
W. Tongbai
Department of Biology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
P. Damrongphol
Department of Biology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 3 | Page No.: 275-281







ABSTRACT
Metabolic activation of chlorpyrifos to chlorpyrifos-oxon via cytochrome-P450 (CYP450) in the riceland prawn Macrobrachium lanchesteri was examined. Direct effects of chlorpyrifos-oxon on Acetylcholinesterase (AChE) activity was studied in vitro by exposing M. lanchesteri supernatants to chlorpyrifos-oxon. And the role of CYP450 in the bioactivation was investigated in vivo by treating M. lanchesteri with piperonyl butoxide, a potent CYP450 inhibitor, prior to chlorpyrifos exposure. The study indicated that chlorpyrifos-oxon was a more potent AChE inhibitor than chlorpyrifos. However, chlorpyrifos concentrations which did not inhibit AChE activity in vitro exerted AChE inhibition in vivo. CYP450 played an important role in the bioactivation of chlorpyrifos since M. lanchesteri pretreated with 500 μg L-1 piperonyl butoxide for 24 h prior to 1.5 μg L-1 chlorpyrifos exposure for 96 h significantly attenuated the inhibition of AChE activity caused by chlorpyrifos. Nevertheless, pretreatment with piperonyl butoxide did not alter the effects of chlorpyrifos in reducing catalase activity, increasing lipid peroxidation as evidenced by increased thiobarbituric acid reactive substance levels and inducing histopathological changes such as swelling of the gills and degeneration of the hepatopancreatic cells. In conclusion, chlorpyrifos was bioactivated to chlorpyrifos-oxon in M. lanchesteri via CYP450. Chlorpyrifos-oxon was a strong AChE inhibitor but did not alter the effects of chlorpyrifos in inducing oxidative stress and histopathological changes.
PDF Abstract XML References Citation
Received: May 21, 2011;
Accepted: July 14, 2011;
Published: August 15, 2011
How to cite this article
W. Tongbai and P. Damrongphol, 2011. Bioactivation of Chlorpyrifos in the Riceland Prawn, Macrobrachium lanchesteri. Journal of Biological Sciences, 11: 275-281.
DOI: 10.3923/jbs.2011.275.281
URL: https://scialert.net/abstract/?doi=jbs.2011.275.281
DOI: 10.3923/jbs.2011.275.281
URL: https://scialert.net/abstract/?doi=jbs.2011.275.281
INTRODUCTION
Organophosphate Pesticides (OPs) have been widely used to control agricultural pests but are frequently harmful to non-target aquatic organisms due to contamination of the aquatic environment by drainage from agricultural areas (Roche et al., 2007; Joseph and Raj, 2011). Chlorpyrifos is one of the most widely used OPs; it is highly toxic to non-target organisms, particularly arthropods (Crane et al., 2003; Demirel, 2007; Deligeorgidis et al., 2008). The primary effect of chlorpyrifos is an irreversible inhibition of Acetylcholinesterase (AChE), an enzyme at the cholinergic synapses in the nervous systems of both vertebrates and invertebrates, resulting in an accumulation of neurotransmitter acetylcholine at the synapses (Taylor, 1990; Mohammad et al., 2007). Such accumulation causes hyperactivity, paralysis and finally death of the organism (Fulton and Key, 2001).
Chlorpyrifos is a relatively weak AChE inhibitor but it undergoes metabolic activation within an organism to become a more potent AChE inhibitor, chlorpyrifos-oxon, via a group of enzymes called cytochrome-P450 (CYP450) isozymes (Ankley and Collyard, 1995; Mutch and Williams, 2006). Studies in mammalian system indicate that the rate of activation is dependent on the level of CYP450 (Smith et al., 2009). The presence of piperonyl butoxide, a non-toxic and potent CYP450 inhibitor, reduced the toxic effects of chlorpyrifos on mortality, malformation and AChE activity in water flea Ceriodaphnia dubia and South African clawed frog Xenopus laevis (El-Merhibi et al., 2004). AChE also plays a role in development of neurons and network formation (Paraoanu and Layer, 2008). Chlorpyrifos was found to interfere with both cholinergic and neural developmental effects of AChE. Exposure to chlorpyrifos-oxon using rat (Yang et al., 2008) or zebrafish (Jacobson et al., 2010) embryo model disrupted neural growth. However, information on chlorpyrifos bioactivation and the role of AChE in neural development in aquatic invertebrates especially freshwater crustaceans are limited.
Chlorpyrifos induces oxidative stress in an organism by over-production of Reactive Oxygen Species (ROS), which are highly reactive thus damaging biological molecules and subsequent increased lipid peroxidation (Livingstone, 2001). Chlorpyrifos also causes alterations in activity of antioxidant enzymes such as catalase (Kavitha and Rao, 2008) and causes histopathological alterations in intoxicant organisms (Ahmed et al., 2010).
Macrobrachium lanchesteri De Man are small freshwater prawns commonly found in the vicinity of agricultural areas throughout Southeast Asia. They are subject to risks of exposure to agrochemicals including OPs and are good bioindicator for freshwater ecotoxicological assessment (Shuhaimi-Othman et al., 2006). The objectives of the present study were to investigate chlorpyrifos bioactivation in freshwater crustaceans using M. lanchesteri as a model. Direct exposure of M. lanchesteri supernatants to chlorpyrifos-oxon was carried out to study the effects of chlorpyrifos-oxon on AChE in vitro. And pretreatment of M. lanchesteri with piperonyl butoxide prior to chlorpyrifos exposure was performed to investigate the role of CYP450 in vivo. Furthermore, the effects of chlorpyrifos on catalase activity, lipid peroxidation and histopathology were also examined.
MATERIALS AND METHODS
Chemicals and supplies: Chlorpyrifos (99.5% purity) and chlorpyrifos-oxon (94.0% purity) were purchased from Dr. Ehrenstorfer GmbH (Augsburg, Germany). Piperonyl butoxide, Acetylthiocholine iodide (ATChI), 5,5-dithio-bis-2-nitrobenzoate (DTNB), 1-chloro-2,4-dinitrobenzene(CDNB) and 1,1,3,3-tetramethoxypropane were purchased from the Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
Duration of study: The studies were carried out from June 2009 to December 2010.
Prawn samples: Adult M. lanchesteri (0.3-0.5 g in weight and 3.0-4.0 cm in length) were purchased from a local market in Bangkok, Thailand. They were acclimatized under laboratory conditions in an aerated glass aquarium (50x120x50 cm) containing about 200-300 L of water at 25-27°C, 12 h : 12 h light: dark cycle for about one wk. They were starved for 24 h prior to experimentation.
Preparation of supernatant: The whole bodies of M. lanchesteri instead of separate organs were used for the study due to their small size. The exoskeleton of each prawn was removed; and the whole body was cut into small pieces and homogenized in 5 mL ice-cold homogenization buffer (100 mM sodium phosphate buffer, pH 7.2). Crude homogenate was centrifuged twice at 10,000 x g, 4°C for 20 min each. Supernatant was collected for biochemical assay immediately or stored at -70°C until use.
Effects of chlorpyrifos-oxon on AChE activity in vitro: Stock solutions of chlorpyrifos-oxon and of chlorpyrifos were prepared in absolute ethanol. Five μL of various concentrations of chlorpyrifos-oxon or chlorpyrifos solution were incubated with 495 μL of pooled prawn supernatant to give final concentrations of chlorpyrifos-oxon or chlorpyrifos ranging from 10¯9 to 10-4 M. Five μL of water or absolute ethanol were used as control and vehicle control. The mixture was incubated at 25°C for 30 min. AChE activity was determined using the method of Ellman et al. (1961). Briefly, 75 μL mixture were incubated with 875 μL reaction buffer (100 mM sodium phosphate buffer, pH 8.0 and 0.36 mM DTNB) at 25°C for 10 min. Then, 50 μL of substrate solution (10 mM ATChI in 100 mM sodium phosphate buffer, pH 8.0) were added. The absorbance was recorded every 30 sec for 3 min at 412 nm. AChE activity was expressed as nmol substrate hydrolyzed min-1 mg total protein-1. Four replicate experiments were performed.
Effects of piperonyl butoxide: Chlorpyrifos and piperonyl butoxide were dissolved in acetone. Prawns were randomly divided into five groups each of 30 prawns. Group I was the control; group II was the vehicle control exposed to 0.05 % acetone; group III was the piperonyl butoxide treated control exposed to 500 μg L-1 piperonyl butoxide for 24 h; group IV was the experimental group pretreated with 500 μg L-1 piperonyl butoxide for 24 h prior to 1.5 μg L-1 chlorpyrifos exposure for 96 h and group V was the experimental group exposed to 1.5 μg L-1 chlorpyrifos for 96 h. At the end of the experiment, prawns were collected for biochemical and histological studies. Piperonyl butoxide and chlorpyrifos concentrations were selected from preliminary studies. Three replicate experiments were performed.
Biochemical assay: AChE activity was determined as described previously. Catalase activity was determined using the method of Aebi (1984). Lipid peroxidation level was determined by measuring Thiobarbituric Acid Reactive Substance (TBARS) level according to the method of Buege and Aust (1978). The protein content of prawn supernatant was determined following the method of Bradford (1976).
Histological study: The gills and the hepatopancreases were dissected out and fixed in Bouin’s solution for 24 h before dehydrating through a graded series of ethanol solutions. They were cleared with xylene, embedded in paraffin, cut at 5 μm thickness and stained with hematoxylin-eosin. Histological alterations were observed using an Olympus BX51 microscope (Olympus Optical Co., Tokyo) and the images were captured with an Olympus DP50 digital camera.
Statistical analysis: Statistical significance of differences were set at p<0.05 calculated with one-way Analysis Of Variance (ANOVA) followed by Duncan's test using the SPSS statistical package program (SPSS, Chicago, IL).
RESULTS
Effects of chlorpyrifos-oxon on AChE activity in vitro: The presence of 10-9 M chlorpyrifos-oxon did not inhibit M. lanchesteri AChE activity in vitro; however, a concentration at 5x10-9 M significantly inhibited (p<0.05) AChE activity and a concentration at 10-8 M or higher almost completely inhibited AChE activity (Fig. 1). However, chlorpyrifos at the same concentrations did not have any effect on AChE activity; a high concentration of chlorpyrifos at 10-4 M was required to significantly inhibit (p<0.05) AChE activity (Fig. 1).
Effects of piperonyl butoxide on biochemical alteration: There were no significant differences in M. lanchesteri AChE activities among the control, the vehicle control and the piperonyl butoxide treated control. Pretreatment with 500 μg L-1 piperonyl butoxide for 24 h prior to chlorpyrifos exposure caused significantly lower (p<0.05) AChE activity than those of the controls but higher (p<0.05) activity than those exposed to chlorpyrifos without piperonyl butoxide pretreatment. Exposure to 1.5 μg L-1 chlorpyrifos for 96 h markedly inhibited AChE activity (Fig. 2a). In contrast, piperonyl butoxide pretreatment did not change the effect of chlorpyrifos on catalase activity or TBARS level.
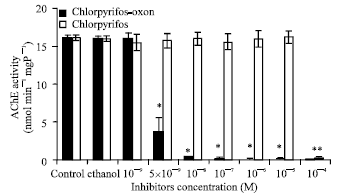 | |
| Fig. 1: | Effects of chlorpyrifos-oxon on M. lanchesteri AChE activity in vitro. Five μL of chlorpyrifos-oxon or chlorpyrifos solution were incubated with 495 μL prawn supernatant at 25°C for 30 min; the AChE activity was determined by adding 75 μL of mixture into 875 μL reaction buffer (100 mM sodium phosphate buffer, pH 8.0 and 0.36 mM DTNB) at 25°C for 10 min; then, 50 μL of substrate solution (10 mM ATChI in 100 mM sodium phosphate buffer, pH 8.0) The absorbance was recorded every 30 sec for 3 min at 412 nm. Each bar represents mean±SE from four replicate experiments. *Indicates significantly different from the control |
Exposure to 1.5 μg L-1 chlorpyrifos for 96 h with or without piperonyl butoxide pretreatment reduced catalase activity (Fig. 2b) but increased TBARS level (Fig. 2c). There were no significant differences (p>0.05) in the activities of catalase or in TBARS levels between these two groups.
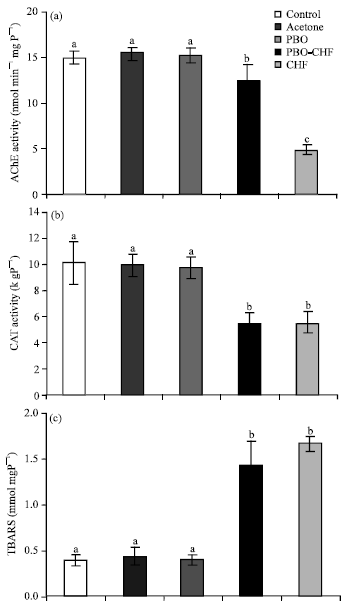 | |
| Fig. 2(a-c): | Effects of piperonyl butoxide on M. lanchesteri (a) AChE activity, (b) catalase activity and (c) TBARS level. Each experiment comprised five different groups: the control, the vehicle control, the 500 μg L-1 piperonyl butoxide treated control, the group pretreated with 500 μg L-1 piperonyl butoxide for 24 h prior to 1.5 μg L-1 chlorpyrifos exposure for 96 h and the group exposed to 1.5 μg L-1 chlorpyrifos for 96 h. Each bar represents mean±SE from three replicate experiments. Different letters indicate significant differences (p<0.05) between groups. PBO = Piperonyl butoxide; CHF = Chlorpyrifos |
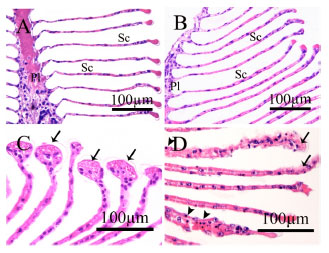 | |
| Fig. 3(a-d): | Light micrographs of M. lanchesteri gill. (A) The control showing primary lamella (Pl) and stacks of secondary lamellae (Sc). (B) The 500 μg L-1 piperonyl butoxide treated control showing similar morphology as the control. (C) The group pretreated with 500 μg L-1 piperonyl butoxide for 24 h prior to 1.5 μg L-1 chlorpyrifos exposure for 96 h showing enlargement at the distal end of secondary lamellae (arrows). (D) The group exposed to 1.5 μg L-1 chlorpyrifos for 96 h showing swollen gills, thickening of the secondary lamellae (arrows) and increased hematocyte infiltration (arrow head) |
Again, no significant differences (p>0.05) in the activities of catalase or the levels of TBARS among the control, the vehicle control and the piperonyl butoxide treated control were observed (Fig. 2b, c).
Effects of piperonyl butoxide on histological alteration: All controls did not show any histopathological changes of the tissues. The gills of the control and the piperonyl butoxide treated control consisted of a primary lamella and plate-like secondary lamellae (Fig. 3a, b). Exposure to 1.5 μg L-1 chlorpyrifos for 96 h with or without pretreatment with 500 μg L-1 piperonyl butoxide caused similar histopathological alterations which included edema of the gills, enlargement of the distal ends of the secondary lamellae forming swollen club-shaped ends (Fig. 3c), increased hematocyte infiltration and lifting off of the epithelium from the secondary lamellae (Fig. 3c, d). The hepatopancreas of the control and the piperonyl butoxide treated control contained branched, tubular structures. Cross-section of the tubules showed star shaped lumens; four different types of hepatopancreatic cells: embryonic cells, fibrillar cells, blister-like cells and resorptive cells, were observed (Fig. 4a , b).
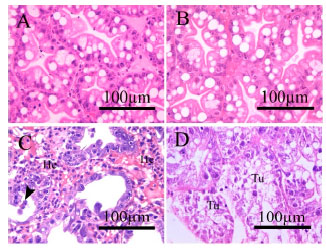 | |
| Fig. 4(a-d): | Light micrographs of M. lanchesteri hepatopancreas. (A) Cross-section of hepatopancreatic tubules of the control showing star-like structure of the lumen and large, relatively round hepatopancreatic cells (B) The 500 μg L-1 piperonyl butoxide treated control showing similar morphology as the control. (C) The group pretreated with 500 μg L-1 piperonyl butoxide for 24 h prior to 1.5 μg L-1 chlorpyrifos exposure for 96 h showing accumulation of hematocytes (He) in the hemocoel, degeneration and detachment of hepatopancreatic cells (arrow head) from the tubules. (D) The group exposed to 1.5 μg L-1 chlorpyrifos for 96 h showing degeneration of hepatopancreatic cells in the tubules (Tu) |
Exposure to 1.5 μg L-1 chlorpyrifos for 96 h with or without pretreatment with 500 μg L-1 piperonyl butoxide for 24 h induced degeneration of the tubules, hematocyte infiltration into the intertubular hemocoels and the lumens became relatively flattened loosing the star- shaped (Fig. 4c, d).
DISCUSSION
In vitro study in the present investigation indicated that chlorpyrifos-oxon was a more potent AChE inhibitor than chlorpyrifos. The concentrations which significantly inhibited AChE activity by chlorpyrifos-oxon did not alter AChE activity by chlorpyrifos. OPs are weak AChE inhibitors because the sulfur moiety of OPs has a lower electronegativity than the oxygen moiety. In an organism, OPs undergo bioactivation into OP-oxon by replacing sulfur with oxygen, which readily binds to a serine hydroxyl group in the active site of AChE leading to the inhibition of AChE activity (Fukuto, 1990). The present study indicated that chlorpyrifos was first bioactivated into chlorpyrifos-oxon before eliciting AChE inhibition in M. lanchesteri since a chlorpyrifos concentration of 1.5 μg L-1 (equivalent to 4.28 nM) did not inhibit M. lanchesteri AChE activity in vitro but exerted AChE inhibition in M. lanchesteri in vivo. Moreover, M. lanchesteri pretreated with a CYP450 inhibitor, piperonyl butoxide, reduced the degree of AChE inhibition caused by chlorpyrifos, suggesting an involvement of CYP450 in the bioactivation of chlorpyrifos into a more potent AChE inhibitor in vivo. Such findings are consistent with previous studies. Piperonyl butoxide has been reported to decrease the degree of AChE inhibition caused by malathion in Nile tilapia Oreochromis niloticus (Pathiratne and George, 1998), diazinon in Medaka Oryzias latipes (Hamm et al., 2001), chlorpyrifos in water flea C. dubia and South African clawed frog X. laevis (El-Merhibi et al., 2004) and to be responsible for insecticide resistance in cotton ballworm Helicoverpa armigera (Ramasubramanian and Regupathy, 2004). Furthermore, CYP450 isozymes, CYP2B6 and CYP3A4, have been shown to be major enzymes mediating bioactivation of chlorpyrifos into chlorpyrifos-oxon in the human liver (Mutch and Williams, 2006).
Pretreatment with piperonyl butoxide attenuated the inhibition of M. lanchesteri AChE activity by chlorpyrifos but did not alter the effects of chlorpyrifos on catalase activity, lipid peroxidation level and histopathological conditions. Chlorpyrifos-oxon did not seem to mediate these reactivities. Chlorpyrifos itself directly induced oxidative stress in M. lanchesteri. This is consistent with previous studies in the amphipods (Steevens and Benson, 1999) and the land snail Helix aspersa (Salama et al., 2005). Oxidative stress occurs by over-generation of ROS and/or disruption of antioxidant enzymes. Increased lipid peroxidation production as evidenced by an increased TBARS level is induced by oxidative decomposition of polyunsaturated fatty acids damaged by ROS (Bagchi et al., 1995). Catalase, on the other hand, is a ROS scavenging enzyme decomposing H2O2 to H2O and O2 (Escobar et al., 1996). The reduction of catalase activity in the present study caused an accumulation of H2O2 which in turn enhanced oxidative stress.
Chlorpyrifos induced histopathological alterations in the gills and the hepatopancreas of M. lanchesteri. Edema of the gills and swelling at the distal ends of the secondary lamellae are defense mechanisms in response to toxicants in aquatic animals to increase the diffusion distance between external and internal environments. The marked increase in hematocyte infiltration into the intertubular hemocoels of the hepatopancreas seemed to be an inflammatory response stimulated by damaged tissues. That more striking tissue damage was observed in the hepatopancreas than in the gills may be due to the fact that the hepatopancreas is a high lipid containing organ, thus enhancing accumulation of lipophilic pesticides including chlorpyrifos. Similar histopathological alterations have previously been reported in the giant freshwater prawn M. rosenbergii exposed to trichlorfon (Chang et al., 2006), the estuarine crab Chasmagnathus granulatus exposed to methyl parathion (Bianchini and Monserrat, 2007) and even in freshwater fish Glossogobius giuris obtained from polluted environment (Venkataraman et al., 2007). Histopathological alterations may be the result of oxidative stress. ROS are highly reactive and readily attack cellular molecules leading to loss of membrane permeability and integrity, inactivation of protein, lipid and DNA and eventually dysfunction of cellular homeostasis and cell death (Livingstone, 2001; Ahmed et al., 2010).
CONCLUSION
In conclusion, chlorpyrifos is bioactivated in M. lanchesteri into chlorpyrifos-oxon, which is more potent than chlorpyrifos in inhibiting AChE activity. Exposure to chlorpyrifos in the present study reduced catalase activity, increased lipid peroxidation production and induced histopathological alterations in M. lanchesteri.
ACKNOWLEDGMENTS
The authors would like to thank John R. Milne for critical reading of the manuscript. This work was supported by the grant from Higher Education Commission, Ministry of Education, Thailand and in part by Center for Environmental Health, Toxicology and Management of Chemicals, CHE (3328 Si Ayutthaya Road, Bangkok, Thailand).
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Ahmed, N.S., A.S. Mohamed and M.A. Abdel-Wahhab, 2010. Chlorpyrifos-induced oxidative stress and histological changes in retinas and kidney in rats: Protective role of ascorbic acid and alpha tocopherol. Pesticides Biochem. Physiol., 98: 33-38.
CrossRef - Ankley, G.T. and S.A. Collyard, 1995. Influence of piperonyl butoxide on the toxicity of organophosphate insecticides to 3 species of fresh-water benthic invertebrates. Comp. Biochem. Physiol., 110: 149-155.
Direct Link - Bianchini, A. and J.M. Monserrat, 2007. Effects of methyl parathion on Chasmagnathus granulatus hepatopancreas: Protective role of Sesamol. Ecotoxicol. Environ. Saf., 67: 100-108.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chang, C.C, P.P. Lee, J.P. Hsu, S.P. Yeh and W. Cheng, 2006. Survival and biochemical, physiological and histopathological responses of the giant freshwater prawn, Macrobrachium rosenbergii, to short-term trichlorfon exposure. Aquaculture, 253: 653-666.
CrossRefDirect Link - Crane, M., P. Whitehouse, S. Comber, C. Watts, J. Giddings, D.R.J. Moore and E. Gist, 2003. Evaluation of probabilistic risk assessment of pesticides in the UK: Chlorpyrifos use on top fruit. Pest. Manage. Sci., 59: 512-526.
PubMed - Deligeorgidis, P.N., N.P. Deligeorgidis, S. Kayoglou, G. Sidiropoulos and M. Vaiopoulou et al., 2008. Monitoring and control of Quadraspidiotus perniciosus (Comstock) Hemiptera: Diaspididae on apple trees in the prefecture of Florina, Greece. J. Entomol., 5: 381-388.
CrossRefDirect Link - Demirel, N., 2007. Mortality of false chinch bug, Nysius raphanus (Howard), to selected insecticides. J. Entomol., 4: 155-159.
CrossRefDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - El-Merhibi, A., A. Kumar and T. Smeaton, 2004. Role of piperonyl butoxide in the toxicity of chlorpyrifos to Ceriodaphnia dubia and Xenopus laevis. Ecotoxicol. Environ. Saf., 57: 201-212.
PubMed - Escobar, J.A., M.A. Rubio and E.A. Lissi, 1996. Sod and catalase inactivation by singlet oxygen and peroxyl radicals. Free Radical Biol. Med., 20: 285-290.
CrossRefDirect Link - Fukuto, T.R., 1990. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect., 87: 245-254.
PubMedDirect Link - Fulton, M.H. and P.B. Key, 2001. Acetylcholinesterase inhibition in estuarine fish and invertebrates as indicator of organophosphorous insecticide exposure and effects. Environ. Toxicol. Chem., 20: 37-45.
CrossRefDirect Link - Hamm, J.T., B.W. Wilson and D.E. Hinton, 2001. Increasing uptake and bioactivation with development positively modulate diazinon toxicity in early life stage Medaka (Oryzias latipes). Toxicol. Sci., 61: 304-313.
CrossRefPubMedDirect Link - Jacobson, S.M., D.A. Birkhlz, M.L. McNamara, S.B. Bharate and K.M. George, 2010. Subacute developmental exposure of zebrafish to the organophosphate pesticide metabolite, chloryrifos-oxon, results in defects in Rohon-Beard sensory neuron development. Aquat. Toxicol., 100: 101-111.
CrossRef - Joseph, B. and S.J. Raj, 2011. Impact of pesticide toxicity on selected biomarkers in fishes. Int. J. Zool. Res., 7: 212-222.
CrossRefDirect Link - Livingstone, D.R., 2001. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull., 42: 656-666.
CrossRefDirect Link - Mohammad, F.K., M.H.I. Al-Zubaidy and A.S. Alias, 2007. Electrometric determination of erythrocyte, plasma and whole blood cholinesterase activities in sheep, goats and cattle and their in vitro inhibition by anticholinesterase insecticides. J. Pharmacol. Toxicol., 2: 131-141.
CrossRefDirect Link - Paraoanu, L.E. and P.G. Layer, 2008. Acetylcholinesterase in cell adhesion, neurite growth and network formation. FEBS J., 275: 618-624.
CrossRefDirect Link - Pathiratne, A. and S.G. George, 1998. Toxicity of malathion to Nile tilapia, Oreochromis niloticus and modulation by other environmental contaminants. Aquatic Toxicol., 43: 261-271.
Direct Link - Ramasubramanian, T. and A. Regupathy, 2004. Magnitude and mechanism of insecticide resistance in Helicoverpa armigera Hub. population of Tamil Nadu, India. Asian J. Plant Sci., 3: 94-100.
CrossRefDirect Link - Roche, H., A. Tidou and A. Persic, 2007. Organochlorine pesticides and biomarker responses in two fishes Oreochromis niloticus (Linnaeus, 1758) and Chrysichthys nigrodigitatus (Lacepede, 1803) and an invertebrate, Macrobrachium vollenhovenii (Herklot, 1857), from the lake Taabo (Cote d`Ivoire). J. Applied Sci., 7: 3860-3869.
CrossRefDirect Link - Salama, A.K., K.A. Osman, N.A. Saber and S.A. Soliman, 2005. Oxidative stress induced by different pesticides in the Land Snails, Helix aspersa. Pak. J. Biol. Sci., 8: 92-96.
CrossRefDirect Link - Shuhaimi-Othman, M., A. Abas, S.S. Yap and M. Maziati, 2006. Bioaccumulation and elimination of copper and lead by freshwater prawn Macrobrachium lanchesteri. J. Biol. Sci., 6: 717-722.
CrossRefDirect Link - Smith, J.N., J.A. Campbell, A.L. Busby-Hjerpe, S. Lee, T.S. Poet, D.B. Barr and C. Timchalk, 2009. Comparative chlorpyrifos pharmacokinetics via multiple routes of exposure and vehicles of administration in the adult rat. Toxicology, 261: 47-58.
PubMed - Steevens, J.A. and W.H. Benson, 1999. Toxicological interactions of chlorpyrifos and methyl mercury in the amphipod, Hyalella azteca. Toxicol. Sci., 52: 168-177.
PubMedDirect Link - Venkataraman, G.V., P.N.S. Rani, N.S. Raju, S.T. Girisha and B.V. Raghavendra, 2007. Physico-chemical characteristics and impact of aquatic pollutants on the vital organs of a freshwater fish Glossogobius giuris. Res. J. Environ. Toxicol., 1: 1-15.
CrossRefDirect Link - Yang, D., A. Howard, D. Bruun, M. Ajua-Alemanj, C. Pickart and P.J. Lein, 2008. Chlorpyrifos and chlorpyrifos-oxon inhibit axonal growth by interfering with the morphogenic activity of acetylcholinesterase. Toxicol. Applied Pharmacol., 228: 32-41.
PubMed - Bagchi, D., M. Bagchi, E.A. Hassoun and S.J. Stohs, 1995. In vitro and in vivo generation of reactive oxygen species, DNA damage and lactate dehydrogenase leakage by selected pesticides. Toxicol., 104: 129-140.
PubMed