
B. Masihipour
Razi Reference Laboratory of Scorpion Research, Razi Vaccine and Serum Research Institute, Ahvaz, Iran
A.M. Zare
2Department of Venomus Animals and Antivenin Production, Razi Vaccine and serum Research Institute, Karaj, Iran
S.H. Navidpour
Razi Reference Laboratory of Scorpion Research, Razi Vaccine and Serum Research Institute, Ahvaz, Iran
A. Taghavi Moghadam
Razi Reference Laboratory of Scorpion Research, Razi Vaccine and Serum Research Institute, Ahvaz, Iran
A. Akbari
Department of Venomus Animals and Antivenin Production, Razi Vaccine and serum Research Institute, Karaj, Iran
H. Rabiei
Department of Venomus Animals and Antivenin Production, Razi Vaccine and serum Research Institute, Karaj, Iran
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 5 | Page No.: 494-498







ABSTRACT
Odonthobuthus belongs to Buthidae family from which 2 species O.doriea and O. bidentatus have been collected and reported from Iran. This study investigated the venom of O. bidentatus to identify mammal toxic fractions. After collecting scorpions, they were milked with electroshock technique and then the venoms was lyophilized. The LD50 of the venom was determined by injecting the venom to 18-20 g mice via., tail vein. In order to separate different fractions, the freeze dried venom was solubilized in distilled water and centrifuged at 15000 rpm for 20 min, to separate insoluble material and the clear supernatant containing soluble venom was loaded on a chromatography column packed with sephadex G50 gel and the fractions were collected according to UV absorption at 280 nm wavelength. Second fraction was toxic in Lab animal and in order to study sub fractions, fraction II (O2) was loaded first on an anionic ion exchange resin (DEAE sephacel) out of which we obtained 2 toxic fractions (O21 and O23). Then these 2 fraction were loaded on a cationic resin (CM sephadex C25) out of which, from fraction 21, we got 2 toxic fractions (O211, O213) and from fraction O23 we got one toxic fraction (O232). SDS PAGE electrophoretic studies on these toxins showed single band appearance that indicates purified toxins with molecular weight of O211 9.2, O213 8.6 and O233 6.8 kD.
PDF Abstract XML References Citation
How to cite this article
B. Masihipour, A.M. Zare, S.H. Navidpour, A. Taghavi Moghadam, A. Akbari and H. Rabiei, 2009. Identification and Purification of the Three Toxins from Odotobuthus bidentatus, Lourenco and Pezier 2002, Venom (Scorpions, Buthidae). Journal of Biological Sciences, 9: 494-498.
DOI: 10.3923/jbs.2009.494.498
URL: https://scialert.net/abstract/?doi=jbs.2009.494.498
DOI: 10.3923/jbs.2009.494.498
URL: https://scialert.net/abstract/?doi=jbs.2009.494.498
INTRODUCTION
Scorpion venom is a rich source of various polypeptides, which are proved to be high affinity ligands of ion channels making them useful as pharmacological probes. Among the well characterized peptide toxins are those from scorpions belonging to Buthidae family (Chirstian et al., 1996; Nastain, 2002; Rajendra, 2001). The lyophilized milked venom was a matrix containing glycoprotein, cellular debris and mucus material (Gawade, 2003) and various concentration of soluble material like neurotoxin, cardiotoxin, nephrotoxin, hyoluronidases, glycosaminoglacans, histamine (Chirstian, 1996; David, 2005). Also, source of highly concentrated active enzymes and cytotoxine (Badhe, 2006). The classification of scorpion toxins have been made on various criteria such as ion channel specificity, number of amino acid residues or target specificity (Cook et al., 2002; Srinivasan, 2002). Odontobuthus is one of the medically important buthid scorpion in Iran (Zare et al., 2007). Navidpour et al. (2008) showed that in South and Southwestern provinces of Iran there are 2 species of Odontobuthus: O. doriae and O. bidentatus. Biochemical and clinical manifestations as well as toxins identification and purification from the venom of O. doriae was previously reported by Zare et al. (2006). However, no report exist about the toxins present in O. bidentatus. Hence, in this study we attempt to identify and purify the mammalian toxins present in O. bidentatus.
MATERIALS AND METHODS
The scorpions were collected with UV light at night (Lowe et al., 2003) from different parts of Khoozestan Province and they were milked by electric shock at the end of tail. The venom was water dialyzed and freeze dried at -20°C until use (Debont, 1998). The LD50 was determined by sperman and carber method on albino mice weighting 18- 20 g (Masihipour, 2005; Kalapothakis and Chavez, 1997).
Preparation of the sample: The freeze dried venom (812 mg) was dissolved in distilled water and placed in dialysis bag and dialyzed against distilled water at 4°C for 48 h (Masihipour, 2005) after dialysis the venom solution was centrifuged at 15000 rpm for 15 min and supernatant was collected (Kalapothakis and Chavez, 1997).
Sephadex gel filtration: Crude venom loaded over a sephadex G50 (fine) (More et al., 2005) in a 100x50 cm column equilibrated with ammonium acetate buffer (0.1 mol and pH = 7) and flow rate was adjusted to 60 mL h-1 (Davila, 1996). Sample collection was carried out in 10 mL tubes and protein content was determined by UV spectrometer at 280 nm wave length. Each fractions toxicity was tested on mice by intravenous injection of 0.5 mL of each fraction to albino mice weighting 18-20 g.
Ion exchange chromatography: Toxic fraction derived from gel filtration was load on DEAE sephacel an anionic ion exchanger (0.1 mL, pH = 8.9) and flow rate was adjusted to 30 mL h-1. The elusion was carried out using gradient of NaCl (0.0 -0.05 mL). The eluted fractions from this column after testing on albino mice and finding the toxic fractions as previously mentioned were load on a CM sephadex C25 an cation exchanger and toxic fractions were collected and checked for toxicity.
SDS PAGE electrophoresis: Crude venom as well as each toxic fraction and finally the purified toxins were loaded on SDS PAGE electrophoresis to determines the trend of purification. The concentration percentage of SDS-PAGE was 10% for crude venom and 12% for pure toxins.
RESULTS AND DISCUSSION
The toxicity of venom was tested by determination of LD50 of crude venom in mice for 24 h. the average LD50 of crude venom was found to be 10.35 μg.
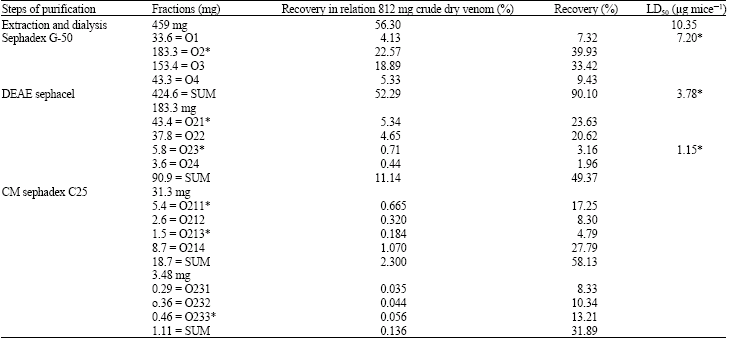
Total protein of the crude venom was found to be 353 mg after removal of mucoproteins. That means percentage of mucoprotein in the crude venom using electroshock method for venom milking in 47.3% (Table 1). Electrophoretic pattern of electrophoresis showed the presence of at least 14 separated bands (Fig. 1). Total amount of protein loaded on the gel filtration column was 459 mg. After elution from gel filtration column we got 4 peaks (Fig. 2) out of which the peaks No O2 was found to be toxic on mice.
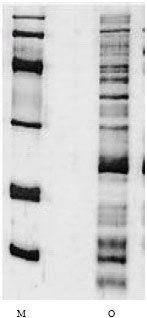 | |
| Fig. 1: | Comparison of electrophoretic bands of O. bidentatus with control marker |
| Table 1: | Percentage recovery and LD50 of extracted toxins |
 | |
| *Fractions which killed mice | |
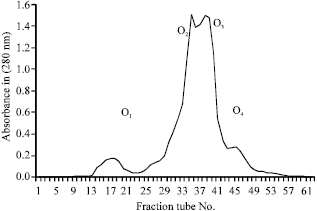 | |
| Fig. 2: | Sephadex G-50 gel filtration crude venom on a 2.5x125 cm column with 0.1 M ammonium acetate buffer, pH = 8.6 and 60-70 mL h-1 elution rate |
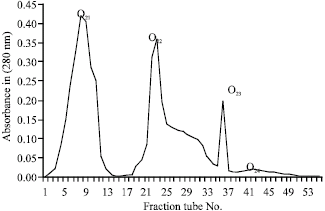 | |
| Fig. 3: | DEAE sephacel ion exchange chromatography of O2 fraction on a 1.3x50 cm column with 0.5 mol ammonium acetate buffer with 0.5 mol salt gradient, pH = 8.9 and 30-35 mL h-1 elution rate |
| Table 2: | Results obtained from IV injection of 0.5 mL of each fractions to mice after 24 h |
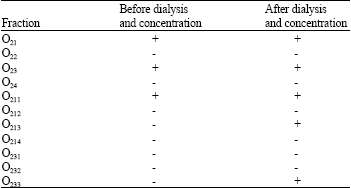 | |
| +: Kill the mice -: Non kill the mice | |
The total protein of O2 were determined and found to be 183 mg. The LD50 for O2 were found to be 7.2 μg, respectively (Table 1). Yield of mammalian toxins for O2 were, DEAE sephacel anionic chromatography of fraction O2 from previous step, four fractions O21, O22, O23 and O24 were obtained (Fig. 3) after injection of these 4 fractions to animals it was found that fractions O21 and O23 are toxic (Table 1).
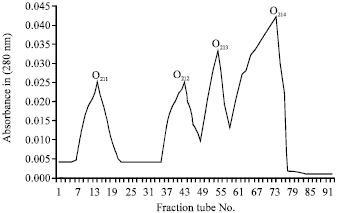 | |
| Fig. 4: | CM sephadex O25 ion exchange chromatography of O2.1 fraction on a 1.5x50 cm column with 0.1 Tri sodium citrate buffer and 0-0.5 mol salt gradient and 30-35 mL h-1 elution rate |
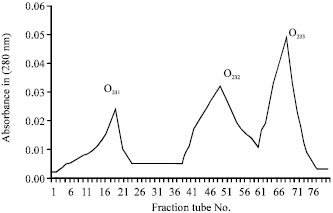 | |
| Fig. 5: | CM sephadex O25 ion exchange chromatography of O2.3 fraction on a 1.5x50 cm column with 0.1 mol Tri sodium citrate buffer and 0-0.5 mol salt gradient and 30-35 mL h-1 elution rate |
After that, the 2 toxic fraction from above step, were loaded on CM sephadex C25 column, from O21 fraction 4 sub fractions O2I1,O2I2, O2I3, O2I4 (Fig. 4) were eluted containing 5.4, 2.6, 1.5 and 8.7 mg protein, respectively (Table 1). After injection of these fractions to mice, it was clear that, 2 fractions O2I1 and O2I3 were toxic (Table 2). Fraction O23 after elution on the CM sephadex O25 gave three sub fractions O23I, O23II, O23III (Fig. 5) with protein content of 0.29, 0.36 and 0.46 mg, respectively (Table 1), out of which only fraction O23III was toxic (Table 1). Electrophoretic pattern of purified toxins showed a single band which was an indicator for purity of toxins.
The present study examined, the toxic fractions and subfractions present in the venom of O. bidentatus which has a wide distribution in South Provinces of Iran and is one of the medically important scorpion in the region. The envenomation by this scorpion cause local pain accompanied with extreme edema in severe cases it manifests dizziness, delirium vomiting nausea and finally sleepiness. The primary action of venom is through both sympathetic and parasympathic postganglionic stimulation (Dittrich et al., 2002). Four different fractions were obtained by sephadex G50 (fine) chromatography, out of which, second fraction O2 was toxic for lab animal and other fraction were not toxic for mammals even though they can be toxic for insects. In a research Zare et al. (2006) separated three hommogen toxic fraction from the venom of scorpion (Odonthobuthus doriea) the O2 fraction from previous step was loaded on a DEAE sephacel anion exchange column. Four Sub fractions were eluted, out of which O21 and O23 were toxic. Then sub fractions toxic were loaded on a CM sephadex O25 cation exchange column from O21 fraction 2 toxic peptide i.e., 211 and 213 were obtained and from O23 one toxic peptide was elutedie 233. The omittion of higher molecular bands from O2 fraction after G50 sephdex gel chromatography, was due to lack of toxicity but O2 fraction which was toxic, had low molecular weight. Electrophoretic results showed that O211, O213 and O233 are homogen. It is necessary to notify that presence of single proteinic band on gel is necessary to proof homogenecity but is not enough. Because neurotoxins purified from a venom may have similar appearance locations and approval of final purity of toxins and the determination of their exact molecular weight needs amino acid analysis in further researches. Usually, toxins with high activities are found in the venoms of Buthidae family which are effective on ionic channels ie Na, K, Cl channels in different tissues. This study showed different toxic fractions of O. bidentatus venom which are specifically active on mammals, thus serving as important pharmaceutical tools for excitability and sodium channel structure and serve as specific targets for many neurotoxins. These toxins occupy different receptor sites and have been used as tools for functional mapping and characterization of the channels. Scorpion toxins affecting inactivation of sodium current may be divided on different groups according to their mammal or insect activities. The first group comprise highest activity on mammals sodium channels and lowest affinity for insect neuronal membrane. Second group are very weakly active on insects but express its toxic activity mainly on mammals. Third group are active on both insects and mammals, fourth group which oppose low affinity to mammals and high affinity for insects. Clarification of these different selectively in action of toxins requires three dimensional structural knowledge of the toxins coupled with molecular localization of amino acids directly interacting with recognition points which is suggested to be important areas of future studies.
ACKNOWLEDGMENTS
We wish to thank Mr. Hayader, Mr. Habibzadeh, Mr. Bahrani and Mr. Rabiei for their helpful support.
REFERENCES
- Badhe, R.V., A.B. Thomas, S.L. Harer, A.D.S. Deshpande and A. Waghmare, 2006. Intraspecific variation in protein pattern of red scorpion (Mesobuthus tamulus coconsis, pocock) venoms from Western and Southern India. J. Venom. Anim. Toxins Incl. Trop. Dis., 12: 612-619.
CrossRefDirect Link - Cook, J.W., Z. Alan, D.D. Watt and S.E. Ealick, 2002. Structure of variant 2 scorpion toxin from Centruroides sculpturatus Ewing. Protein Sci., 11: 479-486.
Direct Link - Dehesa-Davila, M., A.N. Ramfrez, F.Z. Zamudio, G. Gurrola-Briones, A. Lievano, A. Darszon and L.D. Possani, 1996. Structural and functional comparison of toxins from the venom of the scorpions Centruroides infamatus infamatus, centruroides limpidus limpidus and Centruroides noxius. Comp. Biochem, Physiol., 113B: 331-339.
CrossRefDirect Link - Debont, T., A Swerts, J.J. Van-der-Walt, G.J. Muller, F. Verdonck, P. Daenens and J. Tytgat, 1998. Comparison and characterization of the venoms of three parabuthus Scorpion species occurring in Southern Africa. Toxicon, 36: 341-352.
Direct Link - Dittrich, K., R. Ahmad and Q.A. Ahmed, 2002. Cardiac arrest following scorpion envenomation. Ann. Saudi Med., 22: 87-90.
PubMedDirect Link - Gawade, S.P., 2003. Excitatory effects of buthus C56, toxin on drosophilia larval neuro muscular junction. J. Venomous Anim. Toxins Includ. Trop. Dis., 9: 65-75.
Direct Link - Lowe, C., S.R. Kutcher and D. Edwards, 2003. A powerful newlight source for ultraviolet detection of scorpion in the filed. Euscorpius, 8: 1-7.
Direct Link - Kalapothakis, E. and C. Chavez-Olortegui, 1997. Venom variability among several tityus serrulatus specimens. Toxicon, 35: 1523-1529.
CrossRefDirect Link - More, S.S., K.K. Mirajkar, J.R. Gadag, K.S. Menon and K. Mathew, 2005. A novel Kv 1.1 potassium channel blocking toxin from the venom of palamneus gravimanus (Indian black scorpion). J. Venom. Anim. Toxins Includ. Trop. Dis., 11: 315-355.
CrossRefDirect Link - Rajendra, W., I. Bora, H. Rafael, D. Maher, P.V. Choudary and D. Bruce Hammock, 2001. Isolation and characterization of novel lepidopteran-selective toxin from the venom of south Indian red scorpion, mesobuthus tamulus. BMC Biochem., 2: 16-16.
CrossRefDirect Link - Srinivasan, K.N., P.T. Gopalakrishnakone, K.C. Tan, B. Chew and R.M. Cheng et al., 2002. Scorpion, a molecular database of scorpion toxins. Toxicon, 40: 23-31.
Direct Link - Mirakabadi, A.Z., A. Jalali, A.E. Jahromi, H. Vatanpur and A. Akbary, 2006. Biochemical changes and manifestations of envenomation produced by Odonthobuthus doriae venom in rabbits. J. Venom. Anim. Toxins Includ. Trop. Dis., 12: 67-77.
CrossRefDirect Link - Zare, A., H. Zolfagharian, A. Hedayat and A. Jalali, 2007. Clinical and biochemical manifestations produced by scorpion (Hemiscorpius lepturus) venom in experimental animals. J. Venom. Anim. Toxins Includ. Trop. Dis., 13: 758-765.
CrossRefDirect Link - Christian, L., E.C. Brigitte, E.B. Pierre and M.E. Marie France, 1996. Evidence for new class of scorpion toxins active agains K+ channels. FEBS Lett., 431: 375-380.
PubMed - Nastain, C.W., 2002. A short chain peptide toxin isolated from centruroides sculpturatus scorpion venom inhibits ether a-go-related gene K+ channels. Toxicon, 40: 1053-1058.
PubMed