
Dan-Ping Zhang
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
Hui-Jing Zhao
CAAS-ILRI Joint Laboratory on Livestock and Forage Genetic Resources, Institute of Animal Science, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100193, China
Yu-Zhu Luo
College of Animal Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
Jian-Lin Han
CAAS-ILRI Joint Laboratory on Livestock and Forage Genetic Resources, Institute of Animal Science, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100193, China
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 3 | Page No.: 261-267







ABSTRACT
Chicken interleukin 2 (chIL-2), a primary cytokine excreted mainly by activated T cells, can significantly enhance immunity. The chIL-2 has been proven to play an important role not only in the host’s response to coccidiosis and salmonella among parasite and bacterial diseases but also in the host’s enhanced immune responses to vaccines. To examine the genetic polymorphisms and their potential impact on the structure and function of the chIL-2, we first reviewed the history of identification of chIL-2 mRNA and genomic DNA sequences and analyzed the distribution of nucleotide variation in both coding and intronic sequences that were deposited in the GenBank database. Then a short DNA fragment of 374 bp to 386 bp within chIL-2 intron 2, in which a number of discontinuous insertions/deletions (in/dels) were involved in defining major sequence variations and structures, was targeted for re-sequencing in this study. A total of 15 novel haplotypes were identified in 66 indigenous and commercial chickens sampled from six countries. Three major haplogroups were characterized by two big in/dels of eight and 11 nucleotides, of which one group seemed to have experienced a recent and rapid expansion associated with domestication. Complete chIL-2 genomic DNA sequences comprised of these haplotypes are expected to match the major structural lineages and groups within the coding and intronic sequences and to contribute to a deeper understanding of the evolution and function of chIL-2 gene.
PDF Abstract XML References Citation
Received: March 22, 2011;
Accepted: June 03, 2011;
Published: August 15, 2011
How to cite this article
Dan-Ping Zhang, Hui-Jing Zhao, Yu-Zhu Luo and Jian-Lin Han, 2011. Characterization of Haplotype Diversity Defined by Discontinuous Insertions/Deletions Within the Intron 2 of Interleukin 2 in Different Domestic Chicken Populations. Journal of Biological Sciences, 11: 261-267.
DOI: 10.3923/jbs.2011.261.267
URL: https://scialert.net/abstract/?doi=jbs.2011.261.267
DOI: 10.3923/jbs.2011.261.267
URL: https://scialert.net/abstract/?doi=jbs.2011.261.267
INTRODUCTION
Interleukin 2 (IL-2) is a primary cytokine in the form of glycoprotein secreted mainly by activated T helper (CD4+) lymphocytes. It plays a critical role in the rapid expansion, differentiation and survival of T and B lymphocytes, monocytes and natural killer cells during immune response in mammals (Smith, 1988; Kaplan et al., 1992; Lowenthal et al., 1994; Kaiser et al., 2004; Olejniczak and Kasprzak, 2008). Hilton et al. (2002) recognized similar biological activities of chicken IL-2 (chIL-2) to those of mammalian IL-2. The first complete mRNA sequence of chIL-2 was cloned from spleen cells of Obese chicken strain but reported to be homologous to both mammalian IL-2 and IL-15 (Sundick and Gill-Dixon, 1997). Kaiser and Mariani (1999) further cloned a nearly complete genomic DNA sequence of chIL-2 from erythrocytes of inbred White Leghorn line N birds using primers designed according to the coding sequence reported by Sundick and Gill-Dixon (1997) and a modified version of 5’- and 3’-rapid amplification of cDNA ends. They verified this sequence to be chIL-2 in a single copy with four exons encoding 143 amino acids. A backcross analysis was further performed to map chIL-2 as the first non-mammalian cytokine gene on chicken chromosome 4. This was validated via a genomic analysis (Kaiser et al., 2005). However, Choi et al. (1999) wrongly assigned a full length of 800 bp long cDNA sequence cloned from a CD4+ T cell hybridoma of Hy-Line variety SC birds as IL-15 based on its four highly conserved cysteine residues that form two intra-chain disulphide bonds present in mammalian IL-15 sequences (Sundick and Gill-Dixon, 1997). This sequence was later re-classified and correctly assigned back as chIL-2 by Choi and Lillehoj (2000) as well as Lillehoj et al. (2001) based on its sole transcription in activated T lymphocytes and the presence of a short 5’ region preceding the open reading frame and a short leader sequence in its mRNA which is a characteristic of IL-2 (Sundick and Gill-Dixon, 1997). This finding was further supported by the identification of chIL-2 from a clone of the cDNA library constructed from T-cell-enriched population of spleen lymphocytes and chIL-15 from a clone of the cDNA library constructed from liver tissue (Tirunagaru et al., 2000).
On the functional analysis of chIL-2, Choi and Lillehoj (2000) as well as Miyamoto et al. (2002) found that host immune responses to Eimeria acervulina and Eimeria tenella infections involved an up-regulation of chIL-2 secretion. Li et al. (2002) observed that chIL-2 production after re-infection with Eimeria tenella may be an important factor contributing to the genetic differences between SC chickens’ resistance or TK chickens’ susceptibility to coccidiosis. Berndt et al. (2007) found that chIL-2 mRNA expression between 4 h and 9 days after infection seemed to be more dependent on infection of epithelial cells in day-old chicks infected orally with four different Salmonella enterica subspecies. ChIL-2 has also been used as an adjuvant (Asif et al., 2004) to enhance the responses of vaccines to infectious bursal disease virus (Hulse and Romero, 2004; Kumar et al., 2009; Park et al., 2009), H5 subtype avian influenza virus (Hu et al., 2006) and Eimeria tenella (Xu et al., 2008). ChIL-2, therefore, is indicated to have a practical importance in enhancing immune responses to vaccines.
Through a mutational analysis, Kolodsick et al. (2001) found that, similar to mammalian IL-2 and IL-15, the aspartic acid at position 17 is a critical N-terminal contact site for binding to putative chIL-2 receptor while the removal of C-terminal amino acids leads chIL-2 protein with decreased bioactivity as a function of the kind and number of amino acids removed. Gu et al. (2010) used nine neutralizing monoclonal antibodies to the chIL-2 that were produced by recombinant chIL-2 monomer expressed in prokaryotic cells as an immunogen and to map three conformational functional domains with domain I positioned in the long A-B loop and the N terminal of Helix B, II mostly situated in Helix C and III distributed in the C-D loop and Helix D including partial Sheet B of the predicted tertiary structure of chIL-2 molecule.
The DNA sequence variation in chIL-2 has also been investigated. Kaiser and Mariani (1999) found no variation in 517 bp long promoter sequence and in all four complete exons of the eight inbred lines of White Leghorn. Zhou et al. (2001) detected a Single Nucleotide Polymorphism (SNP) of A/G at base -425 between the highly inbred (99%) Leghorn G-B1 and MHC congenic Fayoumi M15.2 lines. Following the mRNA sequence reported by Sundick and Gill-Dixon (1997), Zhou et al. (2003), Yan-Bin et al. (2006) and Huang et al. (2007) cloned mRNAs from concanavalin A stimulated spleen or blood lymphocytes of a few indigenous Chinese chicken breeds and a commercial broiler chicken line to discover sequence variation in chIL-2 cDNA. So far, a total of 19 mRNA or genomic DNA sequences have been screened from different commercial and indigenous chicken lines or breeds (Table 1). In this study, carried out from September 2009 to January 2011, we first analyzed the distribution of DNA sequence variation among these 19 publicly available sequences and then aimed to re-sequence the partial intron 2 carrying discontinuous insertions/deletions (in/dels) for a detailed haplotype characterization of chIL-2 in 66 birds of nine indigenous chicken populations collected from six countries.
MATERIALS AND METHODS
Publicly available mRNA and genomic DNA sequences retrieved from the GenBank database: After a preliminary alignment for general quality check, we retrieved 17 complete or nearly complete mRNA and two genomic DNA sequences of chIL-2 from the GenBank database, of which 12 were reported in various publications and seven directly submitted to the database. Seventeen mRNA sequences were screened from the cDNAs recovered from different lymphocytes and sequenced via cloning procedure (Table 1). Only the coding sequences (three sequences in 429 bp long and 16 sequences in 432 bp long) were included for analysis of their nucleotide variation. To assess their evolutionary relationship, a neighbor-joining phylogenetic tree was reconstructed based solely on the number of nucleotide differences between the 19 coding sequences following 1000 bootstrapping (replicates) using the MEGA software version 4.0 (Tamura et al., 2007).
Samples and DNA extraction: A total of 66 blood samples were collected from nine indigenous and commercial chicken populations in China, Cambodia, Egypt, Fiji, Lao and Uganda (Table 2). All the samples were preserved on Whatman FTA® filter papers (Whatman BioScience, Maidstone, UK) and genomic DNAs were extracted following the protocol described by Smith and Burgoyne (2004).
Selection of genomic region of interest, primer design and PCR amplification: The two genomic DNA sequences generated from the red jungle fowl and White Leghorn chicken (Table 1) were further aligned and compared to each other to identify additional variation within three introns of chIL-2. There were two SNPs within 108 bp of intron 1, 21 SNPs and six in/dels within 1740 bp of intron 2 and seven SNPs and one in/del within 413 bp of intron 3.
| Table 1: | Detailed information of 19 mRNA and genomic DNA sequences of chIL-2 |
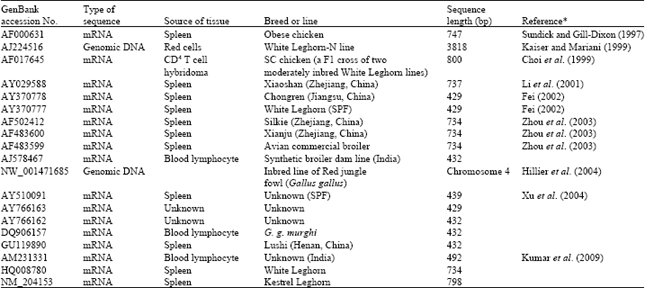 | |
| *Sequences without linked publication were directly retrieved from the GenBank database | |
| Table 2: | Sampling information of indigenous and commercial chicken populations |
 | |
We were interested in a unique structure defined by the last four discontinuous in/dels that consisted of eight, 11, one and two nucleotides within 277 bp long fragments of intron 2. This unique structure seemed to have balanced the fragments of the two sequences into the same length (Fig. 1A). Therefore, a pair of primers was designed according to these two genomic DNA sequences to generate PCR products covering these variations. Forward primer sequence was 5’-AAA GAG TTG GAA TGA GAT A-3’ located in 2092-2110 bp of AJ224516 and reverse primer 5’-TAA GCA GGT TAA TGA GAC A-3’ located in 3686-3704 bp of AJ224516. They were synthesized by the Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China).
PCR amplification reaction was performed in a 50 μL total volume containing approximate 200 ng of genomic DNA, 5 μL of 10x PCR reaction buffer (200 mM Tris-HCl, pH 8.4, 200 mM KCl, 100 mM (NH4)2SO4, 15 mM MgCl2), 0.25 mM of dNTPs, 10 pM of each primer and 2.5 unit of Taq DNA polymerase (TransGen Biotech Co., Ltd., Beijing, China).
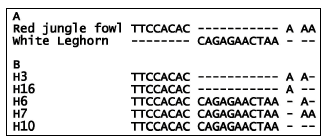 | |
| Fig. 1: | The structure of the four in/dels within intron 2 of chIL-2. (A) the four discontinuous in/dels between the red jungle fowl and White Leghorn genomic DNA sequences; (B) Five new haplotypes with different combinations of the in/dels identified in study. ‘-’ represents the deletions |
The thermocycling profile was as followed: an initial denaturation for 5 min at 94°C, followed by 35 cycles of 30 sec at 94°C, 40 sec at 60°C and 2 min at 72°C and completed by a final extension for 10 min at 72°C. A clear product of around 1600 bp was obtained.
Sequencing and haplotype identification of the PCR products: The amplicons were purified using the gel purification kit following instructions of the manufacturer (Beijing Sunbiotech Co., Ltd., Beijing, China) and then directly sequenced using the forward PCR primer and the BigDye® Terminator v3.1 Cycle Sequencing Kit on an ABI 3730 Sequence Analyzer (Applied Biosystems, Foster City, CA, USA). The raw sequencing data was manually edited using the Chromas version 1.45 (http://www.technelysium.com.au/chromas.html) and aligned with the MEGA4 software (Tamura et al. 2007). Among the 66 samples, the readable sequences of the fragment covering the four in/dels ranged from 374 bp to 386 bp in length. There were 31 homozygous and 35 heterozygous birds. From the homozygous samples, we were able to identify seven haplotypes that were subsequently used to deduce the specific genotypes and haplotypes of the heterozygous individuals. To verify the deduced haplotypes, PCR products of five representatives of different heterozygous genotypes determined by various combinations of the four in/dels were chosen for re-sequencing the fragment using a cloning procedure involving a pBM19-T vector (Biomed Co., Ltd., Beijing, China). Two to six clones per samples were selected for the re-sequencing procedure using the forward PCR primer. Finally, a total of 16 haplotypes (named as haplotypes 1 to 16) were detected within 66 samples.
Statistical analysis: The DnaSP v5 software (Librado and Rozas, 2009) was used to estimate the nucleotide diversity values for each population. Both SNPs and in/dels within the 17 haplotypes were included to construct a median-joining network using the Network 4.6.0.0 software (Bandelt et al., 1999).
RESULTS AND DISCUSSION
Distribution of nucleotide variation in chIL-2 coding sequences: There were 13 SNPs, eight of which yielded eight amino acid replacements and one in/del of AAG leading to one amino acid deletion within the 19 coding sequences. They could be classified into two major lineages defined by one transition and three transversions supported by a bootstrapping value of 98%. One lineage included five haplotypes, consisted of 13 sequences; the other had three haplotypes composed by six sequences. There was no clear pattern of haplotype distribution, e.g., the 10 mRNA sequences derived from both White Leghorn and indigenous Chinese chicken breeds fell into the two lineages (Fig. 2). This implies that there is a need to further characterize the genomic DNA sequences of chIL-2 in different chicken populations of various genetic backgrounds.
Characterization of partial chIL-2 intron 2 fragment carrying the discontinuous in/dels: Apart from the four in/dels as shown in Fig. 1A, the 16 haplotypes identified from 66 birds included in this study were further defined by 15 SNPs and two in/dels, of which seven SNPs and one in/del were present between the two partial genomic DNA sequences isolated from the red jungle fowl and White Leghorn. Eight SNPs and the other in/del were novel. One of the haplotypes (haplotype 11) was identical to the partial genomic sequence of the red jungle fowl.
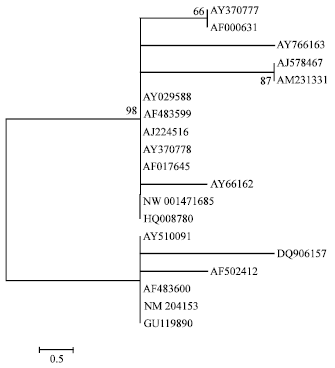 | |
| Fig. 2: | A neighbor-joining phylogenetic tree of the 19 chIL-2 coding sequences |
The remaining 15 haplotypes were novel and formed five additional combinations determined by the four in/dels within the intron 2 fragment, in particular the two big insertions of eight and 11 nucleotides were present in haplotypes 6, 7 and 10 (Fig. 1B).
The phylogenetic relationship of all 17 haplotypes including the partial genomic DNA sequence of the White Leghorn (haplotype 17) was further elucidated using a median-joining network. Haplotypes 2 and 4 were the two predominant ones with frequencies of 41 and 34, respectively, followed by 13 haplotype 12, eight each of haplotypes 1 and 11, seven haplotype 5, six each of haplotypes 3 and 7 and two each of haplotypes 6 and 10 while the remaining seven haplotypes 8, 9 and 13 through 17 were present only once.
These haplotypes were divided into three haplogroups characterized by the two big in/dels of eight and 11 nucleotides. The first group, at a frequency of 42 included haplotypes 2 and 17, was separated from the remaining 15 haplotypes by the big in/del of eight nucleotides while the second group, at a very low frequency of 10, consisted of haplotypes 6, 7 and 10. The third group had a high frequency of 82 with 12 haplotypes defined by the other big in/del of 11 nucleotides. The star like pattern dominant in the third group centered by the potential ancestral haplotype 4 and surrounded by nine derived haplotypes carrying only single nucleotide mutations indicated a recent and rapid ancestralexpansion of this group (Richards et al., 1998; Troy et al., 2001; Malhi et al., 2006; Larson et al., 2005).
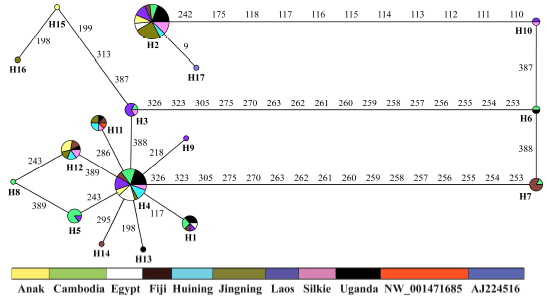 | |
| Fig. 3: | Median-joining network of 17 chIL-2 intronic haplotypes identified in 68 chicken and jungle fowl samples. Circle size is proportional to the number of samples sharing particular haplotypes and color codes indicate 11 different chicken and jungle fowl populations or samples. Lines between haplotypes represent mutational steps between haplotypes |
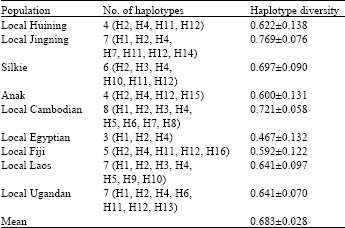
| Table 3: | Haplotype diversity of partial chIL-2 intron 2 fragment in nine chicken populations |
 | |
However, no population or geographic structuring was observed for the third group that was present in all the nine chicken populations, indicating its demic expansion from ancestral haplotypes probably associated with chicken domestication and currently shared by different chicken populations and subsequent migrations over a large geographic area (Joy et al., 2003; Mirol et al., 2008; Tofanelli et al., 2009; Zhao et al., 2011). The second group predominated only by haplotype 2 also showed no geographic structuring, pointing to its multiplication being most likely coupled with very recent, fast spreading of commercial chicken populations (Fig. 3).
Haplotype diversity of partial chIL-2 intron 2 fragments among nine chicken populations: The number of haplotypes ranged from eight in local Cambodian chickens to three in local Egyptian chickens. The highest haplotype diversity was observed in local Jingning chickens but the lowest value was present again in the local Egyptian chickens (Table 3). However, due to limited and different number of samples analyzed among the populations, it is meaningless to draw any conclusion and a further investigation is called for.
CONCLUSION
Chicken interleukin 2 plays a crucial role in the development and regulation of immune systems; therefore, it has important implications to disease control and vaccine development. We briefly reviewed the history of identification of chIL-2 mRNA and genomic DNA sequences, analyzed the distribution of nucleotide variation in both coding and intronic sequences of published data and then targeted re-sequencing a short DNA fragment within the chIL-2 intron 2 where a number of discontinuous in/dels were involved in defining major sequence variations and structures. A total of 15 novel haplotypes were identified in 66 indigenous and commercial chickens representing different genetic backgrounds, from which three major haplogroups were characterized with one of them to have followed a recent and rapid expansion associated with domestication. Further and intensive investigation of complete chIL-2 genomic DNA sequences comprising these haplotypes is expected to match the major structural lineages and groups within the coding and intronic sequences for a deeper understanding of the evolution and function of chIL-2 gene.
ACKNOWLEDGMENTS
This study was supported by the Special Funds for Junior Researchers provided by the Institute of Animal Science, CAAS (No. 2009qn-1) and the National Key Project of Scientific and Technical Supporting Programs funded by Ministry of Science and Technology of China during the 11th ‘Five-Year’ Plan (No. 2008BADB2B01).
REFERENCES
- Asif, M., K.A. Jenkins, L.S. Hilton, W.G. Kimpton, A.G. Bean and J.W. Lowenthal, 2004. Cytokines as adjuvants for avian vaccines. Immunol. Cell Biol., 82: 638-643.
CrossRefPubMedDirect Link - Bandelt, H.J., P. Forster and A. Rohl, 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37-48.
PubMedDirect Link - Berndt, A., A. Wilhelm, C. Jugert, J. Pieper, K. Sachse and U. Methner, 2007. Chicken cecum immune response to Salmonella enterica serovars of different levels of invasiveness. Infect. Immunity, 75: 5993-6007.
CrossRefDirect Link - Choi, K.D. and H.S. Lillehoj, 2000. Role of chicken IL-2 on γδ T-cells and Eimeria acervulina-induced changes in intestinal IL-2 mRNA expression and γδ T-cells. Vet. Immunol. Immunopathol., 73: 309-321.
CrossRef - Gu, J., X. Ruan, Z. Huang, J. Chen and J. Zhou, 2010. Identification of functional domains of chicken Interleukin 2. Vet. Immunol. Immunopathol., 134: 230-238.
CrossRef - Hilton, L.S., A.G. Bean, W.G. Kimpton and J.W. Lowenthal, 2002. Interleukin-2 directly induces activation and proliferation of chicken T cells in vivo. J. Interferon Cytokine Res., 22: 755-763.
Direct Link - Huang, Y.H., Z.X. Xie and J.H. Xun, 2007. Molecular cloning and sequencing analysis of interleukin-2 gene from 10 kinds of Guangxi local chickens. Anim. Husbandry Vet. Med., 39: 7-10.
Direct Link - Hillier, L.W., W, Miller, E. Birney, W. Warren1 and R.C. Hardison et al., 2004. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature, 432: 695-716.
CrossRef - Joy, A.D., X. Feng, J. Mu, T. Furuya, K. Chotivanich and A. Kretti et al., 2003. Early origin and recent expansion of Plasmodium falciparum. Science, 300: 318-321.
Direct Link - Kaiser, P. and P. Mariani, 1999. Promoter sequence, exon:intron structure and synteny of genetic location show that a chicken cytokine with T-cell proliferative activity is IL 2 and not IL 15. Immunogenetics, 49: 26-35.
CrossRef - Kaiser, P., T.Y. Poh, L. Rothwell, S. Avery and S. Balu et al., 2005. A genomic analysis of chicken cytokines and chemokines. J. Interf. Cytok. Res., 25: 467-484.
CrossRef - Kaiser, P., L. Rothwell, S. Avery and S. Balu, 2004. Evolution of the interleukins. Dev. Comp. Immunol., 28: 375-394.
PubMed - Kaplan, G., Z.A. Cohn and K.A. Smith, 1992. Rational immunotherapy with interleukin 2. Nat. Biotechnol., 10: 157-162.
CrossRef - Kolodsick, J.E., J.A. Stepaniak, W. Hu and R.S. Sundick, 2001. Mutational analysis of chicken interleukin 2. Cytokine, 13: 317-324.
CrossRef - Kumar, S., Y.S. Ahi, S.S. Salunkhe, M. Koul, A.K. Tiwari, P.K. Gupta and A. Rai, 2009. Effective protection by high efficiency bicistronic DNA vaccine against infectious bursal disease virus expressing VP2 protein and chicken IL-2. Vaccine, 27: 864-869.
CrossRef - Larson, G., K. Dobney, U. Albarella, M. Fang and E. Matisoo-Smith et al., 2005. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science, 307: 1618-1621.
CrossRef - Li, G., E.P. Lillehoj and H.S. Lillehoj, 2002. Interleukin-2 production in SC and TK chickens infected with Eimeria tenella. Avian Dis., 46: 2-9.
CrossRefDirect Link - Li, J.R., Y.W. Huang, S.S. Meng, X.Y. Liang, R.H. Xie and L. Yu, 2001. Cloning and phylogenetic analysis of interleukin-2 gene in Xiaoshan chicken, a Chinese local chicken breed. Acta Biochim. Biophy. Sin., 33: 713-718.
Direct Link - Librado, P. and J. Rozas, 2009. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25: 1451-1452.
CrossRefPubMedDirect Link - Lillehoj, H.S., W. Min, K.D. Choi, U.S. Babu and J. Burnside et al., 2001. Molecular, cellular, and functional characterization of chicken cytokines homologous to mammalian IL-15 and IL-2. Vet. Immunol. Immunopathol., 82: 229-244.
CrossRef - Malhi, R.S., G. Rhett and A.M. Bell, 2006. Mitochondrial DNA evidence of an early Holocene population expansion of threespine sticklebacks from Scotland. Mol. Phylogenet. Evol., 40: 148-154.
CrossRef - Mirol, P.M., J. Routtu, A. Hoikkala and R.K. Butlin, 2008. Signals of demographic expansion in Drosophila virilis. BMC Evol. Biol., 8: 59-59.
CrossRef - Miyamoto, T., W. Min and H.S. Lillehoj, 2002. Kinetics of interleukin-2 production in chickens infected with Eimeria tenella. Comp. Immunol. Microb. Infectious Dis., 25: 149-158.
CrossRef - Olejniczak, K. and A. Kasprzak, 2008. Biological properties of interleukin 2 and its role in pathogenesis of selected diseases-a review. Med. Sci. Monit., 14: 179-189.
PubMed - Park, J.H., H.W. Sung, B.I. Yoon and H.M. Kwon, 2009. Protection of chicken against very virulent IBDV provided by in OVO priming with DNA vaccine and boosting with killed vaccine and the adjuvant effects of plasmid-encoded chicken interleukin-2 and interferon-γ. J. Vet. Sci., 10: 131-139.
PubMed - Richards, M.B., V.A. Macaulay, H.J. Bandelt and B.C. Sykes, 1998. Phylogeography of mitochondrial DNA in Western Europe. Ann. Hum. Genet., 62: 241-260.
Direct Link - Smith, L.M. and L.A. Burgoyne, 2004. Collecting, archiving and processing DNA from wildlife samples using FTA databasing paper. BMC Ecol., 4: 4-4.
CrossRefDirect Link - Sundick, R.S. and C. Gill-Dixon, 1997. A cloned chicken lymphokine homologous to both mammalian IL-2 and IL-15. J. Immunol., 159: 720-725.
PubMedDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Tofanelli, S., G. Ferri, K. Bulayeva, L. Caciagli and V. Onofri et al., 2009. J1-M267 Y lineage marks climate-driven pre-historical human displacements. Eur. J. Hum. Genet., 17: 1520-1524.
Direct Link - Troy, C.S., D.E. MacHugh, J.F. Bailey, D.A. Magee and R.T. Loftus et al., 2001. Genetic evidence for Near-Eastern origins of European cattle. Nature, 410: 1088-1091.
CrossRefPubMedDirect Link - Yan-Bin, W., L. Zhong-Hua, K. Xiang-Tao, S. Su-Fang and S. Gui-Rong et al., 2006. Cloning and expression of gushi-chicken interleukin 2 gene and detection of its biological activity. J. Henan Agric. Univ., 40: 506-509.
Direct Link - Zhou, H., A.J. Buitenhuis, S. Weigend and S.J. Lamont, 2001. Candidate gene promoter polymorphisms and antibody response kinetics in chickens: Interferon-gamma, interleukin-2 and immunoglobulin light chain. Poult. Sci., 80: 1679-1689.
Direct Link - Zhou, J.Y., J.G. Chen and J.Y. Wang, 2003. Cloning and genetic evolution analysis of chIL-2 gene of Chinese local breeds. Prog. Biochem. Biophys., 30: 384-389.
Direct Link