
K. Nadarajah
Faculty of Science and Technology, School of Biosciences and Biotechnology, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
N.M. Kasim
Faculty of Science and Technology, School of Biosciences and Biotechnology, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
V.V. Fui
Department of Biotechnology, Faculty of Applied Sciences, UCSI University, No. 1 Jalan Menara Gading. UCSI Heights, 56000 Kuala Lumpur, Malaysia
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 5 | Page No.: 402-412







ABSTRACT
The effect of salinity, wounding and drought on the expression of a MAPK gene in rice was analyzed in this research. OrMKK2, an ortholog of the Oryza sativa japonica OsMAP1 (OsMPK5), was isolated and characterized in this study. This gene is 1381 bp long and has a 5’UTR that is 120 bp and a 3’UTR that is 195 bp and it encodes a protein that is 356 amino acids long. When analyzed against a previously isolated OrMKK1, the gene exhibited similarities in the 11 domain locations and showed 96% identity to OrMKK1. Unlike OrMKK1, the OrMKK2 gene contained two copies of the gene in the genome. OrMKK2 appeared to be positively regulated by abiotic stress.
PDF Abstract XML References Citation
How to cite this article
K. Nadarajah, N.M. Kasim and V.V. Fui, 2009. The Modulation of Abiotic Stresses by Mitogen-Activated Protein Kinase in Rice. Journal of Biological Sciences, 9: 402-412.
DOI: 10.3923/jbs.2009.402.412
URL: https://scialert.net/abstract/?doi=jbs.2009.402.412
DOI: 10.3923/jbs.2009.402.412
URL: https://scialert.net/abstract/?doi=jbs.2009.402.412
INTRODUCTION
Rice plants are exposed to diverse external stimuli during their life cycle, including pathogen infection, drought, salinity and various other biotic and abiotic stress. These extracellular stimuli are perceived by the plant through an elaborate signaling network, which elicits a stress response at cellular and organism level. Mitogen-Activated Protein Kinases (MAPK) are a group of serine/threonine-specific protein kinases that have been conserved throughout eukaryotes (Yang et al., 2001). Mitogen-Activated Protein Kinases respond to various extracellular stimuli and act as important signaling components that regulate various cellular activities, such as cell division, differentiation and programmed cell death. As the terminal module of the MAPK cascade, MAPKs are activated by their upstream MAP Kinase Kinase (MAPKK) and MAP Kinase Kinase Kinase (MAPKKK). Mitogen-Activated Protein Kinases can either phosphorylate a substrate or target protein in the cytoplasm or can translocate into the nucleus and eventually regulate gene expression through phosphorylation of specific transcription factors (Jin et al., 2002). A particular MAPK cascade may mediate a multitude of extracellular stimuli and result in a variety of specific responses by regulating a number of substrates or target proteins. The MAPK-mediated signal transduction is not just a simple linear process, but involves complicated cross talks with other signaling pathways (Jonak et al., 1996; Zhang and Klessig, 2001). Recently, a total of 17 rice genes encoding MAP kinases have been identified from the rice genome (Hamel et al., 2006; Reyna and Yang, 2006). The deduced protein products of these genes all share the conserved protein kinase domains I–XI.
In all eukaryotes, receptor mediated activation of a MAPKKK can occur through physical interaction and/or phosphorylation by either the receptor itself, intermediate bridging factors or interlinking MAPKKKs (Frye et al., 2001; Zhang and Liu, 2001). As serine/threonine kinases, MAPKKKs activate MAPKKs through the phosphorylation of two serine/threonine residues in a conserved S/T–X3–5–S/T motif. By contrast, MAPKKs are dual-specificity kinases that phosphorylate MAPKs on threonine and tyrosine residues in the T–X–Y motif. MAPKs phosphorylate a variety of substrates including transcription factors, protein kinases and cytoskeleton- associated proteins. The specificity of different MAPK cascades functioning within the same cell is generated through the presence of the above mentioned docking domains found in various components of MAPK modules and scaffold proteins (Xiong and Yang, 2003; Ichimura et al., 2002).
Although, detailed steps of MAPK cascades have yet to be elucidated in a given plant species, specific upstream MAPKKs for a few well-characterized MAPKs have been determined. A complete MAPK cascade together with its upstream receptor kinase and downstream transcription factors, was characterized in Arabidopsis (Asai et al., 2002). These findings suggest that MAPKs are an important signaling components in plant defense responses and that the cascade of a three kinase module is a general mechanism of defense signal transduction among eukaryotic organisms (Ligterink et al., 1997; Ligterink and Hirt, 2000).
To date, a total of 18 MAPK genes have been identified from the rice genome. Expression profiling, biochemical characterization and/or functional analysis were carried out with many members of the rice MAPK gene family, especially those associated with biotic and abiotic stress responses in mostly dicot model species such as Arabidopsis and tobacco. He et al. (1999) reported that a 60 kDa MAPK (OsBWMK1) was activated in rice leaves by blast fungus infection and wounding. Most recently, a stress-responsive MAPK gene (variously named OsMKK2, OsMSRMK2, OsMAPK2, OsMAP1 and OsBIMK1) was identified by at least six laboratories and shown to be induced at the mRNA level by multiple biotic and abiotic stress (Xiong et al., 2001; Agrawal et al., 2002; Song and Goodman, 2002; Zhang and Liu, 2001).
Here, we report the molecular characterization of OrMKK2, a stress-responsive MAPK gene that was isolated from Oryza rufipogon (designated OrMKK2 based on the order of clones analyzed from the library).
MATERIALS AND METHODS
Isolation and sequence analysis of OrMKK2: A cDNA fragment of OrMKK2 was isolated (unpublished results) by suppression subtractive hybridization, biotin labeled and used as a probe to isolate the corresponding full-length cDNA. Approximately 106 plaques from a blast fungus (Magnaporthe grisea) induced cDNA library were screened. The resulting positive clones carrying OrMKK2 cDNAs were excised in vivo from the ZAP express vector (Stratagene, La Jolla, CA). Subsequently, the full-length OrMKK2 cDNA clone was sequenced from both directions using a primer-walking approach. Automated sequencing service was provided by Research Biolab, Malaysia.
Plant materials: Transgenic rice plantlets (5 to 6 cm in height) from the rooting medium were transplanted into earth soil mix and grown in a 28°C greenhouse with a 16 h light/8 h dark cycle. T1 plants carrying the transgenes were selected by germinating seeds on filter paper soaked with 50 μg mL-1hygromycin. Non transgenic seeds did not germinate in the presence of 50 μg mL-1hygromycin. Positive T1 plants were confirmed by PCR using primers corresponding to the OrMKK2 gene.
Gene constructs and rice transformation: The pCaM-OrMKK2 was constructed by directionally inserting the full cDNA sequence (digested with BamHI and XbaI) into the vector pCAMBIA1300. The OrMKK2 PCR product was generated by PCR with primers OrMKK2 For (5-CGGGATCCGTCGGCTGCATCTTCATG-3) and OrMKK2Rev (5- GCTCTAGATTCAATCTAGTACCGGA-3). The PCR product was digested with BamHI and XbaI and then inserted into pCAMBIA1300 before being introduced into Agrobacterium tumefaciens (strain EHA105) using the freeze-thaw method (Höfgen and Willmitzer, 1988). pCAMBIA1300 also was used for transformation and the resulting empty vector–transformed plants were used as controls. The Agrobacterium-mediated transformation was performed using vigorously growing calli derived from mature embryos of rice according to the method of Hiei et al. (1994) with slight modifications.
Abiotic treatments: Drought stress was induced by withholding water for up to 5 days. Two week old seedlings were grown in greenhouse conditions (28°C and a 16 h light/8 h dark cycle) and used in this experiment. The leaves began to wilt 3 days after the free water was removed. For salt stress, roots of 2 weeks old seedlings were immersed in 200 mM NaCl solution for up to 4 days. The time for returning stressed plants to the normal growth conditions was when approximately half of the control plants became wilted. The levels of drought, salt tolerance and wounding were evaluated based on the percentage of surviving seedlings after a period of recovery.
For the drought analysis, 40 seedlings were grown for harvesting of tissue for Northern analysis on root and leaf tissue. The leaves and root tissue of ten randomly selected plants were harvested separately for RNA extraction. The same sample size was applied to the salinity stress before RNA was extracted from these samples.
Biotic treatment: The fungal infection of 2 weeks old T1 transgenic plants was performed using the typical spray-inoculation method at a concentration of 106 spores mL-1(Lee et al., 2001). Blast resistance was evaluated based on fungal growth in plants (Qi and Yang, 2002) as well as lesion number and size. Samples were randomly selected for expression analysis in the T1 generation for each abiotic and biotic stress.
Southern and Northern analysis: Four micrograms of genomic DNA isolated from transgenic rice lines carrying OrMKK2. The DNA was then digested individually with EcoRI, HindIII, PstI and XbaI, fractionated on a 0.8% agarose gel and blotted onto a nylon membrane according to the standard protocol in Sambrook et al. (1989). Total RNA was isolated from OrMKK1 and OrMKK2 transgenic rice leaves using the Plant RNA Extraction (GibcoBRL, USA) kit. Fifteen micrograms of total RNA from each sample was separated on a 1.2% formaldehyde agarose gel and then transferred onto a nylon membrane. DNA and RNA ladders (Promega, USA) were added to the gel to estimate the sizes of hybridized bands. DNA and RNA gel blots were hybridized with a biotin–labeled gene-specific probe (3’ UTR of gene) in PerfectHyb buffer (Sigma, UK). Hybridization and washing conditions were based on the manufacturer’s instructions. A 300 bp sized probe of the 3’UTR region of OrMKK2 was used as a probe for both the analysis. A 250 bp sized probe of the 3’ UTR of OrMKK1 was used as a probe for the Northern blot analysis in biotic and abiotic stress response.
RESULTS AND DISCUSSION
Isolation and sequence analysis of OrMKK2: We previously identified a rice cDNA (OrMKK1) fragment that is inducible by the blast fungus and that shows high homology with plant MAPK genes. In this study, the corresponding full-length cDNA clone was isolated from a rice cDNA library using the OrMKK1 as a probe. One full length cDNA was selected, completely sequenced and analyzed. Once sequenced this gene was designated OrMKK2 as the sequence wasn’t identical to OrMKK1.
The OrMKK2 is 1381 bp long and has a 5’UTR that is 120 bp and a 3’UTR that is 195 bp and encodes a protein that is 356 amino acids (Fig. 1) with an estimated molecular mass of 53 kDa. The OrMKK2 protein contains all 11 subdomains that are conserved among all MAPK families (Hirt, 1997) and possesses a dual phosphorylation activation motif (TEY) located between subdomains VII and VIII (Fig. 2). Location of this domain is highlighted within the consensus sequence of the rice and Arabidopsis MAP kinases used in the alignment studies in this research. It shares the identical amino acid sequence encoded by OsMAPK5a, OsMAPK5b, OsMAPK2, OsMAPK1, OsMAPK3, OsMAPK6, AtMEK1, AtMAKK2 and AtMKK6 (Huang et al., 2000).
Phylogenetic analysis based on the sequence alignment of the catalytic domain suggest that OrMKK2 belongs to the A subgroup of the plant MAPK family (Fig. 3). Following are the clustering results of the rice sequences used in this study. Figure 3 shows the six subgroups that the sequences were divided into. Three of these (subgroups A, B and E) have been studied extensively and have been shown to be involved in biotic and abiotic stress (Kultz, 1998). The results of the analysis are as follows.
Group A : OrMKK2, OsMKK1, OsMKK6 and OsMAP1: The 4 different rice MAP kinases listed above fall under Group A. In earlier studies, the OsMAP1 (OsMPK5) gene was shown to be activated by ABA, drought and salt treatment(Wen et al., 2002). Xiong and Yang (2003) reported that OsMPK5 were inducible by abiotic (wounding, drought, salt and cold) stress. In their studies, Xiong and Yang (2003) also showed that OsMPK5 is a positive regulator of abiotic stress tolerance, but a negative regulator of the PR gene expression and broad-spectrum disease resistance. This indicates the possibility that OrMKK2 may also play a role in abiotic stress such as in OsMAP1.
Group B: OsMPK2, OsMPK6 and OrMKK1: This is one of the least studied group in rice MAPKs. OsMPK2 has been shown to be induced by an avirulent isolate of M. grisea (Reyna and Yang, 2006). However, not much is known about OsMPK6. OrMKK1 has been shown in the earlier study to be involved in biotic stress as concluded from pathogen infection and insect predation studies conducted on OrMKK1 lines.
Group E: OsMPK5a, OsMPK5b and OsMPK10: This group of genes are induced by both biotic and abiotic stress. Inducers such as BTH, dichloroisonicotinic acid, probenazole, JA, SA, ABA, ethylene and H2O2 were shown to be able to induce expression of these genes. In addition wounding and infection with M. grisea, salt treatment, drought stress and high/ low temperatures have resulted in the induction of these genes (Agrawal et al., 2002; Huang et al., 2002; Song and Goodman, 2002; Wen et al., 2002; Xiong and Yang, 2003; Reyna and Yang, 2006).
Copy number determinationof OrMKK2: The nucleotide sequence of OrMKK2 was analyzed with Webcutter2.0 to determine number of restriction sites for each of the above enzymes. Southern hybridization was performed with a probe covering the 3 region of OrMKK2. Two strongly hybridizing bands were observed when the genomic DNA was digested with EcoRI, PstI and XbaI (Fig. 4). This therefore indicates that there are two copies of OrMKK2 in the genome.
Northern analysis on OrMKK2: OrMKK1 and OrMKK2 lines were analyzed against biotic and abiotic stress.
 | |
| Fig. 1: | The coding region OrMKK2. Nucleotide and predicted amino acid sequences of the coding region provided with the start and stop codon indicated in bold. The 5’ and 3’ UTR are in italics |
This experiment was conducted to detect if both abiotic and biotic stress could induce the expression of both genes or were these genes regulated by different factors.
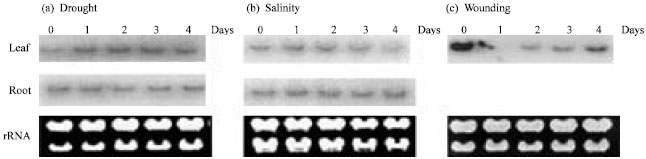
Figure 5a shows that the accumulation of OrMKK2 mRNA was observed in 2 weeks old seedlings that are drought stressed. In roots, drought responsive induction of OrMKK2 was detected within 0 day (12 h) of treatment and the level of expression was maintained until day 4 (Fig. 5a). However, the response of OrMKK2 was found to be much slower in shoots (Fig. 5a). The differential response is probably because roots desiccated prior to shoots in our experimental conditions.
In the drought and salt treatments, OrMKK2 was induced earlier in roots (within 1 day for drought and salinity) than in leaves (within 2 days for drought and salinity). However, under salt stress, thetranscripts declined after day 2 of treatment in the roots and remained constant in salt stressed leaves until day 3 and declined slightly on day 4 (Fig. 5b). The level of transcripts produced were higher in the roots compared to that observed in the leaves as the roots were the point of initial stress as in the case with drought tolerance (Fig. 5a).
Expression of OrMKK2 increased significantly in wounded leaves. The expression profiles peaked 30 min after wounding and then decreased rapidly to the basal
level expression (Fig. 5c). Similar profiles of immediate response and decline in response to wounding has been previously reported in plants (Nadarajah et al., 2000; Nadarajah and Ellis, 2000; Nadarajah, 2001; Nadarajah and Turner, 2003). The expression levels dropped after the initial response. This may indicate that the plant responds to the immediate compromise of its system and when there is no further threat to the system, the transcript levels are reduced.
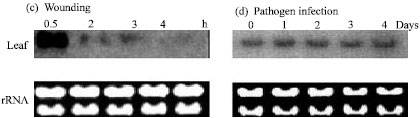
The OrMKK2 plantlets were also inoculated with spores from Magnaporthe grisea. The transcript levels in T1 plants were monitored over a period of 5 days. The level of expression over a 5 day period showed minimal expression levels (Fig. 5d).
However when the experiment was conducted using the OrMKK1 lines, the levels of gene expression were lower when subjected to abiotic stress. OrMKK1 was induced under biotic stress and also showed some form of expression during wounding (Fig. 6a-c). Fig. 6a shows the levels of expression triggered in OrMKK1 lines when subjected to drought stress. Unlike the response seen in Fig. 5a, where there was an elevated level of expression in the roots and some seen in the leaves, here we see that the expression profiles were near basal levels in both tissues (no difference in the roots and leaves).
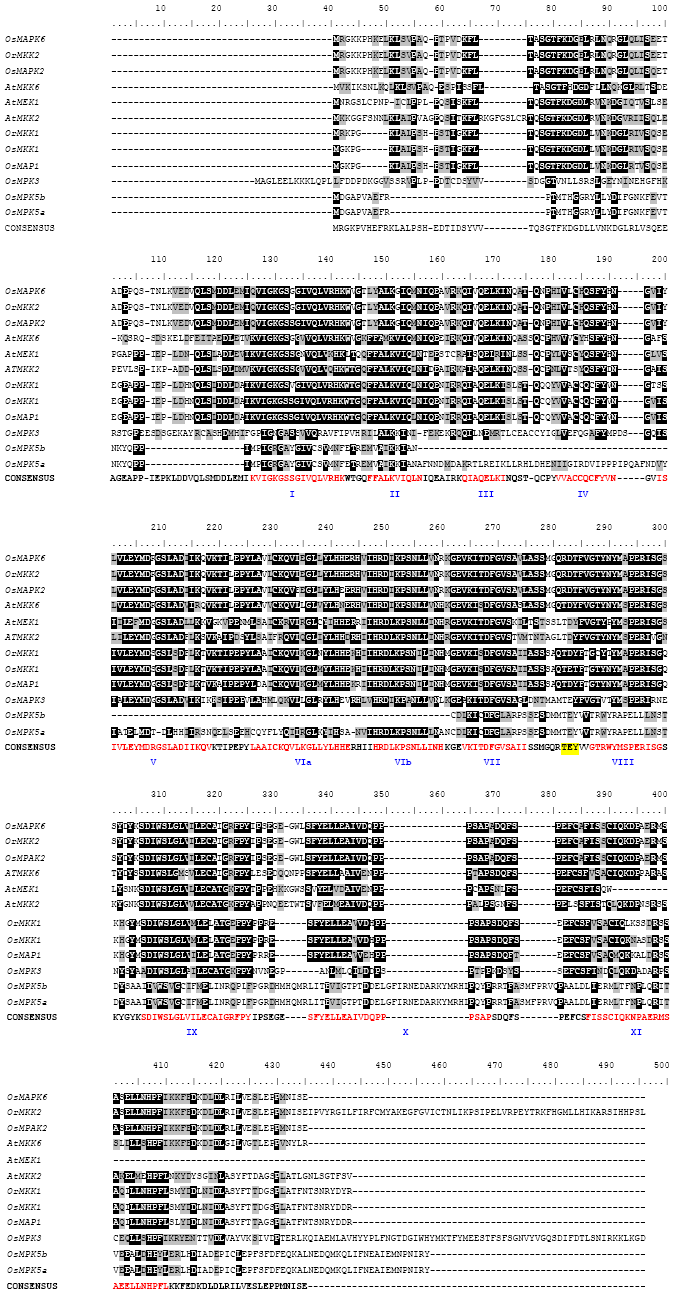 | |
| Fig. 2: | Comparison of OrMKK2 and representative MAPKs from rice and Arabidopsis. Conserved amino acid residues are shown in the consensus sequence. OrMKK1, ORMKK2, OsMKK1, OsMAP1, OsMAPK2, OsMAPK6, OsMPK3, OsMPK5a and OsMPK5b from rice (O. sativa), AtMEK1 and, AtMKK2 from Arabidopsis. The 11 subdomains of the protein kinases are indicated below the consensus sequences with Roman numerals. Thr (T) and Tyr (Y), two residues normally phosphorylated for the activation of MAPKs, are highlighted (location of TEY/TDY) in yellow |
Figure 6b which provides the northern assay results for salt stress also indicates that OrMKK1 has not been induced. There was however a little (basal) expression of the said gene in the leaf and root tissue. Wounding however seemed to induce the expression of OrMKK1 in the tested lines. The expression was also observed as in OrMKK2, 0.5 h post injury (Fig. 6c).
OrMKK2 positively regulates abiotic stress: Stress tolerance was evaluated based on the percentage of seedlings that survived after drought, salinity, wounding and pathogen infections. The OrMKK2 lines showed elevated levels of expressions when submitted to the abiotic stress over the experimental period. However, the response to pathogen infections was minimal. This suggests that the activation of OrMKK2 positively regulatesplant tolerance to abiotic stress such as drought, salinity and wounding but showed lower level of induction to fungal infections by M. grisea (Fig. 5a-d). The OrMKK1 gene however wasn’t induced by salt and drought stress but showed some level of induction in wounding (Fig. 6a-c), indicating involvement in biotic stress more then abiotic.
A complex signaling network that mediates the perception and responses to different environmental cues has evolved in plant systems over time. One such signaling network is the MAPK cascades which has remained conserved through evolution and is known to play a pivotal role in plant responses to multiple biotic and abiotic stress. Several MAPK genes have been identified and characterized functionally in plants where the importance of the three-kinase module (MAPKKK, MAPKK and MAPK) in signal transduction has been supported by numerous experimental studies by Ichimura et al. (2002). The complexity and role of the MAPK cascades in stress responses remains rather limited, considering the redundancy of cascade components, the antagonism among distinct cascades and the potential positive and negative regulation of different stress pathways by the same MAPK cascade (Tena et al., 2001).
Here, we have isolated a MAPK gene from rice and designated the cDNA as OrMKK2. Based on sequence similarity, OrMKK2 is closely related to tobacco WIPK and OsMAP1 in rice. Although, OrMKK2 is the ortholog of WIPK, its function may have evolved after the divergence of dicots and monocots. The alignment of OrMKK2 related sequences with OrMKK1 sequences show that there is a difference between both genes.
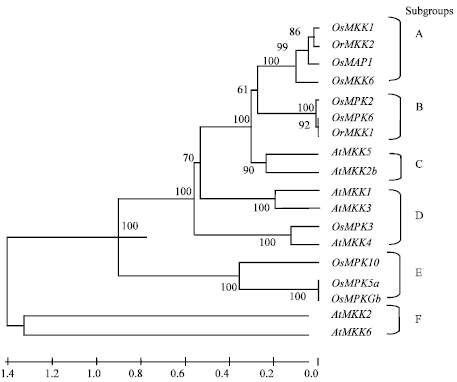 | |
| Fig. 3: | The phylogenetic tree was created using the Mega4 UPGMA programme with the Poisson correlation. The OrMKK2 sequences was analyzed with previously reported plant MAP kinases as follows: OrMKK1, OsMKK1, OsMAP1, OsMAPK2, OsMAPK6, OsMPK3, OsMPK5a, OsMPK5b and OsMKK10 from rice (O. sativa), AtMKK2b, AtMEK1, AtMKK2, AtMKK3, AtMKK4, AtMKK5 and AtMKK6 from Arabidopsis. Numbers on the left show the bootstrap value generated with 1000 replications |
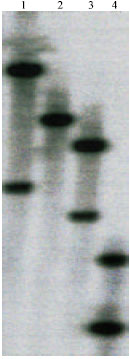 | |
| Fig. 4: | OrMKK2 copy numbers were identified via., Southern hybridization of cleaved genomic DNA. Fifteen microgram aliquots of genomic DNA were digested with 1: EcoRI, 2: HindIII, 3: PstI: and 4: XbaI, for OrMKK2 |
This may explain the difference in the response towards biotic and abiotic stress. However, as in all MAP kinases, these genes contain the 11 subdomains that are important in the regulation and function of the various MAP kinases.
OrMKK2 appears primarily to mediate abiotic stress, even though it also is activated by biotic stress, such as wounding-associated blast infection. The transgenic analysis show that OrMKK2 positively regulates abiotic stress tolerance and is also induced in wound associated pathogen infestations.
OrMKK1 gene was activated rapidly (1 day after infection) before the appearance of hypersensitive cell death, which normally occurs at 2 to 3 days after infection under the experimental conditions. The gene exhibited low levels of transcript production when under salt and drought stress. However when wounded, OrMKK1 was induced in the transgenic lines analyzed. There may be some similarity in the mode of gene activation in wounding and fungal infections, as fungi are known to produce infection pegs for penetration into plant cells. This we believe may trigger a similar response to wounding in plants (Hackett et al., 1998).
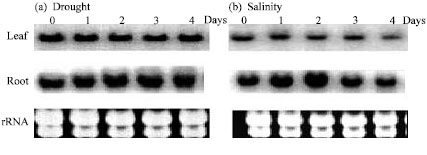 | |
| Fig. 5: | Northern blot analysis of OrMKK2 expression in 2 week old seedlings subjected to (a) drought (water withheld for up to 4 days), (b) salt (200 mM NaCl), or (c) wounding stress and (d) pathogen infections. For drought and salt stress, RNA from both root and leaf tissues was extracted at the times specified daily. Only leaf tissues were collected for wounding treatment. Tissue samples for wounding was harvested at 0.5 h and then at an hourly basis up to 4 h. The 3” UTR of OrMKK2 was used as probe |
 | |
| Fig. 6: | Northern blot analysis of OrMKK1 expression in 2 week old seedlings subjected to (a) drought (water withheld for up to 5 days), (b) salt (200 mM NaCl) and (c) wounding stress. For drought and salt stress, RNA from both root and leaf tissues was extracted at the times specified daily. Only leaf tissues were collected for wounding treatment. Tissue samples for wounding was harvested at 0.5 h and then at an hourly basis up to 4 h. The 3” UTR of OrMKK1 was used as probe |
There are a large number of plant MAPK genes that have been reported to be induced by abiotic stress, including drought, salinity, wounding and low temperatures (Jonak et al., 1996; Berberich et al., 1999; Munnik et al., 1999; Mikolajczyk et al., 2000; Agrawal et al., 2002; Huang et al., 2002). However, none of these genes has been characterized functionally to show positive regulation of abiotic stress by the MAPK cascades. The similarities seen between the expression profiles in wounding and blast fungus infections show that there may be some interlinked signaling and transduction involved in the regulation and expression of the MAPK genes. Since, there are a limited numbers of MAPK genes (e.g., 20 MAPKs, 10 MAPKKs and 60 MAPKKKs in the Arabidopsis genome), it would not be surprising if the plants have evolved an integrated signaling and transduction systems to delicately coordinate various physiological activities. It is therefore necessary for further research to be conducted to examine both the positive and negative effects of MAPK on different agronomic traits when modifying MAPK components for the genetic improvement of crop plants. By further understanding the MAPK cascades and carefully modifying the components, generation of new crop varieties that combine desirable traits such as enhanced disease resistance and abiotic stress tolerance may be produced.
ACKNOWLEDGMENTS
We would like to acknowledge the contribution of the Ministry of Science, Technology and Innovation for awarding a research grant (eScienceFund 02-01-02-SF0139) that enabled us to do the above reported research.
REFERENCES
- Hamel, L.P., M.C. Nicole S. Sritubtim M.J. Morency and M. Ellis et al., 2006. Ancient signals: Comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci., 11: 192-198.
PubMed - Hiei, Y., S. Ohta, T. Komari and T. Kumashiro, 1994. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J., 6: 271-282.
CrossRefDirect Link - Hofgen, R. and L. Willmitzer, 1988. Storage of competent cells for Agrobacterium transformation. Nucl. Acids Res., 16: 9877-9877.
PubMed - Huang, H.J., S.F. Fu, Y.H. Tai, W.C. Chou and D.D. Huang, 2002. Expression of Oryza sativa MAP kinase gene is developmentally regulated and stress-responsive. Physiol. Plant, 114: 572-580.
PubMed - Ichimura, K., K. Shinozaki, G. Tena J. Sheen and Y. Henry et al., 2002. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci., 7: 301-308.
CrossRef - Jin, H., M.J. Axtell, D. Dahlbeck, O. Ekwenna, S. Zhang, B. Staskawicz and B. Baker, 2002. NPK1, an MEKK1-like mitogen-activated protein kinase kinase kinase, regulates innate immunity and development in plants. Dev. Cell, 3: 291-297.
CrossRef - Jonak, C., S. Kiegerl, W. Ligterink, P.J. Barker, N.S. Huskisson and H. Hirt, 1996. Stress signaling in plants: A mitogen-activated protein kinase pathway is activated by cold and drought. Proc. Natl. Acad. Sci. USA., 93: 11274-11279.
Direct Link - Nadarajah, K. and J.G. Turner, 2003. The role of jasmonate in plant-pathogen interactions in Arabidopsis thaliana. J. Teknologi, 39: 9-16.
Direct Link - Nadarajah, K., 2001. The signalling pathway involved in plant-pathogen interactions in Arabidopsis thaliana. Pak. J. Biol. Sci., 4: 508-510.
CrossRefDirect Link - Qi, M. and Y. Yang, 2002. Quantification of Magnaporthe grisea during infection of rice plants using real-time PCR and northern blot/phosphoimaging analysis. Phytopathology, 92: 870-876.
CrossRef - Reyna, N.S. and Y. Yang, 2006. Molecular analysis of the rice MAP kinase gene family in relation to Magnaporthe grisea infection. Mol. Plant Microbe Interact., 19: 530-540.
CrossRef - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Tena, G., T. Asai, W.L. Chiu and J. Sheen, 2001. Plant mitogen activated protein kinase signaling cascades. Curr. Opin. Plant Biol., 4: 392-400.
PubMed - Wen, J.Q., K. Oono and R. Imai, 2002. Two novel mitogen-activated protein-signaling components, OsMEK1 and OsMAP1 are involved in a moderate low-temperature signaling pathway in rice. Plant Physiol., 129: 1880-1891.
PubMed - Xiong, L. and Y. Yang, 2003. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogen-activated protein kinase. Plant Cell, 15: 745-759.
CrossRefPubMedDirect Link - Xiong, L., M.W. Lee, M. Qi and Y. Yang, 2001. Identification of defense-related rice genes by suppression subtractive hybridization and differential screening. Mol. Plant Microbe Interact., 14: 685-692.
PubMed - Yang, K.Y., Y. Liu and S. Zhang, 2001. Activation of a mitogen-activated protein kinase pathway is involved in disease resistance in tobacco. Proc. Natl. Acad. Sci. USA., 98: 741-746.
PubMed - Zhang, S. and D.F. Klessig, 2001. MAPK cascades in plant defense signaling. Trends Plant Sci., 6: 520-527.
CrossRefPubMedDirect Link - Zhang, S. and Y. Liu, 2001. Activation of salicylic acid�induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco. Plant Cell, 13: 1877-1889.
PubMed - Nadarajah, K., N.M. Kasim and V.V. Fui, 2009. Function of mitogen-activated protein kinase gene in biotic stress. Asian J. Plant Sci., 85: 191-198.
CrossRefDirect Link - Mikolajczyk, M., O.S. Awotunde and G. Muszynska, 2000. Osmotic stress induces rapid activation of a salicylic acid-induced protein kinase and a homolog of protein kinase ASK1 in tobacco cell. Plant Cell, 12: 165-178.
PubMedDirect Link - Lee, M.W., M. Qi and Y. Yang, 2001. A novel jasmonic acid-inducible rice myb gene associates with fungal infection and host cell death. Mol. Plant-Microbe Interact., 14: 527-535.
PubMed