
Y. Fujii
Laboratory of Marine Biochemistry, Department of Environmental Biosciences, International Graduate School of Arts and Sciences, Yokohama City University, 22-2 Seto, Kanazawa-Ku, Yokohama 236-0027, Japan
S.M.A. Kawsar
Laboratory of Marine Biochemistry, Department of Environmental Biosciences, International Graduate School of Arts and Sciences, Yokohama City University, 22-2 Seto, Kanazawa-Ku, Yokohama 236-0027, Japan
R. Matsumoto
Laboratory of Marine Biochemistry, Department of Environmental Biosciences, International Graduate School of Arts and Sciences, Yokohama City University, 22-2 Seto, Kanazawa-Ku, Yokohama 236-0027, Japan
H. Yasumitsu
Laboratory of Marine Biochemistry, Department of Environmental Biosciences, International Graduate School of Arts and Sciences, Yokohama City University, 22-2 Seto, Kanazawa-Ku, Yokohama 236-0027, Japan
N. Kojima
Laboratory of Environmental Biosciences, YSFH, 6 Ono, Tsurumi-Ku, Yokohama 230-0046, Japan
Y. Ozeki
Laboratory of Marine Biochemistry, Department of Environmental Biosciences, International Graduate School of Arts and Sciences, Yokohama City University, 22-2 Seto, Kanazawa-Ku, Yokohama 236-0027, Japan
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 4 | Page No.: 319-325







ABSTRACT
To find novel carbohydrate-binding proteins (lectins) from marine invertebrates to understand the binding mechanism of the protein and to apply it for glycan-dependent diagnostics and/or glycoconjugates capture technology. A D-galactoside-binding lectin was purified from foot of bladder moon shell, Glossaulax didyma by lactosyl-agarose affinity chromatography. The crude supernatant by Tris-buffered saline had strong hemagglutination activity against trypsinized and glutaraldehyde-fixed human erythrocyte. However, the activity was not inhibited by any tested saccharides and chilete reagents. On the other hand, the dialyzed crude supernatant obtained from the precipitates with 100 mM lactose in Tris-buffered saline had also hemagglutination activity inhibited by β-galactoside and D-galactose. The lectin was purified with lactosyl-agarose affinity chromatography. The molecular mass of the lectin was determined to be 60 kDa by SDS-PAGE under reducing and non-reducing conditions and being a 60 kDa polypeptide monomer by gel permeation chromatography. The association-rate constant (kass) and dissociation-rate constant (kdiss) determined for the lectin against asialofetuin was determined as 5.4x104 M-1sec-1 and 7.2x10-3sec-1, respectively. Lectin-conjugated Sepharose gel captured asialofetuin and eluted it by lactose-containing buffer from the gel, indicating that the lectin could catch the asialoglycoprotein. It was concluded that a many amount of a D-galactoside-binding lectin which can catch asialoglycoprotein presents in foot of the bladder moon shell.
PDF Abstract XML References Citation
How to cite this article
Y. Fujii, S.M.A. Kawsar, R. Matsumoto, H. Yasumitsu, N. Kojima and Y. Ozeki, 2009. Purification and Characterization of a D-Galactoside-Binding Lectin Purified from Bladder Moon Shell (Glossaulax didyma Roding). Journal of Biological Sciences, 9: 319-325.
DOI: 10.3923/jbs.2009.319.325
URL: https://scialert.net/abstract/?doi=jbs.2009.319.325
DOI: 10.3923/jbs.2009.319.325
URL: https://scialert.net/abstract/?doi=jbs.2009.319.325
INTRODUCTION
Lectins are carbohydrate-binding proteins recognizing specific oligosaccharide structures and they are widely distributed in many organisms. Marine invertebrates locating at lower order in animal kingdom and have not acquired immunity to protect themselves from enemies develop lectins or toxins inside of their organs. Lectin believes to function such as innate immunity, cellular attachment, cellular proliferating regulation and molecular glue (Ebina et al., 1995). Mollusk is one of big phylum and it lives around the world from in aqua to on land with many morphological diversities. Many kind of lectin have been isolated from Mollusk and several new lectins are being purified from them in recent (Yamaura et al., 2008; Takahashi et al., 2008; Gourdine et al., 2007; Naganuma et al., 2006; Sanchez et al., 2006). They are well adapting to their living environment, however there are also many parasitic microorganisms or other predatory species around them to make damage. Sometimes it influences for our ecosystems and economies. In Manila clam, the expressional level of the lectin purified from the species has been found to change according to the levels of infection by parasites (Kim et al., 2006). Besides, the mollusk lectin regulated the cell proliferation activity (Banerjee et al., 2004), suggesting that potentially lectins act for body protection of host cells. Gastropoda, consisting of class conchs, whelks, nudibranch, snails and slugs is the biggest class in mollusk, holding over 70% species. Bladder moon shells, Glossaulax didyma has shell to 3 cm high and is commonly living in sand on lower shore at the Pacific, Atlantic and Indian oceans. The foot is thick and broad. It is folded upwards around the sides of the shell. Bladder moon shells are carnivores, eating clamshell by the penetration of hole by dissolving the shells with sulfuric acid produced from their metabolites. In Japan, they are found under sand of many coasts. As morphological feature, bladder moon shells have big mantle covering shell and spawning eggs characteristic shaped teacup on coast of sand. Physiologically, it has many mucus and a few traits have appeared. Some reports showed that they contain in the body a few amount of tetrodotoxin seemed to be derived from infected microorganisms (Hwang et al., 1991). As this way, the bladder moon shell is harmful for other edible shells. However, the use of this biomass for research tool is valuable, because it can take much amount easily. From the view, we screened the presence of new lectin from the species and characterized the biochemical properties to find the possibility as the applicable tool for the study of glycobiology.
MATERIALS AND METHODS
Animal: Bladder moon shells were collected from the beneath of sand flat in tidal beach at both coasts of the Futtsu coast, Chiba Prefecture and the Misaki, Kanagawa Prefecture, Japan from June to July in 2008. Animals were stored at -80°C or used after collection according to the situation.
Chemicals: Lactose, sucrose, melibiose, D-glucose, D-galactose, D-mannose, D-fucose, L-fucose, N-acetyl D-galactosamine and N-acetyl D-glucosamine were purchased from Wako Pure Chemical Ind. Ltd., Japan and were of the highest purity grade. Lactosyl-agarose was purchased from Seikagaku Kogyo Co. Ltd., Japan. Methyl α-D-galactopyranoside and methyl β-D-galactopyranosides were from Pfanstiehl Laboratories Inc., IL, USA. Glycoproteins, asialofetuin and fetuin were purchased from Sigma, USA. Sephadex G-75, N-hydroxysuccimide (NHS)-activated Sepharose 4 Fast flow, Superdex 75 column, Sensor chip CM5 and Amine coupling kit (including 1-ethyl-3-(3-dimethylamino-propyl) carbodiimide (EDC), NHS and 1 M ethanolamine-HCl, pH 8.5) were from GE Healthcare, USA. Bicinchoninic acid (BCA) kit was purchased from Pierce Co. Ltd., USA.
Purification of lectin: Two hundred grams (wet weight) of bladder moon shells were disrupted to paste with a razor blade and homogenized with 10 volumes (w/v) of Tris-buffered saline (TBS) (10 mM Tris(hydroxylmethyl aminomethane)-HCl, pH 7.4, 150 mM NaCl) containing 2 mM of each β-mercaptoethanol and a protease inhibitor mixture. The homogenates were centrifuged at 14,720 g in 500 mL centrifuge bottles for 1 h at 4°C with a Suprema 21 centrifuge equipped with an NA-18HS rotor (TOMY Co. Ltd., Japan). The supernatant was centrifuged again at 27,500 g for 1 h at 4°C and the precipitate was homogenated again with 3 volumes (w/v) of TBES (TBS containing 10 mM EDTA) containing 50 mM lactose and 2 mM β-mercaptoethanol over night at 4°C. It was centrifuged at 27,500 g for 1 h at 4°C and the supernatant was dialyzed against TBS till free galactose has left. Centrifuged crude extract was applied to an affinity column of lactosyl-agarose (10 mL) being fitted with a Sephadex G-75 pre-column (5 mL). After applying the extracts, the lactosyl-agarose column was extensively washed with TBS containing 2 mM β-merocaptoethanol. The lectin was eluted from the column with 50 mM lactose in TBS and each 1 mL of elution was collected in tubes with a fraction collector. Each chromatography step during washing and elution was monitored using a UV monitor (ATTO Co. Ltd., Japan) by the measurement of the absorbance at 280 nm. The eluted fractions were measured by UV spectrophotometer on the absorbance at 280 nm and SDS-PAGE (Laemmli, 1970) described as below were combined and dialyzed against 1,000 times volumes of TBS to remove free lactose. The protein concentration of crude extract and purified lectin were determined by the BCA protein assay kit with bovine serum albumin as the standard protein (Smith et al., 1985; Wiechelman et al., 1988) by measuring absorbance at 562 nm.
Determination of native and polypeptide molecular mass using FPLC and SDS-PAGE: The purified lectin was dissolved in 2.5% glycerol and subjected to Gel Permeation Chromatography (GPC) utilizing a Superdex 75 column (1.0x65 cm) connected to an FPLC system (GE healthcare, USA) in the presence of 50 mM lactose containing TBS. The elution time of the lectin from the column was detected by UV at an absorbance of 280 nm. Bovine serum albumin (66 kDa), ovalbumin (43 kDa), carbonic anhydrase (30 kDa) and myoglobin (17 kDa) were used as standard molecular marker. The peptide size of the lectin was determined by SDS-PAGE (Laemmli, 1970). The concentration and separation gels were prepared as 5 and 12%, respectively and mixed with an equal volume of sample buffer (20 mM Tris-HCl, pH 6.8; 0.2% SDS and 20% glycerol in the presence or absence of 2% 2-mercaptoethanol) and then heated at 70°C for 15 min. Aliquots of 30 μL were applied to the well of a mini-slab gel (gel size: 80x100 mm with 1 mm thickness; 12.5 and 5% polyacrylamide were used in separation and upper gels, respectively). The molecular mass of the polypeptide was determined by SDS-PAGE (constant current at 30 mA for 1 h) according to a previous report (Laemmli, 1970). After electrophoresis, the gel was stained with 0.1% (w/v) Coomassie Brilliant Blue (CBB) R-250 in 40% methanol and 10% acetic acid and distained excessive staining by 40% methanol and 10% acetic acid. Crude supernatant obtained by preparative steps with or without lactose extracting was also separated by SDS-PAGE to appear the pattern of profile of proteins.
Hemagglutination and sugar inhibition assays of G. didyma lectin: The hemagglutination assay was performed using 1% (w/v) trypsinized and 0.25% glutaraldehyde-fixed rabbit and human erythrocytes as described previously (Matsui, 1984). Erythrocytes were suspended at a concentration of 1% (w/v) with TBS. In the general assay, 20 μL each of TBS, TBS containing 1% Triton X-100 and erythrocytes were added with 20 μL of the serially-diluted lectin with TBS in 96 well V-shape titer plates for 1 h. The hemagglutinating activity of the lectin was expressed as the titer defined as the reciprocal of the highest dilution giving positive hemagglutination. To determine the sugar binding specificity of the lectin, each 20 μL of the sugar (200 mM) and the glycoprotein, fetuin, asialofetuin and asialo-bovine submaxillary mucin (4 mg mL-1 each) were serially diluted with TBS and added with lectin to previously diluted TBS to adjust the titer to 16, 1% Triton X-100 and fixed erythrocytes were added in 96 well V-shape titer plates for 1 h. The minimum inhibitory sugar concentration against the lectin was expressed as a score of negative activity.
Kinetic analysis of the lectin using surface plasmon resonance: Since, asialofetuin strongly inhibited the hemagglutination of the lectin, the kinetic analysis was determined between the lectin and asialofetuin. Asialofetuin was dissolved in 10 mM citrate buffer (pH 5) was manually immobilized on a CM5 sensor chip using an amine coupling kit according to protocol after the activation of carboxylmethyl groups with dextran on the chip with a mixture of NHS (100 μL of 12 mg mL-1) and EDC (100 μL of 75 mg mL-1). The glycoprotein was conjugated on the sensor chip to be 4,200 resonance unit (RU). SPR analysis was performed with a BIAcore instrument (Model 3000, GE Healthcare, USA) whereby 90 μL of G. didyma lectin (0 to 12 nM) as an analyte was applied to the sensor chip for injection (association) and washing (dissociation) for 2.5 min each at a flow rate of 20 μL min-1. The association-rate constant (kass) and dissociation-rate constant (kdiss) were determined with BIA evaluation software Ver. 3.0 (Shinohara et al., 1994).
Preparation of lectin-conjugated affinity column and asialoglycoprotein capturing: Affinity column was prepared according to previously reported (Ozeki et al., 1995). One milligram purified lectin was dissolved into 100 mM sodium bicarbonate containing 150 mM has conjugated with NHS-activated Sepharose (1 mL) for 12 h in 4°C. Non-reacted groups on the medium were masked with 1 M ethanolamine pH 8 for overnight. Lectin-conjugated Sepharose was packed in small column and washed with TBS. Asialofetuin (1 mg mL-1) solubilized by TBS (1 mL) was applied the column. After washing the column with TBS, column bounded glycoprotein was eluted by 50 mM lactose in TBS. Each fraction (1 mL) was collected by fraction collector and eluted position of asialofetuin was confirmed by SDS-PAGE using separation gel at 15%.
RESULTS
Isolation of GdL: Crude extraction from bladder moon shell with TBS showed strong hemagglutinating activity against rabbit and human red blood cells. However, the activity was not cancelled by the co-presence of tested saccharides as galactose and lactose. Since, chilete reagents also did not inhibit the hemagglutinating activity, it suggested that the activity depends on the different mechanism from divalent cations. On the other hand, extraction of the precipitate with the homogenized by lactose containing TBS showed hemagglutination after the dialyzation with TBS. It was specifically inhibited by the co-presence of lactose. The profile of each extraction had slight different patterns by SDS-PAGE in reducing and non-reducing conditions (Fig. 1). G. didyma lectin (GdL) was purified on a lactosyl-agarose column via elution with 50 mM lactose containing TBS (Fig. 2) and it was shown to be a single 60 kDa polypeptide under both reducing and non-reducing conditions by SDS-PAGE, respectively (Fig. 3). Four mg of GdL was purified from 1,000 g (wet weight) of the shell and it was concentrated to 284 times folds by the affinity purification (Table 1). GdL was agglutinated rabbit and human erythrocytes (Table 2). The relative molecular mass of native GdL was estimated to be 60 kDa by GPC on a Superdex 75 column connected to an FPLC system using TBS or 50 mM lactose containing TBS (Fig. 4), indicating that GdL presented as a monomeric molecule consisting of a 60 kDa polypeptide.
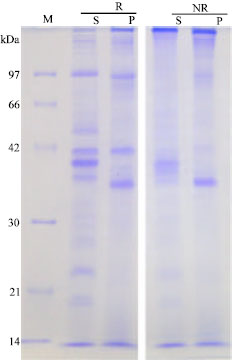 | |
| Fig. 1: | SDS-PAGE pattern of crude extraction of G. didyma. S: Supernatant of crude extract of foot homogenized by TBS. P: supernatant of the precipitate with 100 mM lactose containing G. didyma (R: Reducing and NR: Not reducing conditions). M: Standard marker proteins were used as follows, phosphorylase b (97 kDa); bovine serum albumin (66 kDa); ovalbumin (42 kDa); carbonic anhydrase (30 kDa); trypsin inhibitor (21 kDa); and lysozyme (14 kDa) |
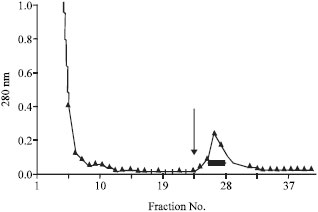 | |
| Fig. 2: | Affinity purification of GdL. Supernatant of the precipitate with 100 mM lactose containing G. didyma was applied to a lactosyl-agarose column equilibrated with TBS. The column was washed with TBS and eluted with TBS containing 50 mM lactose (arrow). Purified GdL was obtained by the dilution with TBS of the collected fractions (bar) |
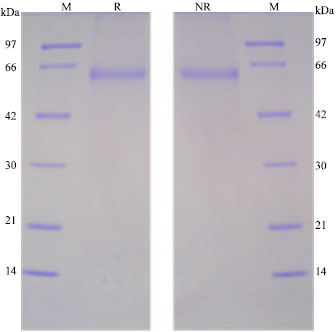 | |
| Fig. 3: | Purification of GdL and SDS-PAGE pattern. Purified GdL (10 μg) was subjected to SDS-PAGE under reducing (R) and non-reducing (NR) conditions. Standard marker proteins (M) were used as follows, phosphorylase b (97 kDa); bovine serum albumin (66 kDa); ovalbumin (42 kDa); carbonic anhydrase (30 kDa); trypsin inhibitor (20 kDa); and lysozyme (14 kDa) |
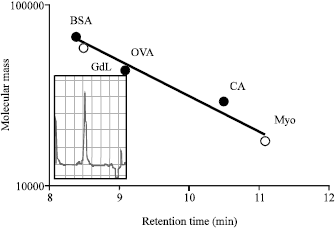 | |
| Fig. 4: | GPC and calibration curve for the determination of the molecular mass of GdL. Purified lectin (20 μg) was separated on a Superdex 75 column using an FPLC system at a flow rate of 0.5 mL min-1 (chart speed is 0.5 cm mL-1). Sample (50 μg) was dissolved in 10% glycerol containing TBS. Abbreviations indicate the standard molecular mass marker proteins: bovine serum albumin (BSA: 66 kDa); ovalbumin (OVA: 42 kDa); carbonic anhydrase (CA: 30 kDa); myoglobin (Myo: 17 kDa). The arrow shows the peak of GdL in GPC |
| Table 1: | Purification of lactose binding lectin from G. didymaaTotal activity is shown by Titerxvolume, bSpecific activity was shown by titer mg-1 of protein |
| Table 2: | Hemagglutination of rabbit and human erythrocytes by GdL |
 | |
| *Trypsinzed and glutaraldehyde fixed erythrocytes were used | |
| Table 3: | Saccharide recognition specificity of GdL |
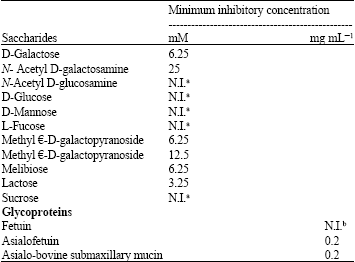 | |
| aInhibition was not occurred even at 100 mM, bFetuin did not inhibit even at 2 mg mL-1, Titer of lectin was previously diluted to 16 | |
Saccharides inhibit hemagglutination activity by GdL: Hemagglutination activity of GdL adjusted as hemagglutinating units into 16 was effectively inhibited by D-galactose, N-acetyl D-galactosamine, methyl α- and β-D-galactopyranoside, lactose, asialofetuin and asialo-bovine submaxillary mucin, indicating that GdL has saccharide binding specificity for D-galactose and β-D-galactoside. However, D-glucose, N-acetyl D-glucosamine and fetuin did not inhibit the activity even at over 100 mM (over 2 mg mL-1 in the case of fetuin) (Table 3).
Kinetic analysis of GdL: SPR analysis showed that the GdL bound asialofetuin on the sensor chip dose-dependently (Fig. 5). Kinetic parameters of kass and kdiss of GdL were 5.4x104 M-1sec-1 and 7.2x10-3sec-1, respectively by the fitting 1:1 affinity model using BIA evaluation 3.0 software.
Capture of glycoprotein by GdL-conjugated column: No asialofetuin appeared at the column pass-through fractions (Fig. 6, column 1-3). All of the glycoprotein was captured in the GdL-conjugated affinity column and it was selectively eluted by the application of 50 mM lactose containing TBS into the column (Fig. 6, column 4).
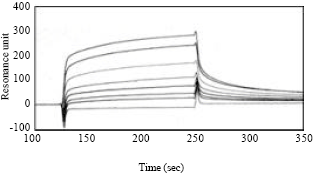 | |
| Fig. 5: | FKinetic analysis of GdL by SPR. GdL was applied onto the CM5 sensor chip coupled with asialofetuin at 4.2 ng mm-2. GdL (0-560 pM each) was applied onto the asialofetuin conjugated sensor chip with 20 μL min-1 for 2.5 min. The chip was washed with TBS for 2.5 min. X axis: Time (sec); Y axis: Resonance unit (indicating association and dissociation of analyte). Kinetics were analyzed by BIAevaluation software |
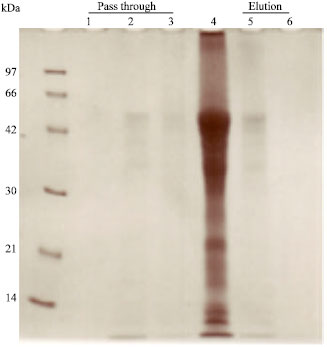 | |
| Fig. 6: | Capture of glycoprotein by GdL-conjugated column. No asialofetuin appeared at the column pass-through fractions (column 1-3). All of the glycoprotein was captured in the GdL-conjugated affinity column and it was selectively eluted by the application of 50 mM lactose containing TBS into the column (column 4) |
DISCUSSION
The characteristic carbohydrate-binding or biological properties of mollusk lectins are expectable to develop for new application of diagnosis as the lectin purified from sea hare gonad specifically recognizes both Bombay and para-Bombay types oligosaccharides (Gilboa-Garber et al., 2006). The lectin was also available to screen for bacteria and cancer associated antigen (Gilboa-Garber et al., 2001). Many lectins isolated from mollusks are galactophilic, however, some lectins have affinity to L-fucose- (Moy et al., 2008) and/or mannose (Gourdine et al., 2008). Even this study, GdL slightly recognizes N-acetyl D-galactosamine in addition to D-galactose. GdL had active in the absence of SH-preservating reagent nor divalent cations. This biochemical property on GdL was similar with that of lectin purified from cattlefish (Ozeki, 1997) and differed from the properties of both C-type lectin and galectin, though some lectins obtained from mollusks were reported to be them (Yamaura et al., 2008; Kim et al., 2008; Gourdine et al., 2008). To profile the precious glycan binding of the lectin as we did (Kawsar et al., 2008) and will be useful for develop on the application of the lectin. Besides, it has been found the tissue localizations and the change of expression level in mollusk lectins (Yamaura et al., 2008; Kim et al., 2006). Interestingly, not a few studies showed that the expression levels of these lectins depend on the infection of their parasite microorganisms, suggesting that lectins closely relate with outer environment and we develop on the bacterial aggregation activity for GdL at next. For the interaction of the lectin with endogenous or exogenous glycoconjugates, the binding affinity should be weak as it can reversibly bind. Surface plasmon resonance biosensor is a useful analytical instrument which can detect the biochemical interaction between immobilized ligand on the gold chip and analyte dynamically by the optical principal. It can detect the association (kass) and dissociation rate constant (kdiss) by the curve from the resonance according to the increase of mass on the gold surface by the binding of the analyte in mobile phase to ligand immobilized on the gold film. From the value of kass and kdiss, the kinetics analysis (KD) is available. The KD value of GdL was similar with that of representative plant lectin (Shinohara et al., 1994). Besides, GdL was not appeared in crude extraction of foot homogenized by TBS, but selectively extracted in the buffer containing lactose after homogenize of the precipitate with the buffer, suggesting that the lectin is binding with glycoconjugate in the foot tissue through the affinity between protein-carbohydrate interaction. Conjugation of GdL to gel for making lectin affinity column was also so useful to try to extract endogenous and exogenous ligands of the lectin. Since, the performance of the GdL-Sepharose gel was confirmed here to be able to catch the asialoglycoprotein, it will be passible to find endogenous ligands of the lectin from their tissues at next study. To show the diversity of oligosaccharides in invertebrates, this research strategy is valuable as a unique Gal-Fuc disaccharide has been discovered as an endogenous ligand of galectin-6 of nematode (Takeuchi et al., 2008), which is expected as the oligosaccharide structure of invertebrate differed from that of higher animals. Almost of the physiological function of GdL is unclear, however the evidence as the lectin presents not a little amount in their tissue suggest that GdL function for important biological phenomena. Some mollusk lectins has shown to aggregate bacterium or act for symbiosis with special marine microorganisms (Melo et al., 2000; Gourdine et al., 2007). In recent, addition to C-type lectin and galectin, sea urchin egg type lectin (Ozeki et al., 1991) also newly isolated from pearl shell (Naganuma et al., 2006). The presence of GdL in bladder moon shell suggest that ligand binding type lectin present in gastropod and both discovering the homologue in other species and finding the structural property of this type of lectin is our next research focus.
ACKNOWLEDGMENTS
This study was supported in part by 2007 Grants-in Aid for Scientific Research (No. 0760110001 to Y.O.) from JSPS (Japan Society for the Promotion of Science) and Strategic Research Project No. K20002 from Yokohama City University. We are thankful to Dr. Torao Sato for advice on the collection of materials. Authors are also appreciated Dr. Robert A. Kanaly for critical reading of this manuscript.
REFERENCES
- Banerjee, S., S. Chaki, J. Bhowal and B.P. Chatterjee, 2004. Muchin binding mitogenic lectin from freshwater Indian gastropod Belamyia bengalensis: Purification and molecular characterization. Arch. Biochem. Biophys., 421: 125-134.
CrossRef - Gilboa-Garber, N., D. Sudakevitz, C. Levene, N. Rahimi-Levene and V. Yahalom, 2006. H-dificient Bombay and para-Bombay red blood cells are most strongly agglutinated by the galactophilic lectins of Aplysia and Pseudomonas aeruginosa that detect I and P1 antigens. Immunohematology, 22: 15-22.
PubMed - Gilboa-Garber, N. and D. Sudakevitz, 2001. Usage of Aplysia lectin interaction with T antigen and poly-N-acetyllactosamine for screening of E. coli strains which bear glycoforms cross-reacting with cancer-associated antigens. FEMS Immunol. Med. Microbiol., 30: 235-240.
CrossRef - Gourdine, J.P., G. Cioci, L. Miguet, C. Unverzagt and D.V. Silva et al., 2008. High affinity interaction between a bivalve C-type lectin and a biantennary complex-type N-glycan revealed by crystallography a microcalorimetry. J. Biol. Chem., 283: 30112-30120.
PubMed - Gourdine, J.P., A. Markiv and J. Smith-Ravin, 2007. The three-dimensional structure of codakine and related marine C-type lectins. Fish Shellfish Immunol., 23: 831-839.
CrossRef - Kawsar, S.M.A., Y. Fujii, R. Matsumoto, T. Ichikawa and H. Tateno et al., 2008. Isolation, purification, characterization and glycan-binding profile of a D-galactoside specific lectin from the marine sponge, Halichondria okadai. Comp. Biochem. Physiol., 150B: 349-357.
CrossRef - Kim, J.Y., Y.M. Kim, S.K. Cho, K.S. Choi and M. Cho, 2008. Nobel tandem-repeat galectin of Manila clam Ruditapes philippinarum is induced upon infection with the protozoan parasite Perkinsus olseni. Dev. Comput. Immunol., 32: 1131-1141.
Direct Link - Kim, Y.M., K.I. Park, K.S. Choi, R.A. Alvarez, R.D. Cummings and M. Cho, 2006. Lectin from the Manila clam Ruditapes philippirum is indicte upon infection with the protozoan parasite Perkinsus olseni. J. Biol. Chem., 281: 26854-26864.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Melo, V.M., A.B. Duarte, A.F. Carvalho, A.F. Siebra and I.M. Vasconcelos, 2000. Purification of a novel antibacterial and haemagglutinating protein from the purple gland of the sea hare, Aplysia dactylomela ring, 1828. Toxicon, 38: 1415-1427.
CrossRef - Moy, G.W., S.A. Springer, S.L. Adams, W.J. Swanson and V.D. Vacquier, 2008. Extraordinaly interspecific diversity in oyster sporm binin. Proc. Natl. Acad. Sci. USA., 105: 1993-1998.
CrossRef - Naganuma, T., T. Ogawa, J. Hirabayashi, K. Kasai, H. Kamiya and K. Muramoto, 2006. Isolation, characterization and molecular evolution of a novel pearl shell lectin from a marine bivalve, Pteria peguin. Mol. Diversity, 10: 607-618.
CrossRef - Ozeki, Y., 1997. Purification of a 63 kDa β-d-galactoside binding lectin from cuttlefish, Todarodes pacificus. Biochem. Mol. Biol. Int., 41: 633-640.
CrossRefDirect Link - Sanchez, J.F., J. Lescar, V. Chazalet, A. Audfray and J. Gagnon et al., 2006. Biochemical and structural analysis of Helix pomatia agglutinin. A hexameric lectin with a novel fold. J. Biol Chem., 281: 20171-20180.
CrossRef - Shinohara, Y., F. Kim, M. Shimizu, M. Goto, M. Tosu and Y. Hasegawa, 1994. Kinetic measurement of the interactin between an oligosaccharide and lectins by a biosensor based on surface plasmon resonance. Eur. J. Biochem., 223: 189-194.
PubMedDirect Link - Smith, P.K., R.I. Krohn, G.T. Hermanson, A.K. Mallia and F.H. Gartner et al., 1985. Measurement of protein using bicinchoninic acid. Anal. Biochem., 150: 76-85.
CrossRefPubMedDirect Link - Takahashi, K.G., T. Kuroda and K. Muroga, 2008. Purification and antivacterial characterization of a novel isoform of the Manila clam lectin (MCL-4) from the plasma of the Manila clam, Ruditapes philippinarum. Comparative Biochem. Physiol., 150B: 45-52.
CrossRef - Takeuchi, T., K. Hayama, J. Hirabayashi and K. Kasai, 2008. Caenorhabditis elegans N-glycans cotaining a Gal-Fuc disaccharide unit linked to the innermost GlcNAc residue are recognized by C. elegans galectin LEC-6. Glycobiology, 18: 882-890.
PubMed - Wiechelman, K.J., R.D. Braun and J.D. Fitzpatrick, 1988. Investigation of the bicinchoninic acid protein assay: Identification of the groups responsible for color formation. Anal. Biochem., 175: 231-237.
CrossRefDirect Link - Yamaura, K., K.G. Takahashi and T. Suzuki, 2008. Identification and tissue expression analysis of C-type lectin and galectin in the Pacific oyster, Crassostrea gigas. Comparative Biochem. Physiol., 149B: 168-175.
CrossRef
Dr. A. K. M. Lutfur Reply
This article is new method to purify novel lectins from marine invertebrates. Thanks to authors.
Yasuhiro Ozeki Reply
Thank you for your attention to our article. Lectins in marine invertebrates should be interested by audiences in the journal, because it expects to be able to discover new molecule with various different carbohydrate binding properties. Our method using carbohydrate-conjugated affinity chromatography is a representative procedure for lectin purification. However, the point, as we used it for the purification of lectin in marine invertebrates and succeeded to isolate a new lectin with attractive biochemical properties is emphasised in the article. Best.