
T. Tanee
Faculty of Environment and Resource Studies, Mahasarakham University, Mahasarakham 44000, Thailand
A. Chaveerach
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
R. Sudmoon
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
A. Tanomtong
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
P. Mokkamul
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 1 | Page No.: 51-56







ABSTRACT
Genetic relationships of cercopithecid species in Thailand are determined based on sequence analysis of the internal transcribed spacer (ITS) regions of ribosomal DNA (rDNA). Aligned sequences of the complete ITS regions obtained from the six taxa and an outgroup, Pan paniscus resulted in 634 characters. Within the Cercopithecidae, ITS sequences are highly conserved with 97.48% homology. The resulting comparisons of ITS sequences were used for analysis of genetic distance and a dendrogram was constructed based on the Neighbor Joining Method. From the dendrogram, Pan paniscus is solely separated. The Colobinae species, Trachypithecus cristatus, T. phayrei, T. obscurus, were placed inside the Cercopithecinae species, Macca arctoides, M. mulatta, M. nemestrina. The dendrogram of these species does not completely match the former classification based on morphological characters. Genetic distances are very low (0.002 to 0.013) among Thai cercopithecid species. The results suggest that the ITS regions of rDNA is highly conserved in Thai cercopithecids.
PDF Abstract XML References Citation
How to cite this article
T. Tanee, A. Chaveerach, R. Sudmoon, A. Tanomtong and P. Mokkamul, 2009. Genetic Relationships of Cercopithecidae in Thailand as Inferred from rDNA ITS Regions. Journal of Biological Sciences, 9: 51-56.
DOI: 10.3923/jbs.2009.51.56
URL: https://scialert.net/abstract/?doi=jbs.2009.51.56
DOI: 10.3923/jbs.2009.51.56
URL: https://scialert.net/abstract/?doi=jbs.2009.51.56
INTRODUCTION
The old world monkeys are a group of primates in the family Cercopithecidae. The family contains two extant subfamilies, Cercopithecinae (cercopithecines; the cheek-pouch monkeys) and Colobinae (colobines; the leaf monkeys). The cercopithecines are omnivorous, having cheek pouches and simple stomachs while the colobines are folivorous, lacking cheek pouches but having complex stomachs (Swindler, 1998).
The Colobinae comprises seven genera. Of these, five have a solely Asian distribution and two have a solely African distribution (Page et al., 1999). The phylogenetic separation of the African colobines has been clearly studied, whereas, there is a lack of information about the Asian group. In Thailand, two genera and four species are recognized, Presbytis femoralis, Trachypithecus cristatus, T. phayrei and T. obscurus (Lekagul and McNeely, 1988; Francis, 2001) and the diploid chromosome number is 44 in the extant genera (Chaveerach et al., 2007).
The Cercopithecinae comprise 11 extant genera. Genus Macaca (macaques) is the most widely distributed. Fifteen species have been found in Asia and one more species is found in Africa. The other ten genera are solely African (Page et al., 1999). In Thailand, there are five species, Macaca arctoides, M. assamensis, M. fascicularis, M. mulatta and M. nemestrina (Lekagul and McNeely, 1988; Francis, 2001). They have diploid chromosome number of 42 (Tanee et al., 2006).
However, in recent years, the genetic relationships among different cercopithecid species are still controversially discussed because of the limited material. The currently accepted classification of cercopithecids is mainly based on morphological studies, while their cytogenetics and molecular genetics have become important tools (Page et al., 1999; Tanee et al., 2006, 2007; Chaveerach et al., 2007). However, few analysis have included information from internal transcribed spacer (ITS) sequence data. As part of a study on the complete genetic relationships of Cercopithecidae, the old world monkeys in Thailand play a key role, because of the high number of endemic species and the low amount of currently available data. Moreover, the genetic relationships within wildlife cercopithecids in Thailand have never been addressed using the sequences of ITS regions.
The internal transcribed spacer (ITS) of nuclear ribosomal DNA (rDNA) is one of the most extensively sequenced molecular markers (Alvarez and Wendel, 2003). The two internal transcribed spacers, ITS1 and ITS2, of rDNA gene normally exhibit high rates of variability and have been widely used for phylogenetic studies of both closely and distantly related species (Insua et al., 2003; Sumida et al., 2004; Chaveerach et al., 2008).
From the advantage of ITS analysis above, the genetic relationships of six old world monkeys in Thailand, Macaca arctoides, M. mulatta, M. nemestrina, Trachypithecus cristatus, T. obscurus, T. phayrei, were investigated using the sequences of ITS regions. The results will be added into scientific data and will be useful for long-term management of these primates.
MATERIALS AND METHODS
Sample collection: Since December 2006, blood samples from three macaque species (Macaca arctoides, M. mulatta, M. nemestrina from Northeastern Thailand) and three leaf monkey species (Trachypithecus cristatus and T. phayrei from Northeastern Thailand, T. obscurus from Southern Thailand) were collected. The collected locations are shown on a map of Thailand (Fig. 1).
DNA isolation: The genomic DNA was extracted from blood samples using the Genomic DNA extraction kit (RBC Bioscience, USA). The quality and quantity of extracted DNA was assessed by 0.8% agarose gel electrophoresis.
 | |
| Fig. 1: | Map of Thailand indicating sample collection of cercopithecids (•). N: Northern, NE: Northeastern, C: Central, W: Western, E: Eastern, S: Southern |
Amplification of ITS regions: Internal transcribed spacers (ITS1, ITS2) and 5.8s rDNA regions were amplified using primers IT1 (5’-TCGTAACAAGGTTTCCGTAGGT-3’) and IT2 (5’-GTAAGTTTCTTCTCCTCCGCT-3’) described by Tsai et al. (2004). PCR amplifications were performed in a total volume of 25 μL, containing PCR master mix (Promega), 0.5μM each primer and 10 ng genomic DNA. Amplification reactions were performed in a thermal cycler (Gene Amp PCR System 9700) as follows: (1) 5 min initial denaturation step at 94°C; (2) 40 cycles of denaturation at 94°C for 45 sec, annealing at 55°C for 45 sec, extension at 72°C for 1 min; (3) a final extension for 10 min at 72°C. The size of PCR products was determined by agarose gel electrophoresis (1.0% w/v in TAE) with ethidium bromide staining.
DNA sequencing: The PCR products with expected size were examined for their nucleotide sequences with either IT1 or IT2 primer. The sequencing was done by using BigDye Version 3.1 (Applied Biosystems, USA) with MyGenie96ThermalBlock (Bioneer, Korea) follow by 3100 Genetic Analyzer with BigDye terminator v.3.1 cycle sequencing kit (Applied Biosystems).
Data analysis: Genetic relationships of the cercopithecid samples based on ITS sequences were analyzed. ITS sequence of pygmy chimpanzee (Pan paniscus), used as outgroup was taken from GenBank (accession number EF647787). The genetic distance matrix was calculated by the two-parameter method of Kimura and then was used to construct a phylogenetic tree using the Neighbor-Joining Method (NJ) with interior branch tests of 1000 replicates. All these analysis were done using MEGA software version 4.0 (Tamura et al., 2007).
RESULTS
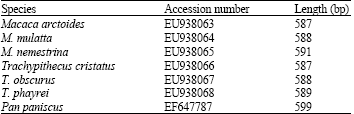
Genomic DNAs of six cercopithecid species, namely Macaca arctoides, M. mulatta, M. nemestrina, Trachypithecus cristatus, T. obscurus and T. phayrei, were amplified for their ITS regions with IT1-IT2 primers. The amplification yielded single fragments of approximately 750 bp on the gel for all samples. The fragments were analyzed for their sequences. From the sequencing results, the length of the ITS regions of six cercopithecids varied from 587 to 591 bp. All of these sequences were submitted to GenBank. The accession numbers were shown in Table 1. All of these sequences were aligned and resulted in 634 characters (Fig. 2). Among six of the Thai cercopithecid species, the number of different sites in the ITS regions is 16 (2.52%) sites and number of common sites is 618 (97.48%) sites.
A dendrogram obtained from comparisons of ITS sequences using Neighbor-Joining showed that the six cercopithecid species are clustered together, whereas Pan paniscus is separated as an outgroup. Within the cercopithecid cluster, T. obscurus is more closely related to T. phayrei than other cercopithecids (Fig. 3). The genetic distances computed from sequence alignment among all taxa are shown in Table 2. The genetic distance values vary from 0.002 (between M. nemestrina and T. cristatus) to 0.013 (between M. arctoides and T. phayrei) among cercopithecid species.
DISCUSSION
ITS sequence comparisons are becoming an increasingly popular tool for genetic relationship studies (Schlotterer et al., 1994; Insua et al., 2003; Sumida et al., 2004; Tsai et al., 2004; Chaveerach et al., 2008). In this study, the genetic relationships among cercopithecid species in Thailand have been determined by using the ITS regions including ITS1, 5.8s ribosomal DNA and the ITS2. From the alignment of ITS sequences (Fig. 2), the ITS regions are highly conserved with 97.48% homology. The results suggest that the ITS regions in family Cercopithecidae are very highly conserved sequences reflecting the close genetic relationships among cercopithecid species, whereas this region in other animals shows great variability such as in mollusks (Insua et al., 2003), amphibians (Sumida et al., 2004) and carnivores (Chaveerach et al., 2008). Moreover, the genetic distances (Table 2) and dendrogram (Fig. 3) show that the genetic relationships within cercopithecids are closely related. The results are supported by other works using morphology (Lekagul and McNeely, 1988; Swindler, 1998; Falk, 2000; Francis, 2001) and g-Globin DNA sequences (Page et al., 1999).
The results from ITS alignment (Fig. 2) and dendrogram (Fig. 3) show that, within langur species, Trachypithecus phayrei is more closely related to T. obscurus than T. cristatus. This agrees with the morphological study (Brandon-Jones, 2006) and DNA fingerprinting study (Chaveerach et al., 2007) viewpoint, i.e., the T. phayrei is not a valid separate species. It is better to regard T. phayrei as a subspecies of T. obscurus.
| Table 1: | The GenBank accession number and ITS length of six cercopithecid species and Pan paniscus |
 | |
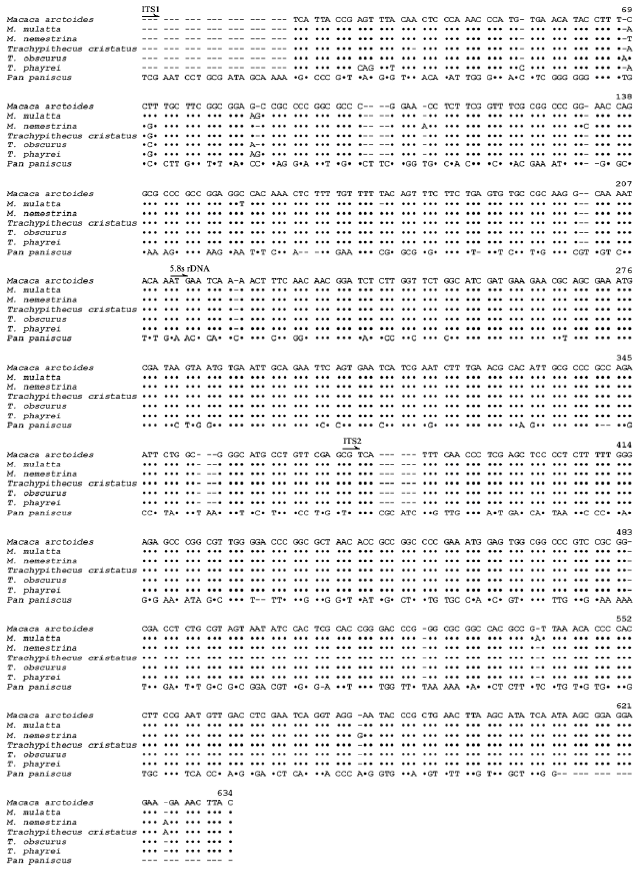 | |
| Fig. 2: | ITS sequence alignments from six cercopithecid species and Pan paniscus as an outgroup. Dots (•) indicate identical nucleotides, gaps (-) mean deletions of the bases |
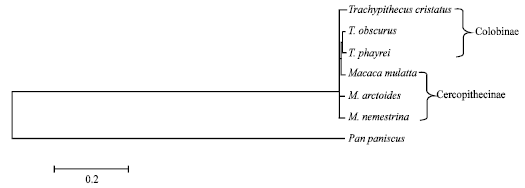 | |
| Fig. 3: | A dendrogram of six cercopithecid species and Pan paniscus as an outgroup constructed from sequence comparisons of the ITS regions using the Neighbor Joining Method (NJ) |
| Table 2: | Genetic distance values based on ITS sequences of six cercopithecid species and Pan paniscus |
 | |
Based on the morphology, the Cercopithecidae are divided into two extant subfamilies, Colobinae (the leaf monkeys) and Cercopithecinae (the cheek-pouch monkeys). They differ in morphological characters as described below. The colobine monkeys are distinguished by their specialized stomachs, which have been modified for a highly folivorous, or leaf-eating, diet. Along with this specialized diet, these leaf-eating monkeys have ruminant stomachs and a dental morphology suited for shearing leaves. Colobines are almost exclusively arboreal and possess a reduced or absent pollux and long, nonprehensile tails (Fleagle, 1999). In contrast, cercopithecines have well-developed thumbs and tails of varying lengths. These cheek-pouched monkeys are omnivorous, concentrating on fruit, but are also known to hunt on occasion (Swindler, 1998). They possess sacs in the buccal region of the oral cavity which are used to store food. Moreover, from the cytogenetic data, the diploid chromosome number of colobine species is 44 (Chaveerach et al., 2007) whereas the macaques have 42 as the diploid chromosome number (Tanee et al., 2006). Therefore, although the morphology and cytogenetics show high variability, the ITS region is highly conserved in these species. However, from the comparison between cercopithecids and the Pan paniscus (Hominoidae), the ITS1 and ITS2 regions show highly variable sequences.
From all of these results, the cercopithecid species have lower evolution than other species. This suggests that the rate of evolution may be different in different species groups. In addition, the low ITS variation suggested that the cercopithecid species may be at risk of extinction. Currently, the loss of specific, genetic and ecological diversities caused by human disturbance is a serious problem in Thailand. Forest destruction negatively affects wild animals such as cercopithecids. It reduces habitats for wild animals and causes population fragmentation. These isolated populations rapidly lose genetic heterogeneity and become vulnerable to environmental change and risk extinction. One more factor leading to genetic loss is the ability of interspecific intercross caused by close evolutionary relationship (Brown et al., 1986). The genetics of a species might be lost. In order to resolve the genetic relationships among cercopithecid species properly, it is necessary to study a larger sample, as well as to consider comparative behavioral studies.
REFERENCES
- Alvarez, I. and J.F. Wendel, 2003. Ribosomal its sequences and plant phylogenetic inference. Mol. Phylogenet. Evol., 29: 417-434.
CrossRefPubMedDirect Link - Brandon-Jones, D., 2006. The pros and cons of a consensus list of Asian primate subspecies. Primate Conserv., 20: 89-93.
CrossRefDirect Link - Brown, C.J., V.G. Dunbar and D.A. Shafer, 1986. A comparison of the karyotypes of 6 species of the genus Macaca and a species of the genus Cerocebus. Folia Primatol., 46: 172-174.
CrossRefPubMedDirect Link - Chaveerach, A., N. Srisamoot, S. Nuchadomrong, N. Sattayasai, P. Chaveerach, A. Tanomtong and K. Pinthong, 2008. Phylogenetic relationships of wildlife order Carnivora in Thailand inferred from the internal transcribed spacer region. J. Biol. Sci., 8: 278-287.
CrossRefDirect Link - Chaveerach, A., T. Tanee, N. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2007. Genetic relationships of langur species using AFLP markers. Pak. J. Biol. Sci., 10: 1445-1451.
CrossRefPubMedDirect Link - Insua, A., M.J. López-Piñón, R. Freire and J. Méndez, 2003. Sequence analysis of the ribosomal DNA internal transcribed spacer region in some scallop species (Mollusca: Bivalvia: Pectinidae). Genome, 46: 595-604.
CrossRefDirect Link - Page, S.L., C.H. Chiu and M. Goodman, 1999. Molecular phylogeny of old world monkeys (Cercopithecidae) as inferred from g-Globin DNA sequences. Mol. Phylogenet. Evol., 13: 348-359.
CrossRefPubMedDirect Link - Schlötterer, C., M.T. Hauser, A. Von Haeseler and D. Tautz, 1994. Comparative evolutionary analysis of rDNA ITS regions in Drosophila. Mol. Biol. Evol., 11: 513-522.
PubMedDirect Link - Sumida, M., Y. Kato and A. Kurabayashi, 2004. Sequencing and analysis of the internal transcribed spacers (ITSs) and coding regions in the EcoRI fragment of the ribosomal DNA of the Japanese pond frog Rana nigromaculata. Genes Genet. Syst., 79: 105-118.
CrossRefPubMedDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Tanee, T., A. Chaveerach, N. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2007. Analysis of the genetic relationships among Thai gibbon species using AFLP markers. Pak. J. Biol. Sci., 10: 1459-1464.
CrossRefPubMedDirect Link - Tsai, C.C., C.I. Peng, S.C. Huang, P.L. Huang and C.H. Chou, 2004. Determination of the genetic relationship of Dendrobium species (Orchidaceae) in Taiwan based on the sequence of the internal transcribed spacer of ribosomal DNA. Sci. Hort., 101: 315-325.
CrossRef - Tanee, T., A. Chaveerach, A. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2006. Cytogenetic and AFLP fingerprints for five species of Thai Macaques. Cytologia, 713: 303-308.
CrossRefDirect Link