
H. Al-Sheikh
Department of Biology, College of Science, King Faisal University, AlHassa, Saudia Arabia
Hani M.A. Abdelzaher
Department of Botany, Faculty of Science, Minia University 61519, El-Minia City, Egypt
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 4 | Page No.: 306-315







ABSTRACT
This study was undertaken to investigate the differences between two distinct isolates of Pythium ultimum var. ultimum, the diseased organisms of wheat and lettuce in agricultural fields in each of Egypt and Germany, respectively. Morphological study indicated that the Egyptian and German isolates comprised a single taxon, P. ultimum var. ultimum, by the current taxonomical keys. The isolates were morphologically distinguished as having smaller or larger sexual organs by the sizes of their antheridia and oogonia. The German isolate grew faster in the lower temperature range of 4-15°C whereas the Egyptian isolate grew faster in the higher temperature range of 25-37°C. Similarities in the sequence of the r-DNA-ITS including the 5.8S rDNA demonstrated evident genetic similarity at the species level between the two isolates and should be used for confirmation of identification of species of Pythium. The two isolates were not distinguishable by their pathogenicity to cucumber seedlings. The effect of temperature, hydrogen ion concentration and osmotic potential on oospores production and germination were studied. Oospores production and germination showed a similar manner, however, the Egyptian isolate produced more oospores than the German isolate. Identification of the Egyptian and German isolates of Pythium ultimum var. ultimum to the species level can be done using sequencing of r-DNA-ITS including the 5.8S rDNA, however, some morphological and physiological differences might present within the same species. This might be attributed to the effect of environmental factors and cultural conditions.
PDF Abstract XML References Citation
Received: March 14, 2010;
Accepted: April 22, 2010;
Published: June 03, 2010
How to cite this article
H. Al-Sheikh and Hani M.A. Abdelzaher, 2010. Differentiation Between Two Isolates of Pythium ultimum var. ultimum Isolated from Diseased Plants in Two Different Continents. Journal of Biological Sciences, 10: 306-315.
DOI: 10.3923/jbs.2010.306.315
URL: https://scialert.net/abstract/?doi=jbs.2010.306.315
DOI: 10.3923/jbs.2010.306.315
URL: https://scialert.net/abstract/?doi=jbs.2010.306.315
INTRODUCTION
Pythium ultimum Trow var. ultimum is one of the commonest Pythium species and can cause severe damage to many plants in cool to moderately warm climates. It is of frequent occurrence in Egypt (Abdelzaher, 1999, 2009), but intraspecific variations of P. ultimum var. ultimum are not well known. During a survey of Pythium flora in vegetable fields in Egypt and Germany, many isolates of P. ultimum var. ultimum were isolated from diseased crop plants in the two countries.
Historically, P. ultimum var. ultimum was originally isolated from rotten cress seedlings in England (Van der Plaats-Niterink, 1981). Pythium ultimum is one of the commonest Pythium species in the soil. It reproduces asexually by means of either hyphae or hyphal swellings and sexually by means of antheridia and oogonia. It has frequently been recorded from California, Texas, Hawaii, Tahiti, Canada, South America, Iceland, South Africa, Nigeria, Kenya, Kongo, Egypt, Japan, Australia, Germany, Britain, Denmark, Greece, Turkey, Slovakia, France, the Netherlands and Pakistan (Van der Plaats-Niterink, 1981; Mubeen et al., 2005; Abdelzaher, 2009). Pythium ultimum can become a severe parasite on many plants; it is a causal agent of damping-off and root-rots of many crops (Van der Plaats-Niterink, 1981; Abdelzaher, 1999, 2009).
Sexual reproduction of Pythium species takes place by means of oogonia and antheridia. After fertilization of the oogonium and then maturation of the oospore, a dormant phase is usually necessary before the germination. At germination, the oospore is converted into a thin-walled structure, which produces either a germ tube or acts as sporangium producing zoospores (Lumsden and Ayers, 1975). Therefore, oospores of Pythium species are commonly considered to be overwintering structures which act as resistant structures permitting long-time survival under unfavourable conditions (Van der Plaats-Niterink, 1981; Abdelzaher, 1999, 2009). Spreading of Pythium sp. in different soils depends upon many factors that stimulate their overwintering structures in such soils (Ayers and Lumsden, 1975). Many investigators studied the dynamics of oospores in soils rather than the biology of oospores and factors affecting their germination of the genus Pythium (Adams, 1971; Stanghellini and Burr, 1973; Ayers and Lumsden, 1975; Ge and Ichitani, 1992; Abdelzaher et al., 1994).
For this reason, studying oospore production and germination of these two Pythia isolated from Egypt and Germany may helps in understanding the dynamics of such fungi in their natural habitats.
In the present study, the characteristics of the two isolates of P. ultimum var. ultimum isolated from Egypt and Germany were compared. The isolates were compared in morphology, growth temperature, r-DNA-ITS including the 5.8S rDNA, pathogenicity and oospores production and germination under different physical and chemical factors. The isolates from Egypt and Germany are here referred to as isolate I and isolate II, respectively.
MATERIALS AND METHODS
Fungi and isolates: Mainly, two isolates of Pythium ultimum var. ultimum were selected for this study because of their economic and scientific importance. Isolate I was isolated from diseased wheat grown in an agricultural field near the bank of River Nile, Minia, Egypt (28° 6' 54" N, 45° 30' 08" E) in January, 2008. Isolate II was isolated from infected lettuce plants grown in an agricultural field in Grossbeeren, Germany, (28° 6' 54" N, 45° 30' 08" E) in July, 2009.
Fungal isolation: Isolation of P. ultimum var. ultimum (isolates I and II) from diseased wheat and lettuce plants Root rotted wheat and lettuce plants were collected from an agricultural near the bank of River Nile in Minia, Egypt (28° 6' 54" N, 45° 30' 08" E) in January, 2008 and agricultural field in Grossbeeren, Germany, (28° 6' 54" N, 45° 30' 08" E) in July, 2009. The infected wheat and lettuce plants were rinsed in ethanol (50%, v/v) for 30 sec, transferred to tap water, followed by sterile tap water, blotted with sterile filter paper and then transferred to the surface of NARM [nystatin (10 mg L-1), ampicillin (250 mg L-1), rifampicin (10 mg L-1) and miconazole (1 mg L-1) in cornmeal agar (CMA)] (Morita and Tojo, 2007; Senda et al., 2009) for isolation of Pythium species, selectively, in 9 cm Petri plates at 4 corners in each plate. Four replicates for each plant were made. The emerging hyphal tips were transferred to Water Agar (WA) for further purification from bacterial contamination as follows: distal end of a colony growing on the NARM medium was cut and re-inoculated on 2.5-3% (WA) in a Petri-dish to obtain a colony of about 1 cm diameter. The whole agar medium in the same Petri-dish and incubated until the colony reached before the edge of the dish wall. During this procedure the non-contaminated mycelia penetrated the agar medium and reached its top. Thin pieces of the agar containing a single hyphal tip of the fungus were taken from the margin of the colony and transferred to Corn Meal Agar (CMA) slants for maintaining the fungus and to CMA plates supplemented with 500 μg mL-1 wheat germ oil to check the formation of sexual structures (Abdelzaher et al., 1994).
Identification of the two isolates of P. ultimum var. ultimum: A- Morphological studies: For purification and morphological identification of isolated pythia, the NARM selective medium was found to be effective in inhibiting the development of bacteria in Pythium cultures whilst not affecting Pythium itself. The position, shape and size of sporangia, the formation of zoospores and the position, shape and size of antheridia, oogonia and oospores were determined in grass blade culture (Waterhouse, 1967). Autoclaved grass blades were placed on 2% Water Agar (WA) inoculated with each isolate. After incubation for 2 days at 25°C, colonized grass blades were transferred to autoclaved distilled water and were incubated for 1-14 days at 20°C to follow the fungal development. Morphological identification was done using the keys of Middleton (1943), Waterhouse (1967), Van der Plaats-Niterink (1981) and Dick (1990), as well as the original description of each species isolated.
Growth temperature: Growth response to temperature was investigated in the range of 1-40°C. Agar discs (7 mm in diameter) of pre-grown culture on CMA were inoculated onto Bacto-CMA (Difco) plates and incubated in darkness. Colony diameter was measured 1-2 day after inoculation and the mean growth rate (mm day-1) was calculated for each isolate. Two replicates were used for each isolate at each temperature. If no growth was observed, plates were subsequently incubated at 25°C to assess the viability of inocula.
Molecular studies: DNA extraction Mycelia were grown in V-8 agar medium (composed of 20% V8 vegetable juice v/v, 0.25% CaCO3 and were clarified by centrifugation at 13200 g for 30 min) at 25°C for 7 days or until adequate growth was observed. To extract the total genomic DNA, mycelia from the edge of Pythium colony from a culture plate were suspended in 200 μL of PrepMan Ultra Sample Preparation Reagent (Applid Biosystems, CN, USA) in a 2.0 mL microcentrifuge tube. Samples were vortexed for 10 to 30 sec and then heated for 10 min at 100°C in dry thermo unit or water bath. Samples were spin for 30 min at 15000 g. Supernatants were transferred into a new microcentrifuge tubes and were ready for (PCR) amplification by the polymerase chain reaction (Senda et al., 2009).
DNA amplification and sequencing The nuclear rDNA region of the Internal Transcribed Spacer (ITS), including the 5.8S rDNA, was amplified with the universal primers ITS4 (5′ TCCTCCGCTTATTGATATGC 3′) and ITS5 (5′ GGAAGTAAAAGTCGTAACAAGG 3′). Depending on the experiment, sometimes, primers of ITS1 (5′ TCCGTAGGTGAACCTGCGG3′) and ITS2 (5′GCTGCGTTCTTCATCGATGC 3′) were used as described by White et al. (1990) and Matsumoto et al. (1999). The amplicons were 700-900 bp long. On the other hand, 563 bp of the cox II gene was amplified in certain Pythia with the primer pair FM66 (5′ TAGGATTTCAAGATCCTGC 3′) and FM58 (5′ CCACAAATTTCACTACATTGA 3′) (Martin, 2000). Amplification of the sequencing template was carried out with DNA Thermal Cycler 2700 (Applied Biosystems) with a cycling profile of pre-PCR at 94°C for 5 min, followed by denaturation at 94°C for 1 min, 1 min primer annealing at 55°C for ITS, 52°C for cox II and elongation at 72°C, 2 min for 40 cycles, with a 7 min extension at 72°C after the final cycle. To check the presence of PCR products, 5 μL of the PCR reaction mixture was loaded in 2% L03 (Takara Bio) agarose gel, electrophoresed at 100 V, 20-30 min and stained with ethidium bromide. The sequencing templates were purified with GenElute PCR Clean-up kit (Sigma Chemical Co., St Louis, Missouri, USA) following the manufacturer’s instructions. Sequencing was performed with BigDye Terminator v3.1 Cycle Sequencing Reaction kit (Applied Biosystems) using the same primers in the initial PCR step. After purifying the sequencing reaction mixture through ethanol precipitation it was run on ABI 3100 DNA Sequencer (Applied Biosystems).
Pathogenicity: Isolates I and II were each cultured in vegetable V-8 juice broth for 3 weeks at 25°C. Suspensions of propagules were prepared from the mycelial mats of the isolates by the method of Kusunoki and Ichitani (1982) and used as the inoculum. The suspension was added to the soil previously sterilized by autoclaving and kept for 3 weeks in a black plastic bag at room temperature and the final density was adjusted to 100 and 1,000 propagules per g dry soil. The soil was passed through a 2 mm mesh, sieve and 5 mL each of soil was immediately poured into 20 cells of plastic plug flats for rice seedbeds. One each of bait seed was buried in each cell. Twenty germinated seeds of cucumber (cv. Baladi) were sown into the cell with one seed per cell. The plug flats were placed in a plastic box in a growth chamber (Precision, United States) at 25°C with 12 h photoperiod (91 μmol/m2/sec) under humid conditions. Soil water contents were maintained at approximately 30% (v/w). The number of seedlings showing damping-off was scored at 10 to 14 days after sowing. Experiments were performed in duplicate.
Oospores production: To examine the effect of temperature on oospores production, the two isolates P. ultimum var. ultimum were cultured to produce oospores at different temperatures ranging from 5 to 45°C for three weeks in 100 mL Erlenmeyer flasks containing 10 mL of clarified V-8 juice medium (composed as described previously) (Ayers and Lumsden, 1975; Abdelzaher et al., 1994). Oospore suspensions were then obtained by mincing mycelial mats in a blender for 3 min. Because average oospores diameter of P. ultimum var. ultimum was 18 μm, the resulting suspension of each isolate was filtered through a sieve, (15 μm pore diameter) in order to produce a suspension of oospores reasonably free of hyphal fragments. The number of mature (vital contents and intact walls) oospores was counted and related to the examined criteria.
The effect of pH on oospores production was conducted using 2-(N-morpholino) ethanesulfonic acid (MES) buffer (Inoue and Ichitani, 1990). Double strength of the buffer and V-8 juice medium were separately autoclaved at 121°C for 15 min and the cooled buffer was then added to an equal volume of the medium to give a final buffer concentration of 50 mM and adjusted between 4.5 and 9 at 0.5 pH unit intervals using 1 N HCl or 1 N NaOH. Flasks were incubated at 28°C for three weeks in the dark.
In the osmotic potential experiment, mannitol which was not found to be utilized by Pythium as a nutrient (Thill et al., 1979; Abdelzaher et al., 1994) was added to V-8 juice to obtain potentials of -0.13, -0.27, -0.47, -1.00, -1.65 and -3.40 MPa according to Robinson and Stokes’s formula (Robinson and Stokes, 1949). Flasks were incubated at 28°C for three weeks in the dark.
Oospores germination: The two test isolates of P. ultimum var. ultimun were cultured to produce oospores at 28°C and pH 7 for three weeks in 100 mL Erlenmeyer flasks containing 10 mL of vegetable V-8 juice. Oospore suspensions were then obtained by mincing mycelial mats in a blender for 3 min. The resulting suspension was filtered through a sieve, the size of which was chosen in relation to the oospore diameter in order to produce a suspension of oospores reasonably free of hyphal fragments (as previously described). This suspension was used directly after counting and adjusting the number of mature oospores and incubated on Difco-CMA (Difco cornmeal agar 17 g and distilled water to 1 L) medium for 24 h at fixed temperature in the dark. Oospores germination was examined microscopically and 200 oospores were selected at random for calculation of germination rate. All the results are the mean of five replicates tests, each repeated twice.
Difco-CMA medium was employed for the investigation of temperature response of oospore germination. V-8 juice medium at pH 7 was used for testing the effect of temperature on oospore germination.
In the pH experiment, CMA medium was adjusted to pH values between 4.5 and 9 at 0.5 pH unit intervals using 50 mM MES buffer (Inoue and Ichitani, 1990; Abdelzaher et al., 1994). Double strength of the buffer salt was dissolved in a fixed volume of distilled water and adjusted to the desired pH using 1 N HCl or 1 N NaOH. Double strengths of MES buffer and CMA medium were separately autoclaved at 121°C for 15 min and the buffer was then added to an equal volume of the medium to give a final buffer concentration of 50 mM. Oospores germination tests were carried out at 25°C for the studied fungi.
For studying the osmotic potential, mannitol, was added in a ratio of 0.04, 0.08, 0.16, 0.32, 0.64 and 1.28 mol kg-1 to CMA medium to create the following potentials: -0.13, -0.27, -0.47, -1.00, -1.65 and -3.40 MPa, respectively, according to Robinson and Stokes’s formula (Robinson and Stokes, 1949). The osmotic potential was determined with a Dew point Microvoltmeter (HR-33T; Wescor, Logan, USA). Incubation temperature of the test was 25°C for the tested species.
RESULTS
Ten isolates of the Egyptian Pythium were isolated from diseased wheat plants grown in an agricultural field near the bank of River Nile, Minia, Egypt (28° 6' 54" N, 45° 30' 08" E) in January, 2008 and 9 isolates of the German Pythium were also isolated from infected lettuce plants grown in an agricultural field in Grossbeeren, Germany, (28° 6' 54" N, 45° 30' 08" E) in July, 2009. The 19 isolated fungi of Pythium, which could be identified on morphological a nd molecular basis and these are; P. ultimum var. ultimum.
These were characterized as follows:
Morphology: Morphology and dimensions of sexual organs and hyphal swellings in Isolate I and Isolate II of P. ultimum var. ultimum and the related data of P. ultimum var. ultimum from the key of Van der Plaats-Niterink (1981) are presented in Table 1. I and II isolates were distinguishable from each other on the basis of morphology of sexual structures (Fig. 1a-h, 2a-i). Antheridia of the German (isolate II) tend to be shorter and thiner than those of the Egyptian (isolate I). Monoclinous antheridia were less frequently observed in the isolate I than in the Isolate II. The number of antheridia per oogonium was usually one, sometimes two in the two isolates. Oogonia and oospores of the isolate I averaged 21.5±1.7 and 18±1.2 μm, respectively, whereas those of the isolate II were 20.5±1.3 and 17.5±1.1 μm, respectively. Oogonia were mostly terminal in the two isolates. The thickness of oospore wall ranged 0.7-1.4 μm for isolate I and 0.7-0.9 μm for isolate II. The average aplerotic index was 69% for the isolate I and 61.1% for the isolate II. The average wall index was 28.6% for the isolate I and 27% for the isolate II. Hyphal swellings were mostly intercalary in all isolates and usually smaller in the isolate II than in the isolate I (Fig. 1, 2). The isolate II tended to produce spherical hyphal swellings (1.1, length/breadth) as compared to the isolate I (1.4, length/breadth). The two isolates were lacking in zoospore production at 5 and 20°C. These morphological characters of isolate I and isolate II were in accordance with the reference data of P. ultimurn var. ultimum (Van der Plaats-Niterink, 1981; Dick, 1990), except for oospore wall thickness and the wall index and the aplerotic index.
| Table 1: | Morphology and dimensions of sexual structures and hyphal swellings in the Egyptian (isolate I) and the German (isolate II) of Pythium ultimum var. ultimum |
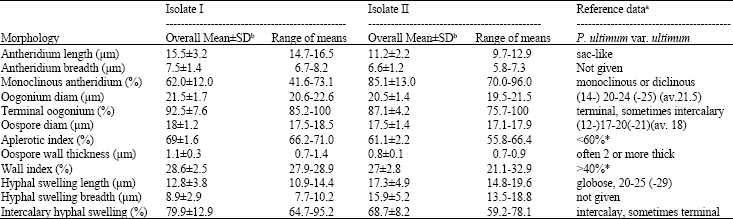 | |
| Data from Van der Plaats-Niterink (1981) (not marked) and Dick (1990) (marked with *). Six fungi of each isolate were examined with at least 30 structures | |
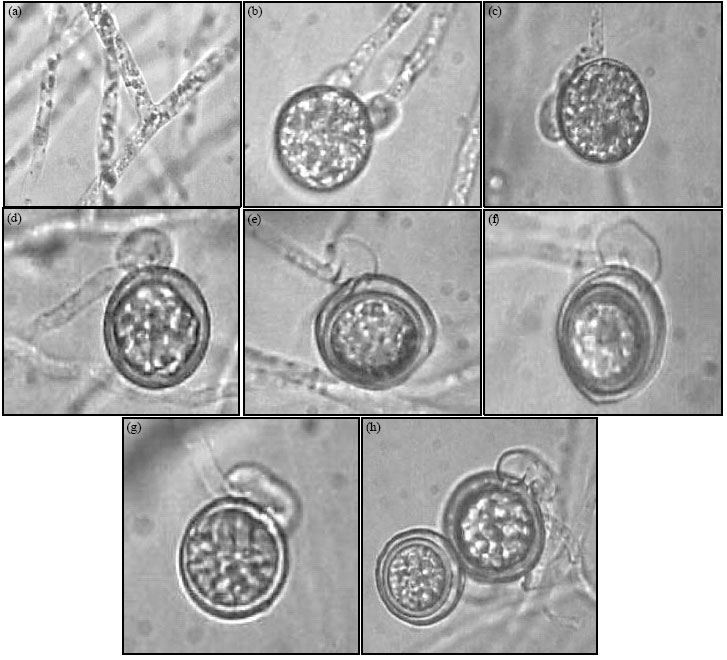 | |
| Fig. 1: | Morphology of Pythium ultimum var. ultimum (isolate I). (a) Mycelia. (b, c) Young oogonia encircled with an antheridia[3] and with 2 antheridia [2]. (d-h) Sac antheridia attached with oogonia. (f) An aplerotic oospore. (h) A thick walled aplerotic oospore. Bar (20 μm) on photo 7 is applicable to the rest photos |
Growth temperature: The isolate II was distinguishable from the isolate I by its faster growth at 1-15°C and slower growth at 25-37°C (Fig. 3). The optimum and maximum temperature for growth of all isolates were 28-30°C and 37°C, respectively. The minimum growth temperature was below 1°C for the isolate II, whereas it ranged 1-5°C for the isolate I. Growth of hypha was observed in the two isolates when colonies were transferred to the optimum temperature following incubation at 40°C. No variations in the data were found among fungi of the same isolate.
Molecular identification: Sequencing of rDNA-ITS including the 5.8SrDNA were analyzed for the two isolates of P. ultimum var. ultimum tested by the method of Kageyama et al. (2003, 2005) to confirm the species identification. The sequence of (EgH25) was closely related with that of P. ultmium var. ultmium (Genbank accession number, AY598657.1) with 100% similarity (Fig. 4).
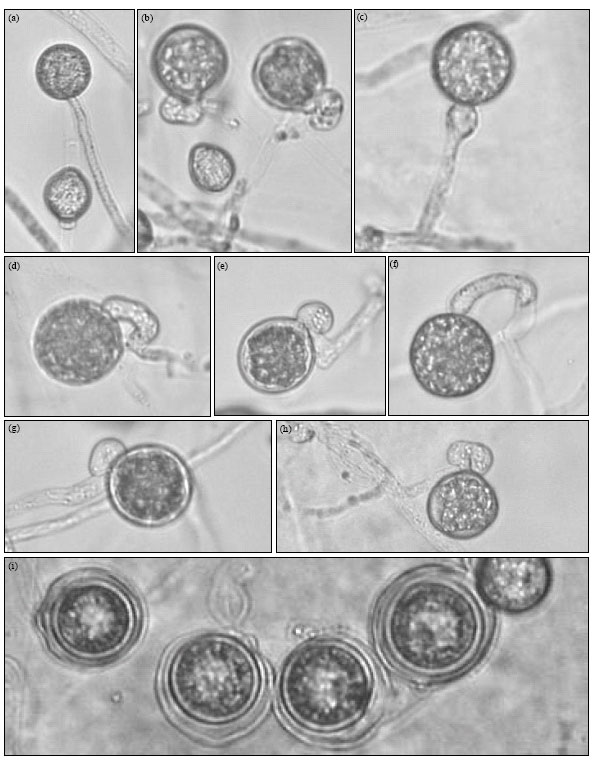 | |
| Fig. 2: | Morphology of Pythium ultimum var. ultimum (isolate II). (a) Mycelia. (1) Terminal and intercalary hyphal swellings, (b-h) Young oogonia, (c) Hypogenous antheridium (d-f) Monoclinous antheridium attached to a young oogonium, (g) Diclinous antheridium, (i) Thick walled aplerotic oospores. Bar (20 μm) on photo 3 is applicable to the rest photos |
Pathogenicity: The pathgenicity of isolates of the two isolates of P. ultimum var. ultimum on cucumber seedling was observed. Percentages of damping-off caused by the isolates were 75% damping-off with 1,000 propagules per g dry soil at 25°C. The results of the pathogenicity test were similar for the two studied isolates.
Factors affecting oospores production and germination Influence of temperature on oospores production: As shown in Fig. 5, oospores produced over a temperature ranging of 5-35°C for the 2 isolates of P. ultimum var. ultimum. The optimum range was between 25-28°C for oospores production of the two studied isolates. The Egyptian isolate I of P. ultimum var. ultimum produced oospores more than that of the German isolate II.
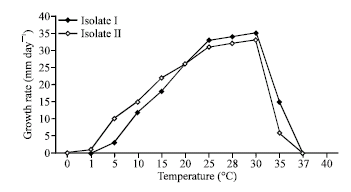 | |
| Fig. 3: | Growth-temperature relations of the Egyptian isolate I and the German isolate II of Pythium ultimum var. ultimum. Twelve fungi comprising isolate I and isolate II were used with duplicates |
 | |
| Fig. 4: | Sequence of the rDNA internal transcribed spacer (ITS) region of Pythium ultimum var. ultimum (isolate I and isolate II). The sequences were completely identical for the two isolates. The amplified DNA consists of 5.8S rDNA, studied by the method of Kageyama et al. (2003) to confirm the species identification. The sequence of EgH015 and EgH010 showed 100% similarity to (AY598657.1) |
Influence of hydrogen ion concentration on oospores production: As shown in Fig. 6, oospores production happened over a range of pH 5-9. Optimum pH values were noticed between 6.5-7 for test isolates. The Egyptian isolate I of P. ultimum var. ultimum produced oospores more than that of the German isolate II.
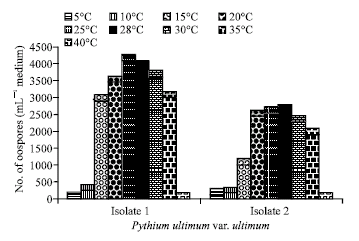 | |
| Fig. 5: | Effect of temperature on oospores production of the E1gyption (Isolate I) and German (Isolate II) fungi of Pythium ultimum var. ultimum grown on V-8 juice medium for 21 days in the dark |
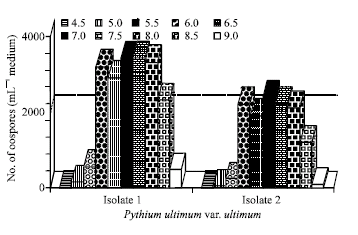 | |
| Fig. 6: | Effect of hydrogen ion concentration on oospores productio n of the Egyption (Isolate I) and German (Isolate II) of Pythium ultimum var. ultimum grown on V-8 juice medium at 28°C for 21 days in the dark |
Influence of osmotic potential on oospores production: The effect of osmotic potential on oospores production at 28°C is indicated in Fig. 7. The two tested isolates showed similar responses. Oospores produced at -0.13 to -1.65 MPa with the optimum production rate at -0.27 to -0.47 MPa. The Egyptian isolate I of P. ultimum var. ultimum produced oospores more than that of the German isolate II.
Oospores germination
Influence of temperature on oospores germination: As shown in Fig. 8, oospores of the Egyptian isolate (isolate I) germinated between 15°C and 30°C, those of the German isolate (isolate II) germinated between 10°C and 30°C, with optimum between 25-30°C for isolate I and 20-25°C for isolate II.
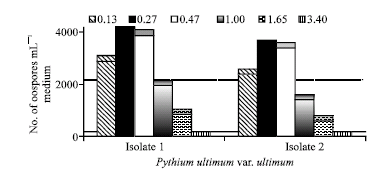 | |
| Fig. 7: | Effect of osmotic potential on oospores production of the Egyption isolate I and German isolate II of Pythium ultimum var. ultimum grown in V-8 juice medium at 28°C for 21 days in the dark |
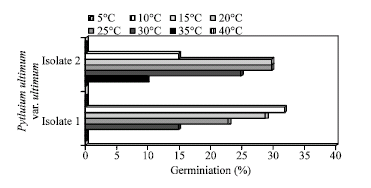 | |
| Fig. 8: | Effect of temperature on oospores germination of the Egyption isolate I and German isolate II of Pythium ultimum var. ultimum on CMA medium after 24 h in the dark |
 | |
| Fig. 9: | Effect of hydrogen ion concentration on oospores germination of the Egyption isolate I and German isolate II of Pythium ultimum var. ultimum on CMA after 24 h at 25°C in the dark |
Influence of hydrogen ion concentration on oospores germination: As presented in Fig. 9, the two isolates tested germinated over pH 5 to pH 9 with an optimum between 6.5~8.
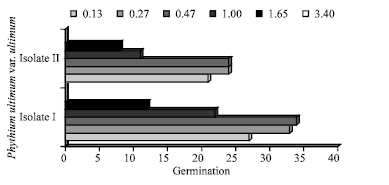 | |
| Fig. 10: | Effect of osmotic potential on oospores germination of the Egyption isolate I and German isolate II of Pythium ultimum var. ultimum on CMA after 24 h at 25°C in the dark |
Influence of osmotic potential on oospores germination: The effect of osmotic potential on oospore germination at 28°C is given in Fig. 10. The two isolates tested showed similar responses. At -3.40 MPa, non could germinate. They germinated at the other osmotic potentials tested, with good germination between -0.13 and -0.27 MPa and the optimum at -0.47 MPa.
DISCUSSION
Morphological comparisons of the reproductive structures, especially their dimensions, revealed clear differences between the Egyptian isolate I and the German isolate II of P. ultimurn var. ultimum. The isolate I was readily distinguished from the is olate II by the combination of larger oogonia and elongated antheridia. The two isolates were also distinguishable by growth temperature and oospores production. Similarities in the sequence of the r-DNA-ITS including the 5.8S rDNA demonstrated evident genetic similarity at the species level between the two isolates. Tojo et al. (1998) compared between two morphological groups of Pythium ultimum var. ultimum strains isolated in a vegetable field in Japan. They showed differences in morphology within their groups of P. ultimum var. ultimum. They further postulated that Random Amplified Polymorphic DNA (RAPD) and isozyme analyses revealed genetic dissimilarity between the two groups of P. ultimum var. ultimum. Present results were comparable with the results of Tojo et al. (1998) except that they used random amplified polymorphic DNA (RAPD) and isozyme analyses for caparisons between the two different groups of Pythium ultimum var. ultimum whereas in our study we used sequencing of the r-DNA-ITS including the 5.8S rDNA for comparison between the Egyptian and German isolates of the same species. Present results indicate that the two isolates were not distinguishable by their pathogenicity to cucumber seedling which is in agreement with the results indicated by Tojo et al. (1998). Sequencing of the r-DNA-ITS including the 5.8S rDNA is more reliable than random amplified polymorphic DNA (RAPD) and isozyme analyses because the first deals with a part of a gene and the second of isozyme analysis affected by environmental conditions (Lévesque and de Cock, 2004). Therefore, identification of Pythium species to species level can be done using in the sequence of the r-DNA-ITS including the 5.8S rDNA, despite of dissimilarities in some morphological and physiological characteristics.
Several morphological characters of isolates I and II differed from the reference data of P. ultimum var. ultimum (Van der Plaats-Niterink, 1981; Dick, 1990). Oospore wall thickness and the wall index of isolates I and II were thinner than those of the reference data. This morphological difference is difficult to explain, but both isolates can be distinguished from other thin oospore wall species such as P. irregulare and by other morphological features such as types and number of antheridia, oogonial wall and shapes of oogonia, plerotic and aplerotic characters of oospores and other features. The aplerotic index of the isolate I ranged 66.2-71% and ranged 55.8-66.4% for isolate II. Dick (1990) arbitrarily classified species with a mean aplerotic index over 60-65% as plerotic. Based on this criterion, oospores of the isolates I and II were plerotic. However, in this study, the oospores did not fill the oogonium and, therefore, they were aplerotic according to classical concepts. This concurs with the findings of Barr et al. (1996), who reported that the aplerotic index of P. ultimum oospores ranged 60.5-75.3% when the oospores did not fill the oogonium.
Oospores are considered important survival structures. They can remain viable after 8 months to 12 years in the soil (Lumsden and Ayers, 1975; Abdelzaher et al., 1994). When the soil conditions become favorable, oospores germinate to produce huge amount of disease inducing elements, which spread from place to place causing infestation. Accordingly, studying oospore production and germination of two isolates of Pythium ultimum var. ultimum isolated from diseased plants grown in Egypt and Germany is of importance in order to shed light on their life cycles under variable conditions.
The production of oospores is dependent on several factors including temperature, hydrogen ion concentration and osmotic potential. Our results here indicate that the most favorable temperatures for oospore production were 25-30°C by the tested fungi. It could be postulated that the very cold winter and very hot summer retard oospore production of such fungi (Abdelzaher et al., 1994). Oospore produced most abundantly around neutral. Previous studies revealed that Pythium sp. non-tolerant to acid soils (Webster, 1980; Abdelzaher et al., 1994). In this study, no oospores have been produced below pH 5 for the two studied isolates of P. ultimum var. ultimum. Pythium species have been absent from heavily polluted places and seldom found in partially polluted locations (Harvey, 1952; Abdelzaher et al., 1994). It would be suggested that, high polluted locations with osmotic potentials above -1.65 MPa represent inhibitive media toward oospore production. Oospores germination of these two pythia occurred over a broad pH range with an optimum around 7. Acidic soil appeared unsuitable for oospore germination. Heavy polluted places retard oospore germination, therefore, it could be expected that high polluted localities with osmotic potentials above -1.65 MPa inhibit oospore germination. Our results are comparable with the results of many previous researches (Abdelzaher et al., 1994; Cliquet and Tirilly, 2002; Ge and Ichitani, 1992).
However, the Egyptian isolate produced more oospores than the German isolate, similarities of oospore behavior (production and germination) of the two tested isolates under the studied factors were confirmed.
Finally, it is worth to mention that identification of Pythium sp. to the species level can be done using sequencing of r-DNA-ITS including the 5.8S rDNA, however, some morphological and physiological differences might present within the same species. This might be attributed to the effect of environmental factors and cultural conditions.
REFERENCES
- Abdelzaher, H.M.A., T. Ichitani and M.A. Elnaghy, 1994. Effect of temperature, hydrogen ion concentration and osmotic potential on oospore germination of five Pythium spp. isolated from pond water. Mycoscience, 35: 315-318.
CrossRefDirect Link - Ayers, W.A. and R.D. Lumsden, 1975. Factors affecting production of oospores of three Pythium [aphanidermatum, Pythium myriotylum, Pythium ultimum] species [fungi]. Phytopathology, 65: 1094-1100.
Direct Link - Cliquet, S. and Y. Tirilly, 2002. Development of a defined medium for Pythium oligandrum oospore production. Biocontrol Sci. Technol., 12: 455-467.
CrossRefDirect Link - Ge, L.P. and T. Ichitani, 1992. Influence of temperature, hydrogen ion concentration and osmotic potential on oospore germination of Pythium paddicum and P. Iwayamai. Bull. Univ. Osaka Prefecture Ser. B, 44: 7-11.
Direct Link - Harvey, J.V., 1952. Relationship of aquatic fungi to water pollution. Sewage Ind. Wastes, 24: 1159-1164.
Direct Link - Inoue, M. and T. Ichitani, 1990. Optimum pH range of mycelial growth of Pythium spp. estimated by MES [2-(N-morpholino) ethanesulfonic acid] buffer. Trans. Mycol. Soc. Jap., 31: 191-196.
Direct Link - Kageyama, K., M. Suzuki, A. Priyatmojo, Y. Oto, H. Suga, T. Aoyagi and H. Fukui, 2003. Characterization and identification of asexual strains of Pythium associated with root rot of rose in Japan. J. Phytopathol., 151: 485-491.
CrossRefDirect Link - Kageyama, K., A. Nakashima, Y. Kajihara, H. Suga and E.B. Nelson, 2005. Phylogenetic and morphological analyses of Pythium graminicola and related species. J. Gen. Plant Pathol., 71: 174-182.
CrossRefDirect Link - Kusunoki, T. and T. Ichitani, 1982. Preparation of mycelium free oospores of Pythium butleri by a freezing method. Ann. Phytopathol. Soc. Jap., 48: 695-698.
Direct Link - Levesque, C.A. and W.A. de Cock, 2004. Molecular phylogeny and taxonomy of the genus Pythium. Mycol. Res., 108: 1363-1383.
PubMedDirect Link - Martin, F.N., 2000. Phylogenetic relationships among some Pythium species inferred from sequence analysis of the mitochondrially encoded cytochrome oxidase II gene. Mycologia, 92: 711-727.
Direct Link - Matsumoto, C., K. Kageyama, H. Suga and M. Hyakumachi, 1999. Phylogenetic relationships of Pythium species based on ITS and 5.8S sequences of the ribosomal DNA. Mycoscience, 40: 321-331.
CrossRefDirect Link - Middleton, J.T., 1943. The taxonomy, host range and geographic distribution of the genus Pythium. Memories Torrey Bot. Club, 20: 1-171.
Direct Link - Morita, Y. and M. Tojo, 2007. Modifications of PARP medium using fluazinam, miconazole and nystatin for detection of Pythium spp. in soil. Plant Dis., 91: 1591-1599.
CrossRefDirect Link - Mubeen, L.A., A. Qayoom, S. Shahzad and A. Ghaffar, 2005. Pythium ultimum var. ultimum, a new record from Pakistan. Pak. J. Bot., 37: 779-782.
Direct Link - Senda, M., K. Kageyama, H. Suga and C.A. Levesque, 2009. Two new species of pythium, P. senticosum and P. takayamanum, isolated from cool-temperate forest soil in Japan. Mycologia, 101: 439-448.
PubMed - Stanghellini, M.E. and T.J. Burr, 1973. Effect of soil water potential on disease incidence and oospore germination of Pythium aphanidermatum. Phytopathology, 63: 1496-1498.
CrossRefDirect Link - Thill, D.C., R.D. Schimman and A.P. Appeby, 1979. Osmotic stability of mannitol and polyethylene glycol 20000 solutions used as seed germination media. Agron. J., 71: 105-108.
Direct Link - Tojo, M., E. Nakazono, S. Tsushima, T. Morikawa and N. Matsumoto, 1998. Characterization of two morphological groups of isolates of Pythium ultimum var. Ultimum in a vegetable field. Mycoscience, 39: 135-144.
CrossRefDirect Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link