
Roushon Ara
Laboratory of Marine Science and Aquaculture, Institute of Bioscience, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A. Arshad
Laboratory of Marine Science and Aquaculture, Institute of Bioscience, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
N. Amrullah
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
S.M. Nurul Amin
Laboratory of Marine Science and Aquaculture, Institute of Bioscience, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
S.K. Daud
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A.A. Nor Azwady
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A.G. Mazlan
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 5 | Page No.: 445-451







ABSTRACT
Diet composition of most common fish larvae of family Sparidae was studied in the Sungai Pulai seagrass bed of Gelang Patah South Western part of Johor, Peninsular Malaysia from December 2007 to May 2008. Larvae were collected by subsurface towing of a bongo net. In situ hydrographic parameters were recorded during the sampling cruises. Stomachs were removed from a total of 80 Sparidae specimens during the study period and the stomach contents were examined. Analysis of prey in the stomach showed 24 important food items belonging to 8 major groups: phytoplankton, zooplankton, algae, insects, plant like matter, decapod appendages, debris and unidentified matters. The predominant food items found in the stomach were phytoplankton (60.85%). This was followed by algae (11.73%), zooplankton (9.35%), plant matters (7.84%), debris (4.60%), insects (2.84%), unidentified matters (1.96%) and decapod appendages (0.82%). Habitat in situ temperatures were recorded at 26.92-30.83°C (Mean± SD, 28.60± 1.38); Dissolved oxygen ranged from 4.73 to 6.24 mg L-1 (5.56± 0.53) and the salinity fluctuation was between 29.37 and 33.68 ppt (31.31± 1.68). Among the food items, phytoplankton was the first rank by Simple resultant index (60.85%) followed by algae (11.73%). Therefore, it could be concluded that the fish larvae of family sparidae are mainly herbivorous.
PDF Abstract XML References Citation
How to cite this article
Roushon Ara, A. Arshad, N. Amrullah, S.M. Nurul Amin, S.K. Daud, A.A. Nor Azwady and A.G. Mazlan, 2009. Feeding Habits and Temporal Variation of Diet Composition of Fish Larvae (Osteichthyes: Sparidae) in the Sungai Pulai Seagrass Bed, Johore, Peninsular Malaysia. Journal of Biological Sciences, 9: 445-451.
DOI: 10.3923/jbs.2009.445.451
URL: https://scialert.net/abstract/?doi=jbs.2009.445.451
DOI: 10.3923/jbs.2009.445.451
URL: https://scialert.net/abstract/?doi=jbs.2009.445.451
INTRODUCTION
The importance of seagrass beds as nursery habitat for commercially important species has reached almost paradigm status, despite few studying mentioning the proportion of fish from seagrass habitats that were early juveniles. For example, in a study of a Guatemalan seagrass beds, 83% of fishes were early juveniles stages (Arrivillaga and Baltz, 1999) and in experiment in Botany Bay (NSW, Australia) 80% of individuals and 8 of the 13 species were juveniles of species found only on coastal reefs as adult. Therefore, seagrass beds are important feeding habitats for juvenile and adult fishes. A dense vegetation of seagrass produces a great quantity of organic material. This may offer a good substrate for smaller algae, diatoms and sessile fauna. Organisms that usually can be found in the seagrass beds are like diatom, hydroids, copepods, amphipods, gastropods, isopods, carid shrimps and fishes (McRoy and Helfferich, 1977).
Studies of larval fishes are often the best way to provide information of great value to fishery biologists and managers of fisheries. These include location of spawning grounds in space and time, determination of habitats used by fish during their larval phase, fishery independent estimates of stock size and stock boundaries, discovery of new fisheries, feeding habits of larvae, condition of larvae, insight into recruitment fluctuations and historical changes in all of the above. Fish larvae must optimize their food intake by being effective prey searchers and predators in order to maintain a high growth rate and survive to recruitment (Ostergaard et al., 2005).
Different fishes consume different types of food and feeding habit of fishes varies from season to season. Defining dietary preferences for the early stages is therefore an important element in the assessment of feeding conditions and larval chances of meeting food requirement (Robichaud-Le Blanc et al., 1997). Observations on larval prey preferences are mainly based on analyses of gut contents. A lack of empirical data on the diets of fish larvae in the wild, however, again leads to a reliance on results from aquaculture study (Humphries et al., 1999). The knowledge about the specification of food items both qualitatively and quantitatively from the present study can be applied in aquaculture development. Various studies have been carried out in terms of food and feeding habits of the adult fishes (Chrisfi et al., 2007; Dadzie et al., 2000; Jardas et al., 2007), but little known about the diet of fish larvae in seagrass beds.
 | |
| Fig. 1: | Geographical location of the sampling site (±) in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
The aim of the present study was to assess the feeding habits and diet composition of larval fishes of most common family Sparidae in the seagrass beds of Sungai Pulai estuary, Johore Striat, Peninsular Malaysia. In terms of management, such studies are essential for evaluation of the ecological role of the fish larvae as well as the understanding of its position in the food web structure in the seagrass ecosystem.
MATERIALS AND METHODS
Samples collection: Monthly samples of fish larvae were collected from Sungai Pulai Seagrass Bed, Johor (N 01° 19’.414” E103° 35.628”) between December 2007 and May 2008 (Fig. 1). Fish larvae were collected by subsurface towing of a bongo net (mesh size of 500 μm). The collected fish larvae samples were preserved with 5% formalin in the container. In situ hydrographic parameters such as water temperature, salinity and dissolved oxygen were recorded by YSI (556 MPS, USA) during the sampling cruises.
Stomach examination: In the laboratory, the fish larvae were separated and identified to family level using Nikon dissecting microscope and Leis and Ewart (2000). The larval fishes of most abundant family Sparidae (Fig. 2) were separated from the other family and zooplankton. Then the total length and standard length of the specimen of Sparidae larvae were taken using digital microscope before the specimens were dissected. The individual larva was laid on a slide before a drop of distilled water was dripped onto it. Then the stomach was carefully removed from the body by a probe under a dissecting microscope. After that, the body was removed from the slide and the stomach was shattered using a fine needle. The slide was covered by a cover glass. Then the stomach contents were identified to the possible lowest systematic level and counted under a compound microscope.
Stomach content analysis: To analyze the composition of the stomach, percentage frequency of occurrence and percentage numerical abundance was followed which as:
where N1i is the number of the stomachs in which food itemi was found and Np, the number of non-empty stomachs (Chrisfi et al., 2007).
where ni is the number of ith food item, m the number of food items (Chrisfi et al., 2007).
The relative importance of food item on the diet of larva was assessed by Simple Resultant Index (%Rs) of the ith food item proposed by Mohan and Sankaran (1988).
where Ci is the percentage numerical abundance and Fpi the percentage frequency of occurrence.
RESULTS AND DISCUSSION
Environmental variables: The mean value of salinity recorded in the Sungai Pulai Seagrass Beds, Johor was 31.31± 1.68 ppt (Mean± SD). The lowest value of salinity recorded was 29.37 ppt during April; while the highest value of salinity recorded was 33.68 ppt during February.
 | |
| Fig. 2: | Fish larvae (50x) of family Sparidae collected from the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
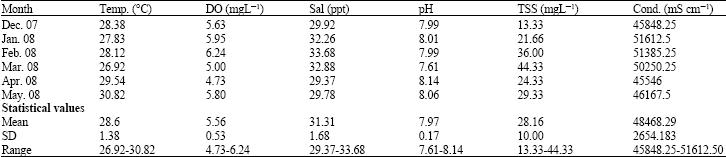
Water temperature was relatively constant between 26.92 and 30.82°C (28.60± 1.38°C) and dissolved oxygen was lowest (4.73 mg O2 L-1) in April and highest (6.24 mg O2 L-1) in February (Table 1). Hydrogen ion concentration varied from 7.61 (March) to 8.14 (April). Maximum values of Total Suspended Solid (TSS) was recorded in March (44.33 mg L-1) and the minimum in December (13.33 mg L-1) during the study period (Table 1). The mean value of conductivity recorded was 48468.29± 2654.18 mS cm-1 and range between 45848.25 and 51612.50 mS cm-1.
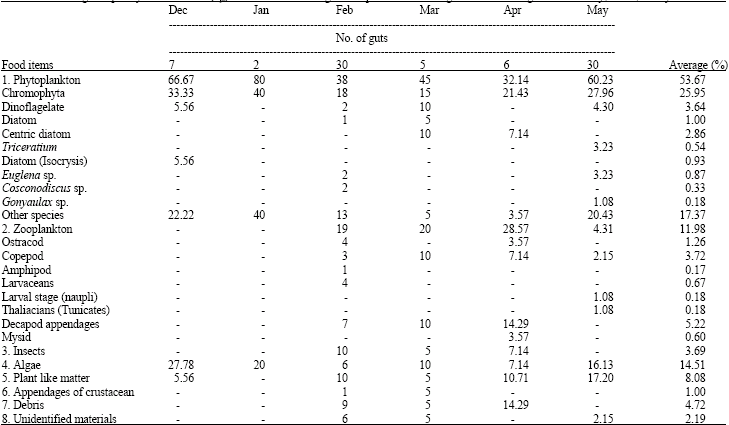
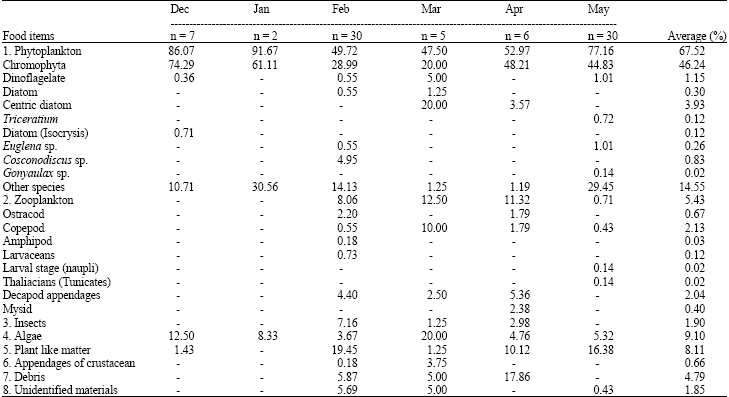
Diet composition: Analysis of prey in the stomachs identified 24 important items belonging to 8 major groups (Table 2): phytoplankton, zooplankton, insect, algae, plant like matter, decapods appendages, debris and unidentified materials. The monthly percentage of occurrence (Fpi) of food items in the guts of fish larvae from Sparidae family are presented in Table 3. The average most important preys in the stomachs according the frequency of occurrence were phytoplankton (53.67%). This was followed by algae (14.51%), zooplankton (11.98%), plant like matter (8.08%), debris (4.72%), insects (3.69%), unidentified species (2.19%) and appendages of crustaceans (1%). Monthly percentage of numerical abundance (Ci) of food items in the guts of fish larvae are The Table 4 also showed that the predominant item was phytoplankton (67.52%) and presented in Table 4. followed by algae (9.10%), plant like matter (8.11%), zooplankton (5.43%), debris (4.79%), insects (1.90%), unidentified species (1.85%) and appendages of crustaceans (0.66%).
Overall diet compositions of the fish larvae of Sparidae ranked by Simple Resultant Index (Rs) are presented Table 5. Analyses of prey in the stomachs identified that the most important prey according the Simple Resultant Index (Rs) were phytoplankton (60.85%). This was followed by algae (11.73%), zooplankton (9.35%), plant matter (7.84%), debris (4.60%), insects (2.84%), unidentified species (1.96%) and appendages of crustacean (0.82%).
Monthly variation of diet: The highest percentage of occurrence of phytoplankton was found in January (80%), zooplankton in April (28.57%) and algae in December (27.78%). Meanwhile for insects, the highest percentage of occurrence was in February (10%) and plant like matter was in May (17.20%). Percentage occurrence of crustacean appendages was highest in March (5%), debris in April (14.29%) and unidentified species was highest in February (6%) (Table 3). The highest percentage of numerical abundance of phytoplankton was found in January (91.67%), zooplankton in March (12.50%), algae in December (12.50%), plant matter in February (19.45%) and appendages of crustacean in March (3.75%) (Table 4). The insects were highest in February (7.16%), debris in April (17.86%) and unidentified species in February (5.69%) (Table 4).
Our study indicates that the fish larvae of Sparidae are exclusively herbivore. According to the obtained data its main prey are the phytoplankton. This prey group represents more than 50% of total Rs (Table 5). The fish larvae appeared to select food of a certain size range varying with the size of the larvae.
| Table 1: | Monthly variation of different water parameters in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
 | |
| Table 2: | Major food items found in the stomach of fish larvae in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
 | |
| Table 3: | Percentage frequency of occurrence (Fpi) of food items in the guts of Sparidae in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
 | |
| Table 4: | Percentage of numerical abundance (Ci) of food items in the guts of Sparidae in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
 | |
| n : No. of guts | |
It is observed that the total length and standard length of fish larvae were between 2.0 and 4.0 mm. Marak (1973) reported that adult copepods approximately 0.7 to 3.5 mm in size. Thus, copepods could be accepted as one of the diets for Sparidae fish larvae. Based on the percentage of frequency of occurrence, phytoplankton is the most dominant item ingested by the larvae was Chromophyta.
| Table 5: | Overall diet composition of Sparidae ranked by Simple Resultant Index (%Rs) in the seagrass beds of Sungai Pulai estuary, Johor, Malaysia |
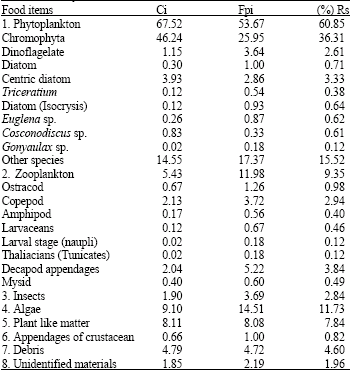 | |
However, when compared to the diet composition of anchovy larvae (Hunter and Thomas, 1972) which the main item of phytoplankton was Gymnodium splendens (Dinoflagellate). May be Chromophyta are the dominant in the seagrass ecosystem of Sungai Pulai estuary and Sparidae larvae prefer this phytoplankton. Instead of phytoplankton, the other important food item consumed by the larvae is algae. Algae were in the second rank (14.51%) for the family of Sparidae. Zooplankton was the thirdly important in the diet of larvae of Sparidae which 11.98%. The larvae of plaice and sand eels feed on appendicularians (larvacean) during the early part of their life (Ryland, 1964; Shelbourne, 1962). Basically, the plant matter consists of dried roots, stems, grass leaves and part of unidentifiable plants. The plant matter was frequently found in the stomachs of the larvae which (8.08%) of Sparidae. Recent study on food and feeding habits of Acetes indicus from the coastal water of Malacca indicated that the species was omnivorou where the main food items was plant matter (Amin et al., 2007). Debris (4.72%) was also one of the food items found in stomachs of larvae as well as insects (3.69%). The seagrass bed area may be also inhabited by insects instead of phytoplankton and zooplankton which usually can be found there. However, debris could not be considered as important food item for fish larvae. The less frequent of food item found in the stomach of larvae were unidentified species (2.19%) and appendages of crustacean (1%).
CONCLUSION
The variety of food consumed by fish larvae of Sparidae family though phytoplankton was most frequent indicates that they are mainly herbivorous. Since the availability of suitable food is one of the important factors influencing the survival of larval fish, the knowledge from this study is hope will be provide information for fisheries development and aquaculture field. Also, the knowledge can be used for further study from any other area instead of seagrass beds.
REFERENCES
- Arrivillaga, A. and D.M. Baltz, 1999. Comparison of fishes and macroinvertebrates on seagrass and bare-sand sites on Guatemala's Atlantic coast. Bull. Mar. Sci., 65: 301-319.
Direct Link - Chrisafi, P., P. Kaspiris and G. Katselis, 2007. Feeding habits of sand smelt (Atherina boyeri, Risso 1810) in Tichonis Lake (Western Greece). J. Applied Ichthyol., 23: 209-214.
CrossRef - Dadzie, S., F. Abou-Seedo and E. Al-Qattar, 2000. The food and feeding habits of the silver pomfret, Pampus argenteus (Euphrasen), in Kuwait waters. J. Applied Ichthyol., 16: 61-67.
CrossRefDirect Link - Humphries, P., A.J. King and J.D. Koehn, 1999. Fish, flows and floodplains: Links between freshwater fish and their environment in the Murray-Darling Rive system, Australia. Environ. Biol. Fish., 56: 129-151.
CrossRef - Mohan, M.V. and T.M. Sankaran, 1988. Two new indices for stomach content analysis of fishes. J. Fish Biol., 33: 289-292.
CrossRef - Ostergaard, P., P. Munk and V. Janekarn, 2005. Contrasting feeding patterns among species of fish larvae from the tropical Andaman Sea. Mar. Biol., 146: 595-606.
Direct Link