
K. Avina-Padilla
Centro de Investigacion y de Estudios Avanzados del Instituto Politecnico Nacional, Campus Guanajuato, km 9.6 Libramiento Norte, Carretera Irapuato-Leon, 36821 Irapuato, Gto., Mexico
F. Parra-Cota
Centro de Investigacion y de Estudios Avanzados del Instituto Politecnico Nacional, Campus Guanajuato, km 9.6 Libramiento Norte, Carretera Irapuato-Leon, 36821 Irapuato, Gto., Mexico
J.C. Ochoa-Sanchez
Centro de Investigacion y de Estudios Avanzados del Instituto Politecnico Nacional, Campus Guanajuato, km 9.6 Libramiento Norte, Carretera Irapuato-Leon, 36821 Irapuato, Gto., Mexico
C. Perales-Segovia
Instituto Tecnologico El Llano Aguascalientes, Apartado Postal 72-4, Administracion Postal 2, Ags., Mexico
J.P. Martinez-Soriano
Centro de Investigacion y de Estudios Avanzados del Instituto Politecnico Nacional, Campus Guanajuato, km 9.6 Libramiento Norte, Carretera Irapuato-Leon, 36821 Irapuato, Gto., Mexico
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 3 | Page No.: 268-271







ABSTRACT
This study was aimed to elucidate the putative causal agents. Healthy and diseased Echinopsis sp. and Opuntia sp. plants were collected from several Mexican nurseries. DNA was extracted from proliferating buds or stems and used in Polymerase Chain Reactions (PCR) to detect phytoplasmas. Two universal phytoplasma primer pairs were tested in nested PCR, initially with primer pair P1/tint followed by primer pair R16F2/R16R2 in a sequential test. The amplified DNA fragments were cloned and sequenced. Two different 16S rDNA partial operons were determined. One of present sequences was always associated to yellow mosaics in Echinopsis while the other to witches-broom syndromes in Opuntia. The comparative analysis of the sequences against the GenBank indicated that they were highly but not 100% homologous to phytoplasmas of group 16SrII. This is of major importance since this is the first report of finding representatives of this group affecting cacti in the Americas; all other reports of this kind of phytoplasmas have been detected in mainly in Asia.
PDF Abstract XML References Citation
How to cite this article
K. Avina-Padilla, F. Parra-Cota, J.C. Ochoa-Sanchez, C. Perales-Segovia and J.P. Martinez-Soriano, 2009. Phytoplasmas Associated to Diseases of Ornamental Cacti in Mexico. Journal of Biological Sciences, 9: 268-271.
DOI: 10.3923/jbs.2009.268.271
URL: https://scialert.net/abstract/?doi=jbs.2009.268.271
DOI: 10.3923/jbs.2009.268.271
URL: https://scialert.net/abstract/?doi=jbs.2009.268.271
INTRODUCTION
America is the center of origin of cacti plants being Mexico a rich region in their biodiversity. In the last years stems and fruits are used as nutritional source but its demand as ornamental plants has been grown substantially. Recently, an increase in field diseases has been observed and a number of new varieties with unusual anatomical characteristics are offered to cacti collectors in national nurseries. These cacti plants show abnormal growth with excessive proliferation of buds, yellow stems, yellow mosaics and purple colorations (Fig. 1). Some of these syndromes have been previously associated to phytoplasmas by present group in scientific meetings (Leyva-López et al., 1999).
Phytoplasmas are nonculturable degenerate gram-positive prokaryotes related with mycoplasmas and spiroplasmas. Lee et al. (2000) have classified these pathogens in 14 main groups according to restriction enzyme patterns of their 16SrRNA genomic operons (16SrI-XIV). These pathogenic agents are usually transmitted by insect vectors and are associated to more than 700 diseases worldwide (Weintraub and Beanland, 2006; Bertaccini, 2007).
Phytoplasmas are associated to a high number of plant diseases in Mexico, being the so-called potato purple top the most notorious due to its financially impact on the Mexican potato industry. Present studies led to conclude that two different phytoplasmas were involved in the potato purple top syndrome (Leyva-López et al., 2002). One of them was identified as potato hair sprouts phytoplasma and is a member of the group 16SrII, being this parasite the first one found in Mexico, since all the reported members have been described affecting plants is Asia. Recently, Lee et al. (2006) reported the presence of another member of this group that might be the same phytoplasma reported in 2002.
The potato hair sprouts phytoplasma was found infecting cacti growing as weed or barriers in the potato fields as well (Leyva-López et al., 2002). Very few phytoplasmas associated to cacti species have been described so it was of interest to investigate its presence in other succulent plants.
It is important to emphasize that these pathogens confer unusual characteristics that make cacti plants attractive in commercial nurseries. Objectives of this study included the detection and molecular analysis of any phytoplasma related to the observed syndromes affecting ornamental cacti being commercialized in Mexican nurseries.
 | |
| Fig. 1: | Cacti displaying phytoplasma-associated symptoms. Panel (A) Opuntia sp. showing phyllody, (B) Opuntia sp. showing witches’ broom, (C) Opuntia sp. showing cladode mosaics |
MATERIALS AND METHODS
DNA isolation: Cacti samples of affected plants were collected from plant nurseries located in the Mexican States of Guanajuato and Colima during 2006-2007. Fresh tissue in active growth was collected and used for DNA isolation and processing using the method reported by Lopes and Larkins (1993).
PCR assays: Universal primer pairs were used for the amplification of the 16SrRNA gene and the 16S/23S spacer region. The universal phytoplasma primers P1 (5’-aagagtttgatcctggctcaggatt-3’) and tint (5’-tcaggcgtgtgctctaaccagc-3’) were used to amplify the 16S rRNA and tRNA operon region in a first PCR (Smart et al., 1996). PCR was performed in a 25 μL total volume of reaction, containing 100 ng of total DNA, 10 pmol of each primer, 1x of buffer solution for PCR, 2 mM MgCl2, 200 mm of dNTPs and 2.5 units of Taq DNA polymerase (PROMEGA Corp. Madison WI). PCR rounds were 3 min to 90°C followed by 30 cycles of three steps: 1 min to 90°C, 1 min at 55°C and 1 min at 37°C. Products obtained in the first amplification were used to made 1:20 dilutions and 1 μL was taken from each dilution like DNA template to make the second amplification for samples that could have low titer of phytoplasmas, where the pair of internal initiators R16F2 (5’-catgcaagtcgaacgga-3’)/R16R2 (5’-tgacgggcggtgtgtacaaaccccg-3’) (Lee et al., 1994) were used with the conditions of reaction mentioned previously. Products were visualized in agarose 1% gels. PCR amplification was conducted in an automated thermocycler (PT-100, MJ Research, San Francisco CA, USA).
Cloning and nucleotide sequencing: RFLP-PCR and DNA sequences analysis were used for molecular characterization. PCR products were digested with Kpn I, Hinf I and Alu I (Promega Corporation, Madison, WI, USA) and visualized in agarose 1.5% gels for enzymatic restriction analysis. Intact amplified products were purified with Wizard PCR preps Kit and inserted into pGEM-T vector Easy Vector System II (Promega, Madison, WI) later used to transform Escherichia coli DH5alfa cells. Plasmids containing the expected sizes were chosen for automatic sequencing using an ABI PRISM 377 PERKIN-ELMER DNA sequencer.
Sequence alignment and phylogenetic analysis: Restriction site maps of the 16S rRNA gene of the phytoplasmas found here were generated using the CLC Free Workbench 4.5.1 program for the Macintosh operating system (Cambridge, MA, USA). DNA sequences were compared to each other and with the existing sequences at the GenBank database using the same software and the NCBI Blast program (Zhang et al., 2000).
RESULTS AND DISCUSSION
PCR assays using the P1/Tint pair allowed the amplification of a 1650 bp fragment in a first reaction, followed by the R16F2/R16R2 pair that yielded the a 1250 bp amplified fragment. PCR analysis demonstrated that cacti plants are important phytoplasma hosts because they maintain a high titre apparently (data not shown).
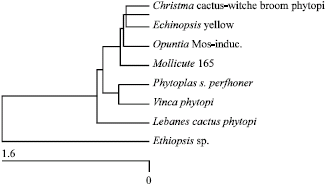 | |
| Fig. 2: | Dendrogram based on sequence analysis showing the cacti phytoplasma sequences presented in this report |
These plants may be important natural hosts for the phytoplasmas to be dispersed by insects to other plants of agronomic importance (Weintraub and Beanland, 2006).
RFLP analysis of the amplified fragments revealed the presence of a single phytoplasma group. Present results showed the presence of single phytoplasma DNA in each screened plant and no mixed RFLPs patterns were found in any case. Different titres of phytoplasmas were observed as suggested by Firrao et al. (2007) and Marzachi and Bosco (2005). Restriction analysis of amplified fragments showed a clear difference between two clones. DNA sequencing of the R16F2/R16R2 amplified region yielded two different sequences belonging to the 16SrII group according to their RFLP patterns (Wei et al., 2007). 16S RNA RFLP comparison between Opuntia sp. Mosaic-inducing phytoplasma and Christmas Cactus Witches broom phytoplasma shows that both maps are identical, but the comparison of nucleotide sequences indicated that they are different phytoplasmas (Fig. 2).
DNA sequence analysis indicated that two of the phytoplasmas described here have not been reported previously at the GenBank database. As result of this work the 16S rDNA sequences of the phytoplasmas associated with cacti were registered as Opuntia sp. Mosaic-inducing phytoplasma (accession No. DQ535899) and Echinopsis sp. yellow patch phytoplasma (accession No. DQ535900).
An important profit in this study is the identification of phytoplasmas associated to two types of syndromes of cacti plants becoming in the first report of phytoplasmas of the 16SrII group present in the Americas. Their presence in Mexico is difficult to explain but it could be speculated that these phytoplasmas arrived to our country via., ornamental plants imported from Asia, where this group (16SrII) has been repeatedly reported.
Thus, the phytoplasmas named here as Opuntia sp. mosaic-inducing phytoplasma (DQ535899) and Echinopsis sp. yellow patch phytoplasma (DQ535900) belong to the 16SrII group. It is remarkable to emphasize that cacti species that are alternating hosts for phytoplasma are common in commercial nurseries and they have a great demand because of the unusual characteristics that make them more attractive may involve involuntary risk of dispersion of diseases to crops of agronomic importance.
ACKNOWLEDGMENTS
KAP and FPC have Ph. D scholarships from Consejo Nacional de Ciencia y Tecnología (CONACYT), Mexico. We also wish to thank to Consejo de Ciencia y Tecnología del Estado de Guanajuato (CONCYTEG) for providing the grant Fitoplasmas de importancia agrícola presentes en viveros del Estado de Guanajuato code 08-03-k662-076. We are grateful with Verónica Sánchez-Briseño, Maribel Moreno-Valenzuela and Claudia Castro-Transviña for their technical support.
REFERENCES
- Firrao, G., M. Garcia-Chapa and C. Marzachi, 2007. Phytoplasmas: Genetics, diagnosis and relationships with the plant and insect host. Front Biosci., 12: 1353-1375.
PubMedDirect Link - Lee, I.M., R.E. Davis and D.E. Gundersen-Rindal, 2000. Phytoplasma: Phytopathogenic mollicutes. Ann. Rev. Microbiol., 54: 221-555.
CrossRefPubMedDirect Link - Lee, I.M., D.E. Gundersen, R.W. Hammond and R.E. Davis, 1994. Use of mycoplasmalike organism (MLO) group-specific oligonucleotide primers for nested-PCR assays to detect mixed-MLO infections in a single host plant. Mol. Plant Pathol., 84: 559-566.
CrossRefDirect Link - Lopes, M.A. and B.A. Larkins, 1993. Endosperm origin, development and function. Plant Cell, 5: 1383-1399.
PubMedDirect Link - Marzachi, C. and D. Bosco, 2005. Relative quantification of chrysanthemum yellows (16Sr I) phytoplasma in its plant and insect host using real-time polymerase chain reaction. Mol. Biotechnol., 30: 117-128.
CrossRefPubMedDirect Link - Smart, C.D., B. Schneider, C.L. Blomquist, L.J. Guerra and N.A. Harrison et al., 1996. Phytoplasma-specific PCR primers based on sequences of the 16S-23S rRNA spacer region. Phytopathol., 62: 2988-2993.
PubMedDirect Link - Wei, W., R.E. Davis, I.M. Lee and Y. Zhao, 2007. Computer-simulated RFLP analysis of 16S rRNA genes: Identification of ten new phytoplasma groups. Int. J. Syst. Evol. Microbiol., 57: 1855-1867.
CrossRefPubMedDirect Link - Weintraub, P.G. and L. Beanland, 2006. Insect vectors of phytoplasmas. Ann. Rev. Entomol., 51: 91-111.
CrossRefPubMedDirect Link - Zhang, Z., S. Schwartz, L. Wagner and W. Miller, 2000. A greedy algorithm for aligning DNA sequences. J. Comput. Biol., 7: 203-214.
CrossRefPubMedDirect Link - Bertaccini, A., 2007. Phytoplasmas: Diversity, taxonomy and epidemiology. Front Biosci., 12: 673-689.
PubMedDirect Link - Lee, I.M., K.D. Bottner, G. Secor and V. Rivera-Varas, 2006. Candidatus Phytoplasma americanum, a phytoplasma associated with a potato purple top wilt disease complex. Int. J. Syst. Evol. Microbiol., 56: 1593-1597.
Direct Link