acid degradation was observed on modified ISP medium No. 2 containing 10 g L-1 malt extract; 4 g L-1 glucose; 10 g L-1 uric acid and 18 g L-1 agar. Uric acid degradation was evaluated after 3 days of inoculation from the clear zone formation around colony. Lignin degradation was observed on modified ISP medium No. 2 containing 10 g L-1 malt extract; 4 g L-1 glucose; 0.01% guaiacol as lignin model compound and 18 g L-1 agar. The positive on oxidation of guaiacol was indicated by red brown zone around the colony after 30 days of inoculation. Cassava and soluble starch degradation were tested on agar medium containing 5 g L-1 soluble starch or cassava starch; 5 g L-1 peptone; 5 g L-1 yeast extract; 5 g L-1 KH2PO4; 0.5 g L-1 MgSO4•7H2O and 18 g L-1 agar. The degradation was observed after 3 days of point inoculation by flooding of iodine solution for 15 min. All of degradation properties were done in petri dish at room temperature.
Morphological characteristic: The morphological characters of all isolated strains were studied on the colors of mature substrate mycelium, aerial mycelium, spore mass, and diffusible pigment by using crosshatch streak technique (Shirling and Gottlieb, 1966). The fine structures of actinomycetes such as the number of spores at the end of mature hyphae, the form of the spore chain and spore bearing hyphae and other special morphological were determined on humic acid vitamin agar plate under light microscope by using slide culture technique after 30 days of cultivation.
Dendogram construction: Polymorphic were identified using hydrolysis properties. The hydrolysis was scored as positive (1) or negative (0). The pair-wise evolutionary distances are given by generated the distance matrix (Saitou and Nei, 1987). The dendogram was constructed from the evolutionary distance data obtained to the algorithm of the UPGMA and Jaccard method (Cenkci et al., 2008). Free Tree (Pavlicek et al., 1999) and Tree View (Page, 1996) software package were used to construct the dendogram for comparison.
RESULTS AND DISCUSSION
Isolation of actinomycetes and compound degradation: Forty-five actinomycetes were isolated from termite. All isolated strains were able to degrade lipid compound. They were 44 isolated strains could degrade protein, soluble starch and cassava starch. Twenty-three of all degraded uric acid and xylan. The isolated actinomycetes were able to degrade avicel and oxidize lignin only 2 and 1 isolate, respectively. This result revealed that most of actinomycetes isolated from termite were able to degrade lipid, protein, soluble starch and cassava starch. Uric acid, xylan and CMC were half of those. The clear zone was shown in Fig. 1. The positive result from uric acid (Fig. 1a), protein (Fig. 1c), soluble starch (Fig. 1d), cassava starch (Fig. 1e), avicel cellulose (Fig. 1f), carboxymethylcellulose (Fig. 1g) and beechwood xylan (Fig. 1h) degradation were detected from the appearance of clear zone around colony.
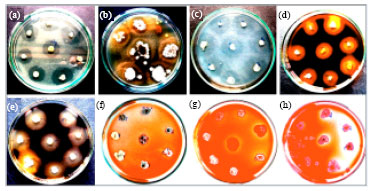 | |
| Fig. 1 (a-h): | Degradation of (a) uric acid, (b) lipid, (c) protein, (d) soluble starch, (e) cassava starch, (f) avicel cellulose, (g) carboxymethylcellulose and (h) beechwood xylan |
 | |
| Fig. 2 (a-i): | Light microscopic micrograph of isolated strains; (a) R2, (b) R8, (c) R42, (d) R51, (e) R61, (f) R62, (g) R66, (h) R69 and (i) R99 showing structure of spore chains and surface spores after 4 weeks growth at 30°C on humic acid vitamin agar |
In contrast, the degradation of lipid (Fig. 1b) was appearing as accumulation of white particle around colony.
Some microorganism was isolated from termites’ gut and has xylanolytic activity such as Paenibacillus (Ohkuma, 2003). Beside the xylanolytic activities, actinomycetes isolated from termites’ guts also have ability to oxidizing uric acid, the nitrogenous waste from purine degradation of termite. This compound was found in small amount in fecal of termite and suppose that gut bacteria recycle uric acid nitrogen in termites, uric acid is serve as nitrogen source for termites (Potrikus and Breznak, 1981). When consider the relationship of termite food-source and degradation ability of actinomycetes, the result indicated that food source of termite not affected to the distribution and degradation ability of actinomycetes habitat in termites’ guts.
Morphological characteristic: Isolated strains were an aerobic, Gram-positive, non-acid alcohol-fast actinomycetes that forms extensively branched substrate mycelia. Our isolated strains have morphological character under microscopic and agar slant as shown in Fig. 2. Strain R2 produced white color aerial mycelium and produce two spores arise from aerial mycelium (Fig. 2a). Strain R8 produced light brow aerial mycelium with flexibile spore chain with 16 spores (Fig. 2b). Light orange soluble pigment was produced. Strain R42 produced light orange aerial mycelium with spiral spore type containing 20-30 spores (Fig. 2c). Strain R51 produced light brown aerial mycelium and rectiflexible-type spores chain containing 20-40 spores (Fig. 2d). Strain R62 produced brown aerial mycelium with longitudinal spirales-type spore chain (Fig. 2f).
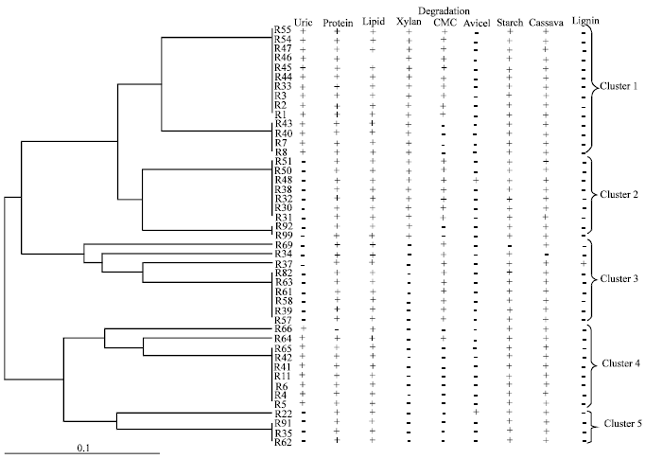 | |
| Fig. 3: | Dendogram indicating relationships of actinomycetes isolated from guts of Termes sp. with degradation activity. UPGMA and Jaccard method are given as dendogram using Free Tree and Tree View software |
Strain R61, R69 and R66 produced light brown to brown aerial mycelium with short spore-chain with hook-like structure as shown in Fig. 2e, h and g, respectively. Strain R99 produced light brown aerial mycelium with dense spore mass (Fig. 2i). The spore chain is a spirales type with compact longitudinal chain.
Grouping of actinomycetes strains based on its degradation ability: Forty-five strains of actinomycetes were isolated from Termes sp. The degradation abilities of each strains were tested on uric acid, protein, lipid, xylan, CMC, avicel cellulose, starch, cassava and lignin compound. The result indicated that the microorganisms have a wide variety of degradation abilities. One strain can degrade various substrate. In this study, we use the dendogram to group the actinomycetes strain by using its ability to degrade test substrate.
Based on the degradation ability, the tested strains were classified into 5 clusters as shown in the dendogram (Fig. 3). The first cluster is large containing 14 strains of actinomycetes. Actinomycetes in this cluster had a wider potential to degrade tested substances that are uric acid, protein, xylan, CMC, starch and cassava but lack the ability to degrade avicel and lignin compound. Among these cluster, the actinomycetes were separated into 2 groups. The minor group contained 4 strains and lack the activity of CMC degradation.
The second cluster consisted of 9 strains which lack of uric acid degradation activity when compare to the first cluster. According to first cluster, the second cluster also separated to 2 branches. Two strains that are R92 and R99 were lack of CMC degradation ability and separated from major branch of these cluster.
The third cluster was separated from the second cluster because them lack of xylan degradation activity. Among this cluster, three strains were formed a differentiate branches from major branch. Strain R69 and R34 formed a differentiate branch by lack of starch degradation and cassava degradation activity, respectively, while strain R37 was separated from major branch because it had lignin degradation activity.
Cluster 4 and 5 were separated from the third cluster by lack of CMC degradation ability. However, among these 2 clusters the actinomycetes were separated by uric acid degradation activity. It seem to be that cluster 4 have uric acid degradation activity while cluster 5 lack its.
From the dendogram of degradation activity, it was found that all actinomycetes strain which was isolated from Termes sp. have a common ability to degrade lipid and most strains can degrade protein, starch and cassava. In contrast, only 1 strain can degrade lignin model compound and 2 strain can degrade avicel. The degradation of uric acid, xylan and CMC were varies and used as the key factors for the grouping of microorganisms by using a dendogram. Our result was similar to Kanmegne et al. (2010) who generated dendogram of Garcinia kola seeds in to cluster by using biochemical parameters such as phenol, soluble carbohydrate, protein and amino acid contents. Furthermore, Khucharoenphaisan et al. (2009) constructed a dendogram from RAPD band and found relationship between genotype and xylanase production. Electrophoretic proteins of seed cotyledons were also used for effective method for grouping (Emre et al., 2006, 2007).
At present, the dendogram was consider as a tool for the study of relationship between the microorganism and used for an application in various field such as molecular genetic, taxonomy etc. From the best of our knowledge, there are no report about the application of dendogram for the relation of biodegradation.
CONCLUSION
Forty-five strains of actinomycetes were isolated from Termes sp. They have a wide variety of degradation abilities such as uric acid, protein, lipid, xylan, CMC, avicel cellulose, starch, cassava and lignin compound. From the above results the construction of dendogram based on the biodegradation activity was an efficience tool for grouping purpose. This method was convinian and helpful for management and selection of the suitable strain carrying an appropriate enzyme activity.
ACKNOWLEDGMENT
This research was supported by Institute of Research and Development Phranakhon Rajabhat University, Thailand.
REFERENCES
- Al-Soqeer, 2011. Genotypic diversity among wild populations of buffelgrass (Cenchrus ciliaris L. Link) in Al-Qassim region. Asian J. Biotechnol., 3: 262-268.
CrossRefDirect Link - Bakalidou, A., P. Kampfer, M. Berchtold, T. Kuhnigk, M. Wenzel and H. Konig, 2002. Cellulosimicrobium variabile sp. nov., a cellulolytic bacterium from the hindgut of the termite Mastotermes darwiniensis. Int. J. Syst. Evol. Microbiol., 52: 1185-1192.
CrossRefDirect Link - Cenkci, S., M. Yildiz, M. Konuk and Y. Eren, 2008. RAPD analyses of some wild Triticum L. and Aegilops L. species and wheat cultivation in Turkey. ACTA Biologica. Cracoviensia Series Botanica, 50: 35-42.
Direct Link - Chaveerach, A., N. Srisamoot, S. Nuchadomrong, N. Sattayasai, P. Chaveerach, A. Tanomtong and K. Pinthong, 2008. Phylogenetic relationships of wildlife order Carnivora in Thailand inferred from the internal transcribed spacer region. J. Biol. Sci., 8: 278-287.
CrossRefDirect Link - Robert, O.E., U.O. Frank and O.U. Agbonsalo, 2007. Influence of activities of termites on some physical and chemical properties of soils under different land use patterns: A review. Int. J. Soil Sci., 2: 1-14.
CrossRefDirect Link - Emre, I., D. Turgut-Balik, H. Genc and A. Sahin, 2006. The use of seed proteins revealed by SDS-PAGE in taxonomy of some Lathyrus L. species grown in Turkey. Pak. J. Biol. Sci., 9: 2358-2361.
CrossRefDirect Link - Emre, I., D. Turgut-Balik and A. Sahin, 2007. Electrophoretic analysis of total protein profiles of some Lathyrus L. (Sect. Cicercula) grown in Turkey. Pak. J. Biol. Sci., 10: 2890-2894.
CrossRefPubMedDirect Link - Femi-Ola, T.O. and B.M. Olowe, 2011. Characterization of α amylase from Bacillus subtilis BS5 isolated from Amitermes evuncifer silvestri. Res. J. Microbiol., 6: 140-146.
CrossRefDirect Link - Hayakawa, M. and H. Nonomura, 1987. Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J. Ferment. Technol., 65: 501-509.
CrossRefDirect Link - Kanmegne, G., H.D. Mbouobda, B. Temfack, E.K. Koffi and D.N. Omokolo, 2010. Impact of biochemical and morphological variations on germination traits in Garcinia kola heckel seeds collected from cameroon. Res. J. Seed Sci., 3: 82-92.
CrossRef - Khucharoenphaisan, K., S. Tokuyama, K. Ratanakhanokchai and V. Kitpreechavanich, 2009. A comparative study of Thermomyces lanuginosus strains on thermostable xylanase production. Afr. J. Biotechnol., 8: 1608-1614.
Direct Link - Khucharoenphaisan, K. and K. Sinma, 2011. Production and partial characterization of uric acid degrading enzyme from new source Saccharopolyspora sp. PNR11. Pak. J. Biol. Sci., 14: 226-231.
CrossRefDirect Link - Kuhnigk, T., E.M. Borst, A. Ritter, P. Kampfer, A. Graf, H. Hertel and H. Konig, 1994. Degradation of lignin monomers by the hindgut flora of xylophagous termites. J. Syst. Applied Microbiol., 17: 76-85.
CrossRefDirect Link - Kuhnigk, T. and H. Konig, 1997. Degradation of dimeric lignin model compounds by aerobic bacteria isolated from the hindgut of xylophagous termites. J. Basic Microbiol., 37: 205-211.
CrossRefDirect Link - Leadbetter, J.R., T.M. Schmidt, J.R. Graber and J.A. Breznak, 1999. Acetogenesis from H2 plus CO2 by spirochetes from termite guts. Science, 283: 686-689.
CrossRefDirect Link - Ohkuma, M., 2003. Termite symbiotic systems: Efficient bio-recycling of lignocellulose. Applied Microb. Biotechnol., 61: 1-9.
CrossRefDirect Link - Page, R.D.M., 1996. TreeView: An application to display phylogenetic trees on personal computers. Comput. Applied Biosci., 12: 357-358.
CrossRefPubMedDirect Link - Pavlicek, A., S. Hrda and J. Flegr, 1999. Free-Tree-freeware program for construction of phylogenetic trees on the basis of distance data and bootstrap/jackknife analysis of the tree robustness. Application in the RAPD analysis of genus Frenkelia. Folia Biol. (Praha), 45: 97-99.
PubMedDirect Link - Potrikus, C.J. and J.A. Breznak, 1981. Gut bacteria recycle uric acid nitrogen in termites: A strategy for nutrient conservation. Proc. Nat. Acad. Sci., 78: 4601-4605.
Direct Link - Ramin, M., A.R. Alimon, N. Abdullah, J.M. Panandam and K. Sijam, 2008. Isolation and identification of three species of bacteria from the termite Coptotermes curvignathus (Holmgren) present in the Vicinity of University Putra Malaysia. Res. J. Microbiol., 3: 288-292.
CrossRefDirect Link - Ramin, M., A.R. Alimon, J.M. Panandam, K. Sijam, A. Javanmard and N. Abdullah, 2008. Digestion of rice straw and oil palm fronds by microflora from rumen and termite bacteria, in vitro. Pak. J. Biol. Sci., 11: 583-588.
CrossRefPubMedDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Schafer, A., R. Konrad, T. Kuhnigk, P. Kampfer, H. Hertel and H. Konig, 1996. Hemicellulose-degradating bacteria and yeasts from the termite gut. J. Applied Microbiol., 80: 471-478.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Varma, A., B. Krishna Kolli, J. Paul, S. Saxena and H. Konig, 1994. Lignocellulose degradation by microorganisms from termite hills and termite guts: A survey on the present state of art. FEMS. Microbiol. Rev., 15: 9-28.
CrossRef - Wenzel, M., I. Schonig, M. Berchtold, P. Kampfer and H. Konig, 2002. Aerobic and facultatively anaerobic cellulolytic bacteria from the gut of the termite Zootermopsis angusticollis. J. Applied Microbiol., 92: 32-40.
CrossRefDirect Link - Ramin, M., A.R. Alimon, K. Sijam and N. Abdullah, 2008. Filter paper degradation by bacteria isolated from local termite gut. Res. J. Microbiol., 3: 565-568.
CrossRefDirect Link