
Hortensia Brito-Vega
Universidad Juarez Autonoma de Tabasco, Division Academica de Ciencias Agropecuarias, Km 25.5 Carretera Villahermosa-Teapa CP8600, Tabasco
David Espinosa-Victoria
Colegio de Postgraduados, Laboratory of Plant-Microorganism Molecular Interaction, Km 36.5 Carretera Mexico-Texcoco, Montecillo, Estado de Mexico, C.P. 56230
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 3 | Page No.: 192-199







ABSTRACT
Anecic, epigeous and endogeous earthworms stimulate or inhibit the growth of bacteria of agricultural importance inside their digestive tracts. It is possible that these bacteria establish a mutual symbiosis within the digestive tract of the earthworm. The bacterial species reported within the intestines of the earthworms belong to the genuses Bacillus, Aeromonas, Pseudomonas, Flavobacterium, Nocardia, Gordonia, Vibrio, Clostridium, Proteus, Serratia, Mycobacterium, Klebsiella, Azotobacte and Enterobacter. These bacteria inhabit the soil and develop considerably when there are easily degradable organic soil nutrients. The bacterial community inside the digestive tract of earthworms pertains to at least four physiological groups: plant growth promoters, free-living nitrogen fixers, biocides and phosphate solubilizers. The diversity of bacterial communities within the digestive tracts of earthworms depends on climate, soil type and organic matter. The objective of this present study was to analyze the state of art on the bacterial diversity within the digestive tracts of earthworms.
PDF Abstract XML References Citation
How to cite this article
Hortensia Brito-Vega and David Espinosa-Victoria, 2009. Bacterial Diversity in the Digestive Tract of Earthworms (Oligochaeta). Journal of Biological Sciences, 9: 192-199.
DOI: 10.3923/jbs.2009.192.199
URL: https://scialert.net/abstract/?doi=jbs.2009.192.199
DOI: 10.3923/jbs.2009.192.199
URL: https://scialert.net/abstract/?doi=jbs.2009.192.199
INTRODUCTION
Anthrotropic activities such as deforestation, contamination and the advance of urban sprawls, lead the gradual disappearance of forests and other natural vegetation along with soil fauna which makes up the mega a macrofauna (Hernández-García and Granados-Sánchez, 2006; Huhta, 2002).
Earthworms are an important part of the macrofauna, visible with the naked eye (>10.000 μm) and represent 82% of the total biomass in tropical zones with a precipitation above 1000 mm (Lavelle et al., 2006). The diversity of these macroorganisms depends on biotic conditions and factors such as temperature, humidity, apparent density, pH and organic matter which form part of their diet (Curry and Schmidt, 2007). Currently more than 7.245 species of Oligochaetes have been classified at global level, from which 4000 earthworms species are described (Fragoso, 2001; Reynolds, 1998). In Mexico, 129 species have been identified from which 46 are native, 47 exotic and 36 others in which the majority belong to the family Megascolecidae (Fragoso, 2001).
Earthworms intervene in soil biological regulation systems, posses the capacity to remove soil particles and produce organomineral structures called biogenic structures (Rossi et al., 2006). They also help to maintain soil structure, water infiltration and regulate the availability of nutrients assimilated for plants, which includes nitrogen (N) in the form of ammonia (NH4+) and nitrates (NO3¯) (Desjardins et al., 2003).
Earthworms mechanically mix mineral particles and organic matter through their digestive system which carries out disintegration, grinding and digestion of the ingested material, increasing or decreasing the activity and number of beneficial or pathogenic microorganisms (fungi, Actinomycetes and bacteria) (Winding et al., 1997). The participation of microorganisms within the digestive tracts of earthworms is of great importance given that a lot of these are involved in the degradation of organic matter (Byzov et al., 2007). For studies on bacteria within the intestines of earthworms, diverse methods and techniques have been used which have helped in identifying species of the genuses Bacillus, Pseudomonas, Klebsiella, Azotobacter, Serratia, Aeromonas and Enterobacter (Valle-Molinas et al., 2007; Byzov et al., 2007; Singleton et al., 2003). These bacteria are mainly plant growth promoters, free-living nitrogen fixers and phosphate solubilizers (Loreno-Osti et al., 2004; Martínez-Romero, 2001). Some researchers have indicated the existence of a possible type of mutualism between these two organisms (Brown et al., 2000; Barois and Lavelle, 1986). Therefore, the objective of this study was to carry out a revision of the state of art of the role of these earthworms and the bacterial diversity within their digestive tracts.
TAXONOMY AND ECOLOGY OF EARTHWORMS
Earthworms have been studied during the last years due to their importance in areas such as biotechnology, ecotoxicology, morphology, ecology, taxonomy, soil physics and soil fertility (Amador and Görres, 2007; Aira et al., 2003; Fraser et al., 2003; Homa et al., 2003; Spurgeon et al., 2003).
Earthworms have been classified within the phylum: Annelida, order: Oligochaeta and class: Clitella (Edwards and Lofty, 1977). The order Oligochaeta consists of 12 families: Moniligastridae, Megascoleidae, Ocnerodrilidae, Acanthodrilidae, Octochaetidae, Eudrilidae, Glossoscolecidae, Sparganophilidae, Microchactidae, Hormogastridae, Criodrilidae and Lumbricidae (Edwards and Lofty, 1977). From an agricultural point of view, the most important family is Lumbricidae and includes the genuses Lumbricus, Aporrectodea, Allolophora, Dendrobaena, Eisenia, Helodrilus, Octalasion and Eophila (Edward, 2004). The family Glossoscolecidae includes two genuses, Periscolex and Pontoscolex, which are of agricultural importance in tropical zones (García and Fragoso, 2003; Reynolds, 1998).
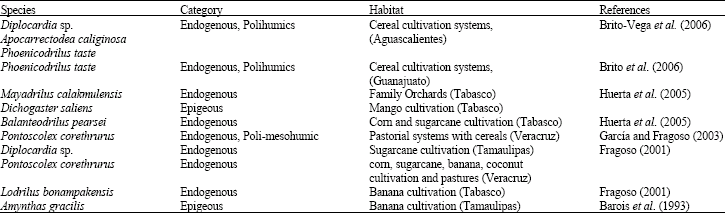
From an ecological point of view, earthworms have been classified in three categories: (1), epigeous, which live within the soil, interact with edaphic microbiota and modify soil profile with their underground galleries and excrements; (2) endogenous, which transform dead fallen leaves, are soil consumers and are subdivided in poli, meso and oligohumics, with reduced interaction with microbiota and (3) anecic, which lives in and consumes leaf litter (Curry and Schmidt, 2007). This classification is used in diverse studies in order to establish the ecological category of these invertebrates under different agricultural systems in the Mexican Republic (Table 1).
In Mexico 129 species of earthworms have been identified where only 10 species have been studied from a point of view based on population, soil fertility and plant growth (Fragoso, 2001). The exotic endogenous specie Pontoscolex corethrurus is considered geophagous and is one of the most researched species in tropical zones (García and Fragoso, 2003; Barois et al., 1993; Barois, 1992).
Earthworms affect processes within the soil in a direct (incorporation and redistribution of several organic and inorganic materials, aeration, moisture distribution, infiltration) or indirect manner (formation of microbial communities, transportation of propagules and inhibition of pathogens) (Byzov et al., 2007).
To some degree, the functions carried out by earthworms depend on the efficiency of their digestive systems which comprises of: mouth, pharynx, crop, intestines and anus (Fig. 1). These invertebrates interact with soil microorganisms, as well as biological structures known as turricules (feces) which they produce within the soil (Curry and Schmidt, 2007).
| Table 1: | Ecological categories of earthworm species under different agricultural systems in Mexico |
 | |
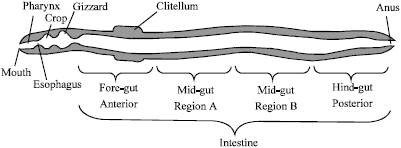 | |
| Fig. 1: | Diagram of the digestive system of an earthworm (Horn et al., 2003) |
Within their digestive systems, enzymatic activity is stimulated and may promote or inhibit the proliferation of certain fungal, actinomycetes and bacterial communities (Byzov et al., 2007; Barois, 1992).
DIVERSITY OF SOIL BACTERIA
Earthworms posses an immense bacterial diversity within their digestive tracts and is very little explored mainly because of the non-cultivatable character of a large quantity of microorganisms which mainly come from soil.
Soil is an appropriate environment for the development of eukaryotic (algae, fungi, protozoa) as well as prokaryotic (bacteria and archeas) microorganisms. Virus and bacteriophage are also present (Nogales, 2005). All these organisms establish relationships among themselves in highly varied and complex ways which contribute to soil characteristics because of their role in the modification of solid, liquid and gaseous stages.
Plant beneficial prokaryotes may have a considerable potential as biocontrol agents and biofertilizers. Two large groups are distinguished: (a) nitrogen fixing microorganisms and (b) plant growth promoting bacteria. The last group of bacteria is known as PGPR (plant growth promoting rhizobacteria) or as root living bacteria which significantly stimulate plant growth (Loreno-Osti et al., 2004; Espinosa-Victoria et al., 2006).
Nitrogen fixing prokaryote microorganisms are classified into two groups: (1) obligated symbiotic, which infects the roots of legumes and (2) non-obligated symbiotics or free-living, which establishes relations with a range of gramineous plants. Within the second group, bacterial species from some genuses like Azospirillum, Acetobacter, Azotobacter, Beijerinckia, Pseudomonas, Bacillus and Vibrio have been reported (Bashan et al., 2004; Young et al., 2001).
Another group of soil bacteria is the Phosphate Solubilizing Bacteria (PSB) which perform an important role in supplementing di and monobasic phosphorous to plants. There are studies which have demonstrated the capacity of different species of bacteria which solubilize this insoluble phosphate and some of these bacteria belong to the genuses Bacillus, Enterobacter, Erwinia, Pseudomonas, Rhizobium, Serratias, Agrobacterium, Burkholderia, Achromobacter, Microccocus, Aerobacter, Flavobacterium and Erwinia (Fernández et al., 2005).
Rizopheric bacteria are capable of producing physiologically active substances such as vitamins, giberelines, citochinines and Acetic Indole Acid (AIA) in significant quantities. The genuses Azospirillum and Klebsiella produce auxin (AIA), which causes morphological changes in the root and is related with the absorption of minerals mainly in corn and teocintle (Carcaño et al., 2006; Cattelan et al., 1999).
The alternative of using bacteria as a control agent against pathogens has developed certain bacterial strains with a wide spectrum of effectivity. Some species include Bacillus subtilis, B. cereus, B. thuringiensis, B. cepacia, Pseudomonas aerufasciens, P. chlororhapis, P. corrugata, P. fluorescens, P. putida, Burkholderia cepacia, Enterobacter sp. BF 14, Serratia plymuthica, Serratia marcescens, Agrobacterium sp. (De Lima-Ramos et al., 2004).
Soil bacteria are not randomly distributed and thus follow special aggregation patterns at different scales (from nm to km) which they superimpose. This structuring obeys the effect caused by different control factors and is completely dynamic (Ettema et al., 2002). Bacteria organize themselves in micro colonies comprised of few cells that may pertain to different morphotypes (Nunan et al., 2003).
MICROBIAL DIVERSITY WITHIN THE DIGESTIVE WITHIN THE DIGESTIVE TRACTS OF EARTHWORMS
Plant detritus in soil is the main nutritional source for earthworms although a few microorganisms like protozoans, Actinomycete, bacteria and fungi have been found to be part of the diet of these worms (Table 2) (Byzov et al., 2007; Hyun-Jung et al., 2004; Furlong et al., 2002). This relationship between microorganisms and Oligochaetes is not necessarily limited to a predatory process, as it has been demonstrated that the digestive enzymes produced by the earthworms do not significantly affect some microorganisms (Barois et al., 1993).
One of the first forefathers on microbial presence in the digestive system was Parle (1963), who reported bacterial, fungal and Actinomycetic populations in three species of earthworms; Lumbricus terrestres, Allolobophora caliginosa and A. longa.
| Table 2: | Microbial presence within the intestines of different species of earthworms |
 | |
A study carried out by Kristufek et al. (1994) found populations of bacteria, actinomycetes, fungi, sterile mycelium and plant cells in soil, however within the intestines of the earthworm Lumbricus rubellus, these organisms were found lysate except for a few actinomycetes, endospores and encapsulated bacteria. On the other hand, Márialigeti (1979) found that the microbial flora within the posterior segment of the intestines of Eisenia lucens, contained 473 organisms where 73% pertain to the genus Vibrio.
Contreras (1980) reported that 70% of flora in the intestines of Eisenia lucens was represented by only one species of Actinomycete, Streptomyces lipmanii, an organism rarely found in nature. On the other hand, Kristufek et al. (1993) identified two species of Actinomycetes denominated Streptomyces diastatochromogenes and Streptomyces noglalater, which are characteristic of soil within the digestive tracts of two earthworm species: Lumbricus rubellu and Octolasion montanum.
Dash et al. (1986) did a microfungal characterization in the digestive tract of the three species of earthworms (Ortochaetona surensis, Lampito maurittii and Drawida willsi) found in the tropical zones of India and identified 18 species of fungi from the genuses Aspergillus, Penicillium, Thielavia, Botryotrichum, Fusarium, Rhizopus, Curvularia, Chaetomium and Trichoderma. Four more genuses, namely Neocosmospora, Cladosporium, Syncephalastrum and Actinomucor were found in L. mauritii, unlike the other two species of earthworms. It is important to mention that the digested material for all three species came from organic waste.
Kristufek et al. (1992, 1993) observed and increase in the number of bacteria, Actinomycetes and fungi in the anterior section of the digestive tract of Lumbricus rubellus while the opposite occurred in Aporrectodea caliginosa and A. caliginosa.
The Actinomycete community present in the intestine of Eisenia fetida, mainly Streptomyces caeruleus, develops better in the intestines in comparison to soil and helps the earthworms to metabolize organic matter and decomposition of substances from plant origin (Polyanskaya et al., 1996).
BACTERIAL DIVERSITY IN THE DIGESTIVE TRACT OF EARTHWORMS
Soil is the key system in the functioning of terrestrial ecosystems. Vital processes take place within this system: decomposition and nutrient flow (Bashan et al., 2004) Biological activities control theses processes, among them, prokaryotes and earthworms (Table 3) (Davidson and Stahl, 2006; Fragoso et al., 2001).
Jolly et al. (1993) demonstrated the existence of physical contact between some filamentous, segmented bacteria and intestinal mucus of the species Octolasion lacteum and Lumbricus terrestris. The results showed bacterial filaments joined to the intestinal walls of the earthworms by means of hook structures. Therefore, it is concluded that the bacteria may be adapted to live within the intestines of the earthworms. Thus diverse studies on the main bacteria within the intestinal walls of earthworms have been conducted.
An observation was made for the specie Eisenia fetida coming from contaminated soil in an industrial zone. There was an increase in 91 colonies, further divided into 12 groups: Aeromonas 6%, Agromyces 3%, Bacillus 31%, Bosea 1%, Gordonia 6%, Klebsiella 6%, Microbacterium 7%, Nocardia 2%, Pseudomonas 10%, Rhodococcus 19%, Tsukamurella and Streptomyces 7%. The genus Bacillus was the dominant group found in the intestines of the earthworm (Hyun-Jung et al., 2004).
| Table 3: | Bacterial diversity identified within the intestinal walls of earthworms |
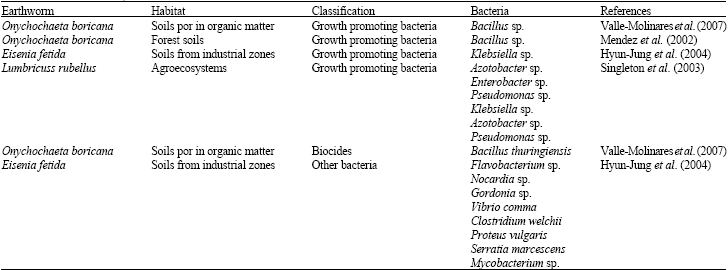 | |
The specie Eisenia fetida may establish a symbiotic relationship with bacteria from the genus Acidovorax, given that these bacteria form nodules in the ampules of the nephridium of the earthworms and help in the process of protein decomposition (Davidson and Stahl, 2006).
On the other hand, Valle-Molinares et al. (2007) identified seven species of bacteria from the genus Bacillus: (B. insolitus, B. megaterium, B. brevis, B. pasteurii, B. sphaericus, B. thuringiensis and B. pabuli) within the intestines of Onychochaeta borincana. All these species are typical soil bacteria. In addition, it was found that the microbial weight of the intestinal region decreased from the anterior to posterior section. Additionally, it was observed that some bacteria increased in the posterior section of the intestines, maybe because for a many bacteria this portion presents adequate conditions for their development.
Mendez et al. (2003) indicated that the bacteria can accomplish a type of mutualism during their passage through the digestive tracts of earthworms, which have not yet been studied in other genuses of bacteria. The existence of a mutual symbiotic relation between O. boricana and B. cereus has been proposed.
MOLECULAR TECHNIQUES USED IN THE STUDY OF BACTERIAL DIVERSITY WITHIN THE DIGESTIVE TRACTS OF EARTHWORMS
The use of molecular techniques has manifested the unawareness of biological diversity, systemic classification and taxonomy given that the majority of microorganisms are not cultivatable in conventional mediums and also the analysis of functional genes key in important soil processes such as denitrification, nitrification nitrogen fixation and methane oxidation (Nogales, 2005).
Table 4 shows some molecular techniques used in the identification of bacteria found within the digestive tracts of earthworm, applying basic microbiological culture techniques (Santiago, 1995). Many forms of microscopy (Krištufek et al., 1994) and molecular biology (Hyun-Jung et al., 2004; Singleton et al., 2003) (with the last being currently one of the most used techniques) and its application in soil microbiology studies mainly in the digestive tracts of earthworms, represent a great advance in the knowledge of different ecosystems.
The study of the diversity of microorganism is currently based on protein analysis, DNA or RNA of the ribosomal genes 16S or 23S and the presence of enzymes or enzyme alleles (Curry and Schmidt, 2007).
The identification process of bacteria associated with the intestines of earthworms is difficult if these microorganisms require growing conditions present only in the intestines of the earthworms. Singleton et al. (2003) used molecular techniques such as Polymerase Chain Reaction (PCR), for identifying bacteria associated with the intestines of Lumbricus rubellus, which were absent from the earthworms skins.
Fluorescent microscopy has been used for studying population variations of the bacteria within each intestinal segment of the Oligochaetes.
Fischer et al. (1995, 1997) studied the digestive intestinal walls of O. borincana using trace electronic microscopy and found bacteria from the genus Bacillus adhered to the intestine of this oligochaete being abundant in the frontal area and less abundant in the posterior parts. The researchers suggested the smooth surface of the posterior parte offers less possibility of adhesion for the bacteria on the intestinal wall but is appears that they may have found a favorable environment in the posterior part.
CONCLUSIONS
The importance of microbial diversity using conventional and molecular techniques is still far from understanding the role of the microorganisms within the digestive tracts of earthworms and within the soil in the functioning of ecosystem, particularly in those which have not being laboratory grown and for those whose metabolic capacities is totally unknown. They are key in important soil processes such as denitrification, nitrification, nitrogen fixation, methane oxidation, growth hormone production, phosphorous solubilizers and control of microbial pathogens.
| Table 4: | Molecular techniques used in the identification of bacterial diversity within the digestive tract of earthworms |
 | |
The bacterial diversity within the digestive tracts of earthworms from different genuses and ecotypes presents a variety of geniuses and prokaryote species, attributed to their habitat, soil type, climate, substrate type and biota.
The earthworm is an incubator and disperser of bacteria of agricultural importance given that some species found, produced microbial control of certain pathogens (Burkholderia, Enterobacter, Agrobacterium Serratia, Pseudomonas), plant growth promoters through the production of the indole-3-acetic acid (Klebsiella, Azotobacte, Aeromonas, Bacillus), produced siderophore (Pseudomonas, Bacillus, Proteus) and phosphate solubilizers unavailable to plants (Bacillus, Enterobacter, Erwinia, Pseudomonas, Rhizobium, Serratias, Agrobacterium, Burkholderia, Achromobacter, Microccocus, Aerobacter, Flavobacterium and Erwinia).
The genetic characterization of the isolated bacteria allows us to broaden our knowledge of the bacterial diversity within the different structures or parts of the digestive tracts and under different natural ecosystems presenting anthropogenic activities and the study of plant effect.
ACKNOWLEDGMENTS
National Science and Technology Council (Conacyt) for the economic support as scholarship student 208219, to the Juaréz Autonomous University of Tabasco (UJAT) for the valuable support and to the Postgraduate College as the institution for boarding and in the formation of the Doctorate in Agricultural Sciences.
REFERENCES
- Aira, M., F. Monroy and J. Domínguez, 2003. Effects of two species of earthworms (Allolobophora spp.) on soil systems: A microfaunal and biochemical analysis. Pedobiologia, 47: 877-881.
CrossRef - Amador, J.A. and J.H. Gorres, 2007. Microbiological characterization of the structures built by earthworms and ants in an agricultural field. Soil Biol. Biochem., 39: 2070-2077.
CrossRef - Barois, I., 1992. Mucus production and microbial activity in the gut of two species of Amynthas (Megascolecidae) from cold and warm tropical climates. Soil Biol. Biochem., 24: 1507-1510.
Direct Link - Barois, I., G. Villemin, P. Lavelle and F. Toutain, 1993. Transformation of the soil structure through Pontoscolez corethrurus (Oligochaeta) intestinal tract. Geoderma, 56: 57-66.
CrossRefDirect Link - Barois, I. and P. Lavelle, 1986. Changes in respiration rate and some physcochemical properties of a tropical soil during transit through Pontoscolex corethrurus (Glossoscolecidae, Oligochaeta). Soil Biol. Biochem., 18: 539-541.
CrossRefDirect Link - Bashan, Y., G. Holguin and L.E. De-Bashan, 2004. Azospirillum-plsnt relation-ships: Physiological, molecular, agricultural and environmental advences (1997-2003). Can. J. Microbiol., 50: 521-577.
Direct Link - Brito-Vega, H., D. Espinosa-Victoria, B. Figueroa-Sandoval, C. Fragoso and J.C. Patrón-Ibarra, 2006. Diversidad de lombrices de tierra con labranza de conservación y convencional. Terra Latinoamericana, 24: 99-108.
Direct Link - Brown, G.G., I. Barois and P. Lavelle, 2000. Regulation of soil organic matter dynamics and microbial activityin the drilosphere and the role of interactionswith other edaphic functional domains. Eur. J. Soil Biol., 36: 177-198.
CrossRefDirect Link - Byzov, B.A., N.V. Khomyakov, S.A. Kharin and A.V. Kurakov, 2007. Fate of soil bacteria and fungi in the gut of eartworms. Eur. J. Soil Biol., 43: 146-156.
CrossRef - Carcaño, M.M.G., R. Ferrera-Cerrato, J. Pérez-Moreno, J.D. Molina-Galán and Y. Bashan, 2006. Actividad nitrogenasa, producción de fitohormonas, sideróforos y antibiosis en cepas de Azospirillum y Klebsiella aisladas de maíz y teocintle. Terra Latinoamericana, 24: 493-502.
Direct Link - Cattelan, A.J., P.G. Hartel and J.J. Fuhrmann, 1999. Screening for plant growth-promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J., 63: 1670-1680.
CrossRefDirect Link - Contreras, E., 1980. Studies on the intestinal actinomycete Flora of Eisenia lucens (Annelida: Oligochaeta). Pedobiologia, 20: 411-416.
Direct Link - Curry, J.P. and O. Schmidt, 2007. The feeding ecology of earthworms-a review. Pedobiologia, 50: 463-477.
CrossRef - Dash, H., B.N. Beura and M.C. Dash, 1986. Gut load transit time, gut microflora and turnover of soil, plant and fungal material by some tropical earthworms. Pedobiologia, 29: 13-20.
Direct Link - Davidson, S.K. and D.A. Stahl, 2006. Transmission of Nephridial bacteria of the earthworm Eisenia fetida. Applied Environ. Microbiol., 72: 769-775.
CrossRef - Desjardins, T., F. Charpentier, B. Pashanasi, A. Pando-Bahuon, P. Lavelle and Mariotti, 2003. Effects of earthworm inoculation on soil organic matter dynamics of cultivated ultisol. Pedobiologia, 47: 835-841.
CrossRef - De Lima-Ramos M.R., A.S. Paulino De, A.M. André Goms, N.A.R. Peixoto and S.D.V.M. Tenório, 2004. Importância de bactérias promotoras de crescimento e de biocontrole de doenças de plantas para uma agricultura sustentável. Anais da Academia Pernambucana de Ciência Agronômica, Recife, 1: 89-111.
Direct Link - Espinosa-Victoria D., L. Hernández-Flores and L. López-Reyes, 2006. Diversidad genética de Azosperillum brasilense en suelos cultivados con maíz bajo labranza convencional y de conservación. Terra Latinoamericana, 24: 215-223.
Direct Link - Ettema, C.H. and D.A. Wardle, 2002. Spatial soil ecology. Trends Ecol. Evol., 17: 177-183.
CrossRefDirect Link - Fernández L.A., P. Zalba, M.A. Gómez and M.A. Sagardoy, 2005. Bacterias solubilizadoras de fosfato inorgánico aisladas de suelos de la región sojera. Cienc. Suelo, 23: 31-37.
Direct Link - Fischer, K., H. Dittmar, H. Wolfgang and J. Zeyer, 1997. Effect of passage through the gut of the earthworm Lumbricus terrestris L. on Bacillus megaterium studied by whole cell hydridization. Soil Biol. Biochem., 29: 1149-1152.
CrossRef - Fischer, K., D. Hahn, R.I. Amann, O. Daniel and J. Zeyer, 1995. In situ analysis of the bacterial community in the gut of of the earthworm Lumbricus terrestris L. by whole-cell hybridization. Can. J. Microbiol., 41: 666-673.
Direct Link - Fragoso, C., 2001. Las lombrices de tierra de México (Annelida, Oligochaeta): Diversidad, ecología y manejo. Acta Zool. Mex.(n.s.) Número Especial., 1: 131-171.
Direct Link - Fraser, P.M.M.H., R.C. Baera, T.B. Harrison-Kirk and J.E. Piercy, 2003. Interaction between earthworms (Aporrectodea caliginosa), plants and crop residues for restoring propierties of a degraded arable soil. Pedobiologia, 47: 870-876.
CrossRef - Furlong, M., D. Singleton, D. Coleman and W. Whitman, 2002. Molecular and culture-based analyses of prokaryotic communities from an agricultural soil and the burrows and cast of the earthworm lumbricus rubellus. Applied Environ. Microbiol., 68: 1265-1279.
CrossRef - Garcia J.A. and C. Fragoso, 2003. Influencia of different food substrates on growth and reproduction of two tropical eartworm species (Pontoscolex corethrurus and Amynthas corticis). Pedobiologia, 47: 754-763.
CrossRef - Hernández-García, M.A. and D. Granados-Sánchez, 2006. El parque nacional Iztaccíhualtl-Popocatépetl-Zoquiapan y el impacto ecológico-social de su deterioro. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 12: 101-109.
Direct Link - Homa, J., M. Niklinska and B. Plytycz, 2003. Effect heavy metals on coelomocytes of the earthworm Allolobophora chlorotica. Pedobiologia, 47: 640-645.
CrossRef - Horn, M.A., A. Schramm and H.L. Drake, 2003. The earthworm gut: An ideal habitat for ingested N2O-producing microorganisms. Applied Environ. Microbiol., 69: 1662-1669.
CrossRef - Huerta, E., J. Rodriguez-Olan, I. Evia-Castillo, E. Montejo-Meneses, M. De la Cruz-Mondragón and R. García-Hernández, 2005. La diversidad de lombrices de tierra (Annelida, Oligochaeta) en el estado de tabasco, México. Universidad y Ciencia, 21: 73-83.
Direct Link - Huhta, V., 2002. Soil macroarthtropod communities in planted birch stands in comparison with natural forests in central Finland. Applied Soil Ecol., 20: 199-209.
CrossRef - Hyun-Jung, K., S. Kwang-Hee, C.H. Chang-Jun and H. Hor-Gil, 2004. Analysis of aerobic and culturable bacterial community structures in earthworn (Eisenia fetida) intestine. Agric. Chem. Biotechnol., 47: 137-142.
Direct Link - Jolly, J.M., H.M. Lappin-Scott, J.M. Anderson and C.D. Clegg, 1993. Scanning electron microscopy of two earthworms: Lumbricus terrestris and Octolasion cyaneum. Microb. Ecol., 26: 235-245.
Direct Link - Krisrtuek, V., K. Ravasz and V. Pizl, 1992. Changes in densities of bacteria and microfungi during gut transit in Lumbricus rubellus and Aporrectodea caliginosa (Oligochaeta: Lumbricidae). Soil Biol. Biochem., 24: 1499-1500.
Direct Link - Krištůfek, V., K. Ravasz and V. Pizl, 1993. Actinomycete communities in earthworm guts and surrounding soil. Pedobiologia, 37: 379-384.
Direct Link - Krištůfek, V., K. Ravasz and V. Pizl, 1994. Ultrastructural analysis of earthworm Lumbricus rubellus Hoff. (Annellida, Lumbricidae). Acta Microbiol. Immunol. Hung., 41: 283-290.
Direct Link - Lavelle, P., T. Decaens, M. Aubert, S. Barot and M. Blouin et al., 2006. Soil invertebrates and ecosystem services. Eur. J. Soil. Biol., 42: S3-S15.
CrossRef - Loreno-Osti C., L. López-Reyes and D. Espinosa-Victoria, 2004. Bacterias promotoras del crecimiento vegetal asociadas con gramíneas: Una revisión. Terra Latinoamericana, 22: 225-239.
Direct Link - Márialigeti, K., 1979. On the community-structure of the gut-microbiota of Eisenia lucens (Annelida, Oligochaeta). Pedobiologia, 19: 213-220.
Direct Link - Martínez-Romero, E., 2001. Poblaciones de Rhizobia nativas de México. Acta Zool. Mex. (n.s.) Número Especial., 1: 29-38.
Direct Link - Mendez, R., S. Borges and C. Betancourt, 2003. A microscopical view f the intestine of Onychochaeta borincana (Oligochaeta: Glossoscolecidae). Pedobiologia, 47: 900-903.
CrossRef - Nogales, B., 2005. La microbiología del suelo en la era de la biología molecular: Descubriendo la punta del iceberg. Ecosistemas, 14: 41-51.
Direct Link - Nunan, N., K. Wu, I.M. Young, J.W. Crawford and K. Ritz, 2003. Spatial distribution of bacterial communities and their relationships with the micro-structure of soil. FEMS Microbiol. Ecol., 44: 203-215.
CrossRef - Parle, J.N., 1963. Micro-organisms in the intestines of earthworms. J. Gen. Microbiol., 31: 1-11.
Direct Link - Polyanskaya, L.M., N.I. Babkina, G.M. Zenova and D.G. Zvyagintsev, 1996. Fate of actinomycetes in the intestinal tract of soil invertebrates fed on Streptomycete spores. Microbiology, 65: 560-565.
Direct Link - Rossi J.P., E. Huerta, C. Fragoso and P. Lavelle, 2006. Soil properties inside earthworm patches and gaps in a tropical grassland (la Mancha, Veracruz, Mexico). Eur. J. Soil Biol., 42: S284-S288.
CrossRef - Singleton, D.R., P.F. Hendrix, D.C. Coleman and W.B. Whitman, 2003. Identification of uncultured bacteria tightly associated with the intestine of the earthworm Lumbricus rubellus (Lumbricidae; Oligochaeta). Soil Biol. Biochem., 35: 1547-1555.
CrossRefDirect Link - Spurgeon, D., J.M. Weeks and C.A.M. Gestel, 2003. A summary of eleven years progress in earthworm ecotoxicology. Pedobiologia, 47: 588-606.
CrossRef - Valle-Molinares, R., S. Borges and C. Rios-Velazquez, 2007. Characterization of possible symbionts in Onychochaeta borincana (Annelida: Glossoscolecidae). Eur. J. Soil Biol., 43: S14-S18.
CrossRefDirect Link - Winding, A., R. Rohn and N.B. Hendriksen, 1997. Bacteria and protozoa in soil microhabitats as affected by earthworms. Biol. Fertil. Soils, 24: 133-140.
Direct Link - Young, J.M., L.D. Kuykendall, E. Martinez-Romero, A. Kerr and H. Sawada, 2001. A revision of Rhizobium Frank 1889, with an emended description of the genus and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undica de Lajudie et al., 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. Rubi, R. undicola and R. vitis. Int. J. Syst. Evol. Microbiol., 51: 89-103.
Direct Link