
Md. Mer Mosharraf Hossain
Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Kenji Kawai
Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Syunichirou Oshima
Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 5 | Page No.: 392-401







ABSTRACT
To find out adequate inactivator in substitution for the high concentration of formalin, different organic and inorganic chemicals, heat and chemicals combined with heat were tested for the inactivation of a fish-pathogenic bacterium Edwardsiella tarda strain. The survivality, sustainability of the cell surface antigenicity, bacterial protein and the total cell antigens of E. tarda were determined after treated with those inactivators. A major antigen at 37 kDa was detected by Sodium Dodecyl Sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) and Western blot analysis. Formalin (0.1%) with heat (70°Cx10 min) and 0.9% citric acid killed E. tarda resulted moderate killing activity with enough antigen-sustainability. This study suggests that formalin (0.1%) combined with heat (70°Cx10 min) and citric acid (0.9%) killed E. tarda as a new vaccine candidate.
PDF Abstract XML References Citation
How to cite this article
Md. Mer Mosharraf Hossain, Kenji Kawai and Syunichirou Oshima, 2009. Effective Inactivation of Edwardsiella tarda for the Development of Vaccine for Fish. Journal of Biological Sciences, 9: 392-401.
DOI: 10.3923/jbs.2009.392.401
URL: https://scialert.net/abstract/?doi=jbs.2009.392.401
DOI: 10.3923/jbs.2009.392.401
URL: https://scialert.net/abstract/?doi=jbs.2009.392.401
INTRODUCTION
Edwardsiella tarda is an enteric Gram-negative bacterium of the Enterobacteriaceae, first isolated from pond-cultured eel by Hoshina (1962). It is the causative agent of the systemic disease, edwardsiellosis, in many freshwater and marine fishes worldwide (Thune et al., 1993; Ling et al., 2000; Castro et al., 2008), Such as eel, Anguilla japonica (Wakabayashi and Egusa, 1973), chinook salmon, Oncorhynchus tshawytscha (Amandi et al., 1982), Japanese flounder, Paralichthys olivaceus (Nakatsugawa, 1983), tilapia, Tilapia niloticus (Kubota et al., 1981), carp, Cyprinus carpio (Sae-Oui et al., 1984), channel catfish, Ictalurus punctatus (Meyer and Bullock, 1973) striped bass, Morone saxatilis (Thune et al., 1993). E. tarda is widely distributed in nature, having been isolated from reptiles, birds, mammals (Van Damme and Vandepitte, 1984) including humans (Wilson and Waterer, 1989) and environmental water (Pitlik et al., 1987).
The use of synthetic chemicals and antibiotics (DePaola et al., 1995) for the control of fish disease may result with the emergence of antibiotic-resistant microbes, drug residues and environmental impacts (Aoki and Takahashi, 1987; Aoki et al., 1989). To limit the use of chemicals and antibiotics, vaccination is highly recommended (Chinabut and Puttinaowarat, 2005).
Over the last decade vaccination has become increasingly important for the prevention of infectious diseases in farmed marine and freshwater fish (Kwon et al., 2007; Carrias et al., 2008; Castro et al., 2008). Relatively very few research has been performed on the inactivation of E. tarda concerning to vaccine preparation. Most of the available vaccines for fish are inactivated with formalin. Several attempts made to induce protection by immunizing fish against edwardsiellosis with formalin-killed whole cells, cellular lipid and lipopolysaccharides (Bachmann et al., 1993; Inaba et al., 1973; Mussgay and Weiland, 1973; Salati et al., 1987; Huang et al., 2008; Dumrongphol et al., 2008), produced variable results.
Inactivation by heat in the preparation of inactivated vaccine (Ahmed et al., 1995; Farber and Brown, 1990) is not suitable for large-scale treatment as well as in fish culture (Mann et al., 2000) resulted heat resistance of pathogenic bacteria including harmless derivatives (Blatchley et al., 2001; Smith et al., 2000).
To produce microbicidal effects on Gram-negative or Gram-positive bacteria (Kabara et al., 1972; Kabara, 1978; Shibasaki ad Kato, 1978; Isaacs et al., 1995; Bergsson et al., 1998) with variety of lipids have been extensively studied in recent years.
The effective inactivator for the development of E. tarda vaccine for fish other than formalin has been receiving increasing attention. In this study, the effect of inactivation with chemicals, fatty acids, heat and chemicals combined with heat treatment were compared.
MATERIALS AND METHODS
Bacterial strains and growth conditions:This study was conducted at Fish Disease Laboratory, Department of Aquaculture, Faculty of Agriculture, Kochi University, Japan. From October 2006 to October 2008, E. tarda strain V-1, originally isolated from kidney of diseased eel (Anguilla japonica) in Japan, was pre-cultured for 24 h at 30°C in Brain Heart Infusion (BHI, Difco) broth and was inoculated into 1000 mL BHI broth, cultured with shaking at 30°C for 18 h followed by centrifugation at 4000 x g for 15 min at 4°C, harvested and stored at -80°C until used.
Inactivation by chemicals: Broth culture of the bacterium was centrifuged at 5,000 xg for 15 min, cells washed three times with PBS, resulting pellet suspended in Phosphate Buffered Saline (PBS) containing below 1% (from 0.1 to 0.9%) of formalin, ethanol (0.3 and 0.6%), Na2SO3 (0.3 and 0.6%), (NH4)2SO4 (0.4 and 0.7%), KCl (0.4, 0.6 and 0.9%) and amphotericin B (0.3 and 0.6%), incubated overnight at 15°C for 24 h. Excess chemicals was removed by three washes with PBS and stored at 4°C.
Inactivation by fatty acids: To achieve the treatment of fatty acids (0.1 to 0.9%), lauric acid (0.3, 0.6 and 0.9%), myristic acid (0.3, 0.6 and 0.9%), tannic acid (0.3, 0.6 and 0.9%) and citric acid (0.3, 0.6 and 0.9%) were used as inactivating fatty acids. Treated suspensions were incubated overnight at 15°C for 24 h.
Heat inactivation: The bacterial solution was heat at 60, 70, 80, 90 and 100°C for 10 min (Heating at 70°C for 10 min was also repeated). Treated solution was incubated overnight at 15°C for 24 h.
Inactivation by chemical combined with heat: Inactivation was performed with formalin with 0.05, 0.1% at 60 and 70°C for 10 min (0.05, 0.1% Phenol at 60 and 70°C for 10 min was repeated) incubated overnight at 15°C for 24 h. Excess chemicals was removed by three washes with PBS and stored at 4°C.
Viable counts (confirmation of bacterial inactivation): The number of cells surviving chemicals, fatty acids, heat and chemicals combined with heat treatments was estimated from colony counts (Table 1). The drop plate counting method was employed (Reed and Reed, 1948). The viable count was also calculated from at least two different dilutions, with weighting for dilution (Farmiloe et al., 1954).
| Table 1: | Agglutination titers of Edwardsiella tarda V-1 strain inactivated with chemicals and fatty acids |
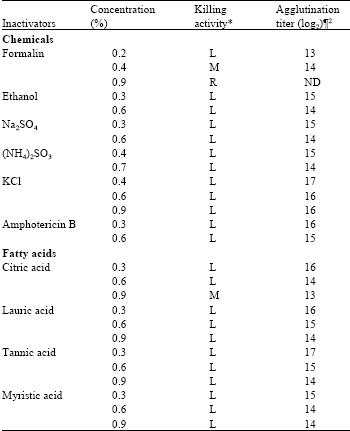 | |
| *L: Low killing where bacteria still remain alive after 24 h, M: Moderate killing in 24 h, where almost all bacteria were killed within 24 h, R: Rapid killing in 6 or 12 h. ¶ Values indicate the highest dilution times of the serum with positive agglutination. 2Geometric mean reciprocal log2 value of the highest dilution of the serum that showed positive agglutination. ND: Agglutination value was not determined by autoagglutination | |
Agglutination: The inactivated E. tarda suspension was adjusted to an OD of 1.0 at 550 nm and 30 μL of the suspension (McFarland No. 2) was added to serum dilutions. Rabbit anti-E. tarda whole-cell antiserum served as a positive control and PBS served as a negative control. Plates were incubated 2 h at 25°C and overnight at 4°C and the agglutination was scored (Table 1).
SDS-PAGE and Western blots: Protein electrophoresis was performed by using the method of Laemmli (1970). The proteins were electrophoretically transferred to nitrocellulose paper (0.45 μm pore size, Bio-Rad) by using a semi-dry apparatus (Bio- Rad) as described by Towbin et al. (1979) after blocking with 1% skim milk at 4°C overnight, the membrane was reacted with rabbit anti-E. tarda V-1 serum.
Statistical analysis: Data were analyzed by the student’s t-test/ANOVA. Tukey’s test was used to compare the mean values (Zar, 1984) using the StatPlus 2007 Professional.
RESULTS
Effect of chemical inactivation: Compared to different treatment of E. tarda with formalin, only the treatment formalin (0.4%) that showed moderate killing activity in Fig. 1 and Table 1. Inactivation profile of E. tarda with different chemicals (Fig. 2, 3, Table 1), ethanol (0.3, 0.6%), Na2SO3 (0.3, 0.6%), (NH4)2SO4 (0.4, 0.7%), KCl (0.4, 0.6, 0.9%) and amphotericin B (0.3, 0.6%) showed low killing activity, where the treated bacteria still remained alive after 24 h.
Effect of fatty acids inactivation: Compared to fatty acids killed E. tarda vaccine (Fig. 4), citric acid 0.9% only the treatment that showed moderate killing activity. The
bacteria still remained alive after 24 h with expressing low killing activity by the treatment of lauric acid (0.3, 0.6, 0.9%), myristic acid (0.3, 0.6, 0.9%), tannic acid (0.3, 0.6%, 0.9%) and citric acid (0.3, 0.6%).
Effect of heat and chemicals combined with heat inactivation: The bacterium was completely inactivated by 6 to 12 h with heating at 80, 90 and 100°C for 15 min. Bacterium still remains alive after 24 h heating at 60 and 70°C for 15 min, 0.05% formalin with heating at 60 and 70°C for 10 min, 0.1% formalin with heating at 60°C for 10 min and 0.05 and 0.1% phenol with heating at 60 and 70°C for 10 min. Compared to different bactrins of E. tarda only the treatment 0.1% formalin with heating at 70°C for 10 min showed moderate killing activity in 24 h (Fig. 5).
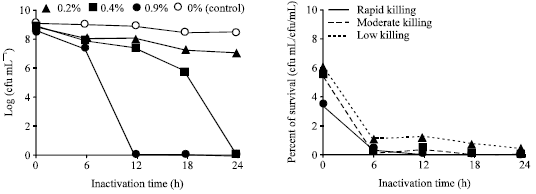 | |
| Fig. 1: | Inactivation and survival of Edwarsiella tarda V-1 strain after incubation with different concentration of formalin |
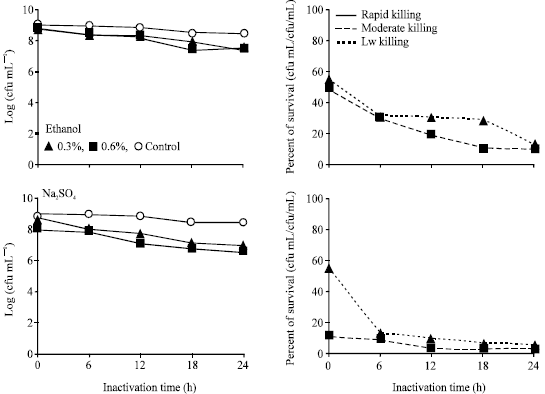 | |
| Fig. 2: | Inactivation profile and survival of Edwarsiella tarda V-1 strain after incubation with different concentration of chemicals |
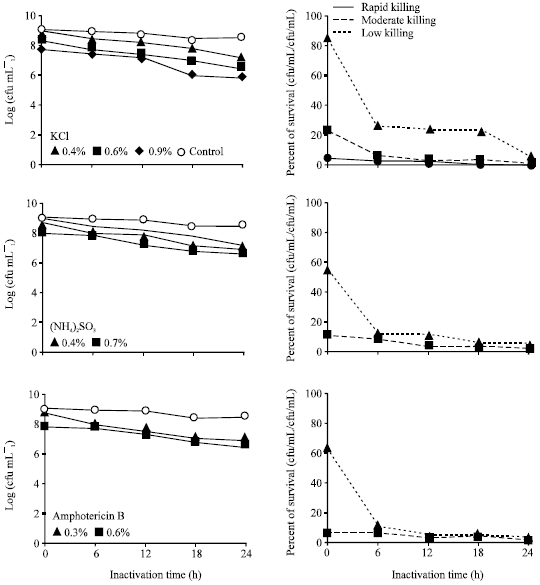 | |
| Fig. 3: | Inactivation profile and survival of Edwarsiella tarda V-1 strain after incubation with different concentration of chemicals |
Antigenicity of inactivated E. tarda measured by agglutination reaction
Inactivation by chemicals: The agglutination titers of different bactrins of E. tarda against rabbit antiserum were shown in Table 1. Higher percentage of formalin (0.9%) rose autoagglutination which hid the agglutination titer. Moderately high agglutination titer was observed with formalin (0.4%).
Inactivation by fatty acid: The agglutination titers with E. tarda cells inactivated by lauric acid, myristic acid, tannic acid and citric acid against rabbit antiserum are shown in Table 1. The 0.9% citric acid brought moderately high agglutination titer.
Inactivation by heating and chemicals combined with heating: Compared to different bactrins of E. tarda, only the treatment 0.1% formalin with 70°C for 10 min that showed moderately high agglutination titer in Table 2. The agglutination titer was hid by autoagglutination with treatment at 80, 90 and 100°C for 15 min, where the treatment with 60 and 70°C for 15 min, 0.05% formalin with 60 and 70°C and 0.1% formalin with 60°C for 10 min; 0.05 and 0.1% phenol with 60 and 70°C for 10 min brought higher agglutination titer in Table 2.
Antigenicity of inactivated E. tarda measured by Western blot: The profiles of the total proteins (SDS-PAGE) and major protective antigen of this bacterium at 37 kDa location of E. tarda V-1 strain was strongly reacted with rabbit antiserum against citric acid (0.9%) and formalin (0.1%) with heat (70°C for 10 min) killed cells in Fig. 6. At 97, 75, 74, 45, 43 and 37 kDa locations were reacted with rabbit antiserum against formalin (0.4, 0.9%), myristic acid (0.3, 06, 0.9%), citric acid (0.3, 0.6%) and formalin (0.05%) with heat (60°C for 10 min) killed E. tarda in Fig. 6.
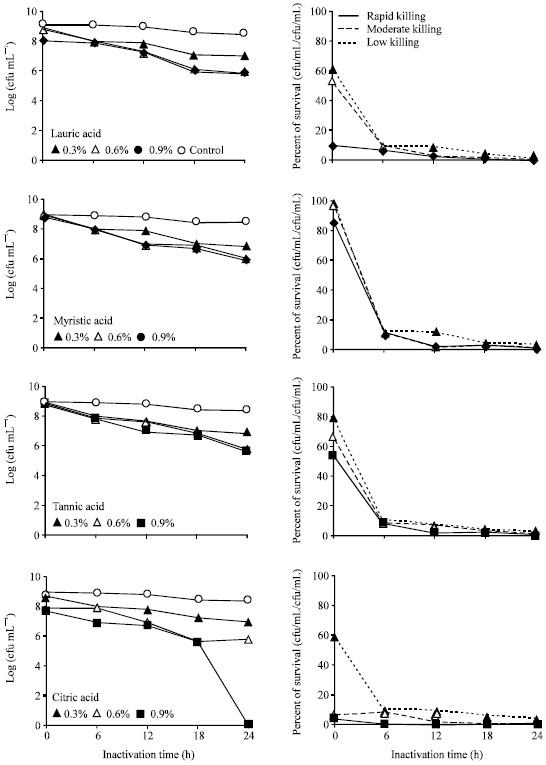 | |
| Fig. 4: | Inactivation profile and survival of Edwardsiella tarda V-1 strain after incubation with different concentration of fatty acids |
DISCUSSION
E. tarda is a pathogen of aquaculture industry which can lead to serious diseases. Present new approach to produce an inactivated E. tarda vaccine considering awareness of residual effect to human is based on the chemicals fatty acids, heat, low concentration of formalin combined with heat other than formalin inactivation of cells with high concentration which has been extensively investigated in this study.
In the present study, the high levels of inactivating activity (Bergsson et al., 1998; Lampe et al., 1997) showed high concentrations of citric acid that had not been seen with fatty acids.
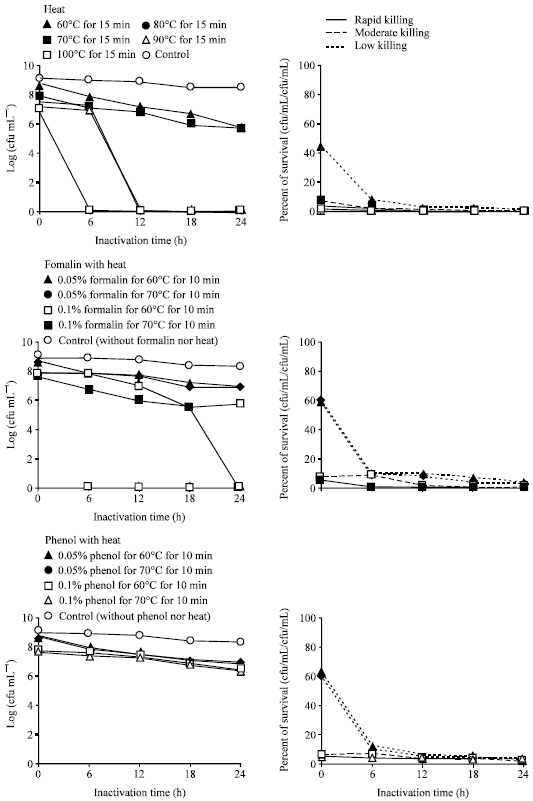 | |
| Fig. 5: | Inactivation profile and survival of Edwarsiella tarda V-1 strain after incubation with heat and chemicals combined with heat |
We therefore hypothesized that citric acid kills the bacterium by affecting outer membrane by citric acid, leading to disruption the elementary bodies of the cell membrane and remaining the bacterial cell membrane intact (Bergsson et al., 1998; Lampe et al., 1997).
Significantly elevated agglutination titer and resistance against E. tarda with formalin killed cells were found by Swain et al. (2007), which was similar to the treatment with low concentration of formalin combined with heat. E. tarda, strains positively responded to heating at 60°C for 30 min and it was rapidly destroyed by heating at 100°C for 15 min (Bockemühl et al., 1983).
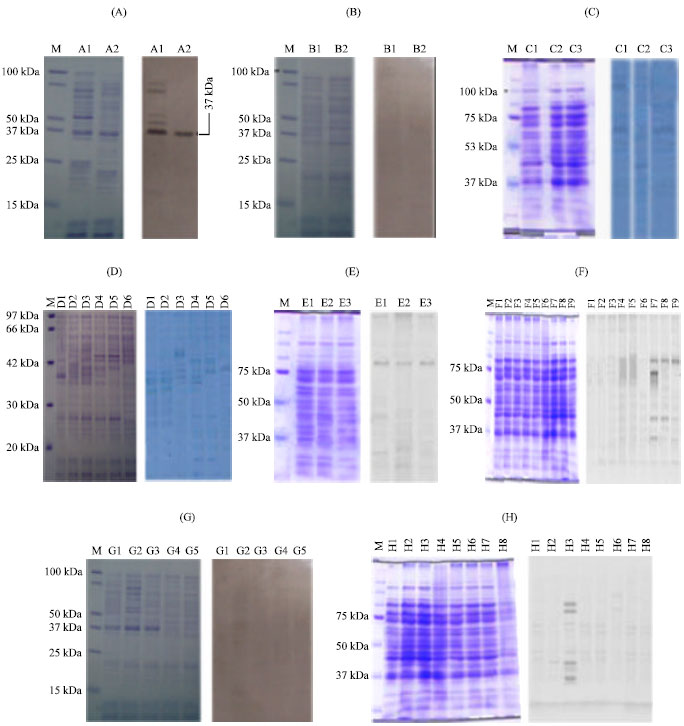 | |
| Fig. 6: | SDS-PAGE and Western blotting profiles of chemicals, fatty acid, heat and chemicals combined with heat inactivated Edwardsiella tarda V-1 strain. Left, SDS-PAGE; Right, Western blot. M: Molecular weight marker; A1, 0.4% formalin; A2, 0.9% formalin; B1, 0.3% ethanol; B2, 0.6% ethanol; C1, 0.4% KCl; C2, 0.6% KCl; C3, 0.9% KCl; D1, 0.3% NaSO4; D2, 0.6% NaSO4; D3, 0.4% (NH4)2SO4; D4, 0.7% (NH4)2SO4; D5, 0.3%amphotericin B; D6, 0.6% amphotericin B; E1, 0.3% myristic acid; E2, 0.6% myristic acid; E3, 0.9% myristic acid; F1, 0.9% tannic acid; F2, 0.6% tannic acid; F3, 0.3% tannic acid; F4, 0.9% lauric acid; F5, 0.6% lauric acid; F6, 0.3% lauric acid; F7, 0.9% citric acid; F8, 0.6% citric acid; F9, 0.3% citric acid; G1, heat at 60°C for 15 min; G2, heat at 70°C for 15 min; G3, heat at 80°C for 15 min; G4, heat at 90°C for 15 min; G5, heat at 100°C for 15 min; H1, 0.05% formalin with heat at 70°C for 10 min; H2, 0.05% formalin with heat at 60°C for 10 min; H3, 0.1% formalin with heat at 70°C for 10 min; H4, 0.1% formalin with heat at 60°C for 10 min; H5, 0.05% phenol with heat at 70°C for 10 min; H6, 0.05% phenol with heat at 60°C for 10 min; H7, 0.1% phenol with heat at 70°C for 10 min and H8, 0.1% phenol with heat at 60°C for 10 min |
| Table 2: | Agglutination titers of Edwardsiella tarda V-1 strain inactivated by heat and chemicals combined with heat |
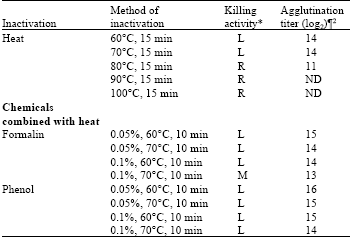 | |
| *L: Low killing where bacteria still remain alive after 24 h, M: Moderate killing in 24 h, where almost all bacteria were killed within 24 h, R: Rapid killing in 6 or 12 h. ¶ Values indicate the highest dilution times of the serum with positive agglutination. 2Geometric mean reciprocal log2 value of the highest dilution of the serum that showed positive agglutination. ND: Agglutination value was not determined by autoagglutination. | |
Compared to different bactrins of E. tarda (Salati and Kusuda, 1985; Salati et al., 1987; Kwon et al., 2007; Swain et al., 2007), only the treatment citric acid (0.9%) and formalin (0.1%) with heat (70°C for 10 min) killed E. tarda that showed enough-sustainability and expressing moderate killing activity where the major protective antigen of this bacterium at 37 kDa location (Tu and Kawai, 1999) was strongly reacted with rabbit antiserum against E. tarda.
Over the last decade effective inactivation and antigenic potency of the bacterin after inactivation was not properly evaluated with E. tarda. Compared to heat or formalin killed bacterial vaccines (Gutierrez and Miyazaki, 1994; Crosbie and Nowak, 2004; Castro et al., 2008), in present study, citric acid and low concentration of formalin combined with heat could be the potential tool for E. tarda inactivation. Also, suggested as an alternate approach for the inactivation of bacteria (Nakayama et al., 1996; Sonoike, 1997) which have frequently caused the reduction of antigenicity, residual effect (Smith et al., 1956).
Exactly why the low concentration of formalin with heat at 70°C for 10 min and 0.9% citric acids induced effective inactivation is remained unclear. The quantity of the protective antigens 37 kDa in this preparation is higher, but another possibility is that the method of production destroyed less of the protective epitopes than bacterins of whole cells. A further influencing factor might be differences in particle structure between the vaccines (Taisuke et al., 2003). We can only speculate about the structure of the PKC vaccine; however another study showed that the antigen form can influence immunogenicity (Schirmbeck et al., 1995).
In conclusion, as this promising type of an inactivation bacterin preparation seems to be favorable over conventional vaccines, we suggested formalin (0.1%) with heat (70°C for 10 min) and citric acid (0.9%) killed E. tarda as a new vaccine candidate.
ACKNOWLEDGMENT
We would like to thank the staff and students of Fish Disease Laboratory, Faculty of Agriculture, Kochi University for their help in the field.
REFERENCES
- Bergsson, G., J. Arnfinnsson, S.M. Karlsson, O. Steingrimsson and H. Thormar, 1998. In vitro inactivation of Chlamydia trachomatis by fatty acids and monoglycerides. Antimicrob. Agents Chemother., 42: 2290-2294.
PubMedDirect Link - Blatchley, E.R., N. Dumoutier, T.N. Halaby, Y. Levi and J.M. Laine, 2001. Bacterial responses to ultraviolet irradiation. Water Sci. Technol., 43: 179-186.
Direct Link - Castro, N., A.E. Toranzo, S. Nunez and B. Magarinos, 2008. Development of an effective Edwardsiella tarda vaccine for cultured turbot (Scophthalmus maximus). Fish Shellfish Immunol., 25: 208-212.
Direct Link - Chinabut, S. and S. Puttinaowarat, 2005. The choice of disease control strategies to secure international market access for aquaculture products. Dev. Biol., 121: 255-261.
Direct Link - Crosbie, P.B.B. and B.F. Nowak, 2004. Immune responses of barramundi, Lates calcarifer (Bloch), after administration of an experimental Vibrio harveyi bacterin by intraperitoneal injection, anal intubation and immersion. J. Fish Dis., 27: 623-632.
Direct Link - DePaola, A., J.T. Peeler and G.E. Rodrick, 1995. Effect of oxytetracycline-medicated feed on antibiotic resistance of gram-negative bacteria in catfish ponds. Applied Environ. Microbiol., 61: 2335-2340.
PubMedDirect Link - Farber, J.M. and B.E. Brown, 1990. Effect of prior heat shock on heat resistance of Listeria monocytogenes in meat. Applied Environ. Microbiol., 56: 1584-1587.
Direct Link - Farmiloe, F.J., S.J. Cornford, J.B.M. Coppock and M. Ingram, 1954. The Survival of Bacillus subtilis spores in the baking of bread. J. Sci. Food Agric., 5: 292-304.
CrossRefDirect Link - Gutierrez, M.A. and T. Miyazaki, 1994. Responses of Japanese eels to oral challenge with Edwardsiella tarda after vaccination with formalin-killed cells or lipopolysaccharide of the bacterium. J. Aquat. Anim. Health, 6: 110-117.
CrossRefDirect Link - Inaba, Y., H. Kurogi, K. Sato, Y. Goto, T. Omori and M. Matumoto, 1973. Formalin-inactivated, aluminum phosphate gel-adsorbed vaccine of bovine ephemeral fever virus. Arch. Virol., 42: 42-53.
CrossRefDirect Link - Isaacs, C.E., R.E. Litov and H. Thormar, 1995. Antimicrobial activity of lipids added to human milk, infant formula and bovine milk. Nutr. Biochem., 6: 362-366.
CrossRefDirect Link - Kabara, J.J., D.M. Swieczkowski, A.J. Conley and J.P. Truant, 1972. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother., 2: 23-28.
Direct Link - Kwon, S.R., E.H. Lee, Y.K. Nam, S.K. Kim and K.H. Kim, 2007. Efficacy of oral immunization with Edwardsiella tarda ghosts against edwardsiellosis in olive flounder (Paralichthys olivaceus). Aquaculture, 269: 84-88.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Mann, A., M. Knifer and H. Leuenberger, 2000. Thermal sterilization of heat-sensitive products using high-temperature short-time sterilization. J. Pharm. Sci., 90: 275-287.
Direct Link - Meyer, F.P. and G.L. Bullock, 1973. Edwardsiella tarda, a new pathogen of channel catfish (Ictalurus punctatus). Applied Microbiol., 25: 155-156.
Direct Link - Mussgay, M. and E. Weiland, 1973. Preparation of inactivated vaccines against Alphaviruses using Semliki forest virus-white mouse as a model. I. Inactivation experiments and evaluation of double inactivated subunit vaccines. Int. Virol., 1: 259-268.
CrossRefDirect Link - Nakatsugawa, T., 1983. Edwardsiella tarda isolated from cultured young flounder. Fish Pathol., 18: 99-101.
Direct Link - Nakayama, A., Y. Yano, S. Kobayashi, M. Ishikawa and K. Sakai, 1996. Comparison of pressure resistances of spores of six Bacillus strains with their heat resistances. Applied Environ. Microbiol., 62: 3897-3900.
Direct Link - Pitlik, S., S.A. Berger and D. Huminer, 1987. Nonenteric infections acquired through contact with water. Rev. Infect. Dis., 9: 54-63.
CrossRefDirect Link - Reed, R.W. and G.B. Reed, 1948. Drop plate method of counting viable bacteria. Can. J. Res., 26: 317-326.
CrossRefDirect Link - Sae-Oui, D., K. Muroga and T. Nakai, 1984. A case of Edwardsiella tarda infection in cultured colored carp Cyprinus carpio. Fish Pathol., 19: 197-199.
Direct Link - Salati, F. and R. Kusuda, 1985. Vaccine preparations used for immunization of eel Anguilla japonica against Edwardsiella tarda infection. Nipp. Sui. Gakka., 51: 1233-1237.
CrossRefDirect Link - Schirmbeck, R., W. Bohm, K. Melber and J. Reimann, 1995. Processing of exogenous heat-aggregated (denatured) and particulate (native) hepatitis B surface antigen for class I-restricted epitope presentation. J. Immunol., 155: 4676-4684.
Direct Link - Smith, R.J., S.C. Kehoe, K.G. Mcguigan and M.R. Barer, 2000. Effect of simulated solar disinfection of water on infectivity of Salmonella typhimurium. Lett. Applied Microbiol., 31: 284-288.
Direct Link - Swain, P., A. Behura, S. Dash and S.K. Nayak, 2007. Serum antibody response of Indian major carp, Labeo rohita to three species of pathogenic bacteria; Aeromonas hydrophila, Edwardsiella tarda and Pseudomonas fluorescens. Vet. Immunol. Immunopathol., 117: 137-141.
Direct Link - Taisuke, W., F. Soichi, H. Junichi, K. Tetsuya, O. Hirokazu and Y. Makari, 2003. Inactivation of geobacillus stearothermophilus spores by high-pressure carbon dioxide treatment. Applied Environ. Microbiol., 69: 7124-7129.
CrossRefDirect Link - Thune, R.L., L.A. Stanley and R.K. Cooper, 1993. Pathogenesis of gram-negative bacterial infections in warmwater fish. Annu. Rev. Fish. Dis., 3: 37-68.
CrossRefDirect Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Van Damme, L.R. and J. Vandepitte, 1984. Isolation of Edwardsiella tarda and Plesiomonas shigelloides from mammals and birds in Zaire. Rev. Elev. Med. Vet. Pays Trop., 37: 145-151.
Direct Link - Wilson, J. and R. Waterer, 1989. Serious infections with Edwardsiella tarda: A case report and review of the literature. Arch. Int. Med., 149: 208-210.
CrossRefDirect Link - Carrias, A.A., J.S. Terhune, C.A. Sayles and J.A. Chappell, 2008. Effects of an extended hatchery phase and vaccination against enteric septicemia of catfish on the production of channel catfish, Ictalurus punctatus, fingerlings. J. World Aqua. Soc., 39: 259-266.
CrossRefDirect Link - Dumrongphol, Y., T. Hirota, H. Kondo, T. Aoki and I. Hirono, 2008. Identification of novel genes in Japanese flounder (Paralichthys olivaceus) head kidney up-regulated after vaccination with Streptococcus iniae formalin-killed cells. Fish Shellfish Immunol., 26: 197-200.
CrossRef - Huang, Y., H. Lou, X. Wu and Y. Chen, 2008. Characterization of the BPI-like gene from a subtracted cDNA library of large yellow croaker (Pseudosciaena crocea) and induced expression by formalin-inactivated Vibrio alginolyticus and Nocardia seriolae vaccine challenges. Fish Shellfish Immunol., 25: 740-750.
PubMed - Kawai, K., Y. Liu, K. Ohnishi and S.I. Oshima, 2004. A conserved 37 kDa outer membrane protein of Edwardsiella tarda is an effective vaccine candidate. Vaccine, 22: 3411-3418.
CrossRefDirect Link