
H.J. Makun
National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Zaria, Nigeria
J.O. Ajanusi
Department of Veterinary Parasitology, Faculty of Veterinary Medicine, Ahmadu Bello University, Zaria, Nigeria
C.A.M. Lakpini
National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Zaria, Nigeria
O.W. Ehoche
National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Zaria, Nigeria
P.I. Rekwot
National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Zaria, Nigeria
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 4 | Page No.: 753-759







ABSTRACT
The study was conducted to determine the relative resistance of Red Sokoto and Sahelian goats to experimental infection with infective larva (L3) of Haemonchus contortus. At the age of about 7-9 months weaned goats were transferred to individual feeding pens and fed concentrate at 08:00 h and given Digitaria smutsii hay ad libitum. Fifteen goats of each breed were divided into 3 groups of five animals each. Goats were either given 0 L3 kg-1 (treatment 1), 75 L3 kg-1 (treatment 2) or 100 L3 kg-1 (treatment 3), three times weekly for 3 weeks. The faecal egg count expressed in Eggs per Gram (EPG) was significantly higher (p < 0.05) for the Sahelian (1477±153.4) than the Red Sokoto (607±147.5). The PCV didn`t show any significant difference between breed however the difference between treatment was significant at p < 0.05. The average daily gain was higher for the Red Sokoto (11.2 g day-1) than the Sahelian (5.6 g day-1) even though the difference was not significant (p > 0.05). The mean total protein was statistically different between breeds. The Red Sokoto had higher (p < 0.05) than the Sahelian (64.6 ± 1.81 vs 57.6 ± 1.86). The mean albumin and globulin concentration did not differ significantly between breed. It was concluded that the Red Sokoto were relatively more resistant to Haemonchus contortus on the basis of faecal egg count, weight gains, changes in the PCV and total serum protein concentration.
PDF Abstract XML References Citation
How to cite this article
H.J. Makun, J.O. Ajanusi, C.A.M. Lakpini, O.W. Ehoche and P.I. Rekwot, 2008. Response of Red Sokoto and Sahelian Goats to Trickle Haemonchus contortus Infection. Journal of Biological Sciences, 8: 753-759.
DOI: 10.3923/jbs.2008.753.759
URL: https://scialert.net/abstract/?doi=jbs.2008.753.759
DOI: 10.3923/jbs.2008.753.759
URL: https://scialert.net/abstract/?doi=jbs.2008.753.759
INTRODUCTION
The population of goats and sheep in Nigeria has been estimated to be about 34.5 and 22.1 million, respectively (Bourn et al., 1994) and they are predominantly made up of the Red Sokoto (Maradi), West African Dwarf and the Sahelian breeds of goats (Blench, 1999; Malau-Aduli et al., 2003). In combination with sheep, small ruminants contribute 17 and 12% meat and milk consumed in sub-Saharan Africa (Lebbie, 2004). The Sahelian is large, long-legged goats mainly found in the semi-arid and arid regions of Nigeria, while the Red Sokoto is more cosmopolitan in distribution. Average flock size of small holder farmers is between 3-5 goats, (Jaitner et al., 2001; Makun et al., 2006). In West Africa traditional goat production is characterized by free roaming in the day and tethering at night or during cropping season (Ogebe et al., 2000). In many places in Nigeria and elsewhere in West Africa goats managed in traditional systems scavenge on road side forage, unimproved rangeland and are minimally supplemented with kitchen waste.
The most common diseases affecting the productivity of goats are pneumonia and helminthiasis, with Haemonchus contortus as the most prevalent genus encountered in the field (Fakae et al., 1999; Nwosu et al., 1996). Helminthosis in small ruminants and goats in particular, is of considerable importance because it affects production through losses due to mortality and reduced weight gain (Tembely et al., 1996). In Nigeria the losses due to helminthosis was estimated at 14 million Naira (N) annually, at a time in the 1970 and early 1980s when goats and sheep were sold at N6 and N4 per head respectively (Okon, 1998). There is therefore a need to obtain the current economic significance of the disease since the price of sheep and goats has now risen by 1000%.
The most important nematodes responsible for economic losses in Nigeria include species of Haemonchus, Bunostomum, Cooperia, Gaigeria,Oesophagostomum, Trichostrongylus and Trichuris. Amongst these nematodes, Haemonchus contortus has the highest prevalence (Ikeme, 1997). Previously helminth control involved the use of anthelmintics and pasture rotation to minimize the devastating effect of the parasite. In recent times, resistance and consumer concern for drug residue is influencing alternative strategies to control these parasites (Athanasiadou et al., 2008; Ogeda-Robertas et al., 2008; Chandrawathani et al., 2003; Emery et al., 2000; Borgsteede et al., 1998).
This trial was designed to investigate the susceptibility or otherwise of the Red Sokoto and Sahelian breeds of goats with respect to Packed Cell Volume (PCV), faecal egg count, Average Daily Gain (ADG) and total serum proteins.
MATERIALS AND METHODS
Location: The study was conducted in September of 2006, at the National Animal Production Research Institute, (NAPRI), Shika, Ahmadu Bello University, Zaria, Nigeria. Shika is situated in the Northern Guinea Savannah between latitudes 11° and 12° N and between longitudes 7° and 8° E; at an altitude of 650 m; with a mean annual rainfall of 1150 mm. The climate of the site is sub humid; rainfall is well distributed during the rainy season between May and October, with about 70% of the rain occurring during the months of July and August. The average temperature and humidity during the wet season are 24.7 °C and 72%, respectively. The early dry season commences from October with a period of cold dry weather known as harmattan lasting until February. This is followed by the hot weather when temperatures fluctuate during day (14-34 °C) and relative humidity is between 10-20%.
Experimental animals and design: Fifteen Red Sokoto and Sahelian goats, aged 7-9 months were obtained from the research farm of NAPRI. The health management of the goats in NAPRI comprises of an annual immunization with pestes des petit (PPR) vaccine purchased from the Nigerian Veterinary Research Institute Vom, ( which is about 200 km southwest of Zaria) weekly dipping to control ectoparasites and strategic deworming to reduce helminth burden. Goats were transferred in groups to concrete floor and acclimatized for 2 weeks during which faecal and blood samples were obtained for gastrointestinal and haemoparasitic examination. Animals were stratified according to breed and body weight and randomly allocated to 3 groups of 5 animals each. The feed consisted of Digitaria simutsii hay and concentrate, which included maize (26.1%); wheat offal (27.8%); cottonseed cake (43.1%); bone meal (2%) and salt (1%) and the goats were fed at 4% of their body weight. Table 1 shows the treatment groups of the animals based on breed. Animals in treatment 1 were the controls and they had no larva infection; those in treatment 2 had 75 larvae (L3 kg-1) live weight given as trickle infection 3 times per week for 3 weeks. While kids in treatment 3 had 100 L3 kg-1 live weight 3 times a week for 3 weeks. (Romjali et al., 1996; Wallace et al., 1996). Water and mineral salts were supplied ad libitum.
| Table 1: | Experimental design of the studies |
 | |
Laboratory analysis: Infective larvae of H. contortus were produced from female worms obtained from abomasums purchased from small ruminant slaughter slabs in 3 market district of northern Nigeria. Abomasums were transported to the laboratory in a cooler and then washed immediately, worms were gently crushed to rupture the uteri and release their eggs. Eggs were cultured at 28 °C in damp heat-sterilized bovine faeces for 7 days. The harvested larvae were counted and stored in water at 4 °C. Thereafter goats were dosed according to their treatment group (Table 1) and then flushed with water.
Parasitological techniques: From time of infection, goats were weighed each week for 14 weeks. Similarly faecal samples collected from the rectum and blood for analysis. Blood samples with or without anticoagulant (EDTA) were collected by jugular venipuncture. Packed cell volume was determined by microhaemotocrit method, while the serum was harvested from clotted blood and total serum proteins were determined by the bieuret method using standard boehring diagnostic kit described by Weichselbaum (1946). The serum albumin fraction was determined by the bromocresol green methods of the boechringer standard diagnostic kit. Globulin fraction was estimated by difference between the total protein and albumin fractions (Nnadi et al., 2007). Faecal samples were collected from the rectum of individual goat for Faecal Egg Count (FEC) using modified McMaster technique.
Analysis was made of the weight changes, intake and digestibility. The differences in faecal egg count between the breeds were analyzed by repeated measures of variance using the general linear model procedure of SAS (2000) package.
RESULTS
The mean weight during the entire duration of the experiment was influenced significantly by breed (p < 0.05). The mean weight for the Sahelian was 12.8 ± 0.83 kg which was higher than the Red Sokoto goats (9.7 ± 0.83 kg). Also the mean weight for the different treatment group varied significantly based on the level of infection, (p < 0.05). When the mean weight of the kids was analysed for the whole duration of the experiment, the control goats had the highest mean weight of 12.1 ± 0.98 kg while low and high levels of infection had mean levels of 11.3 ± 0.98 and 10.6 ± 1.10 kg, respectively.
| Table 2: | Least square means of epg, PCV, total protein, albumin, globulin and weight changes of Sahelian and Red Sokoto goats infected with 3 levels of H. contortus |
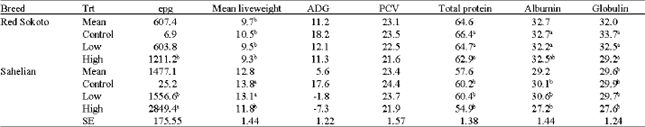 | |
| ab: Means in the same column bearing different superscript letter(s) different significantly (p < 0.05), Trt: Treatment groups, Epg: eggs per gram, ADG: Average Daily Gain, PCV: Packed Cell Volume | |
Average daily gain was significantly higher for the control than for the other 2 infection levels (Table 2). The Red Sokoto had a positive ADG in all 3 infection levels (18.2, 12.1 and 11.3 g day-1) in comparison to the Sahelian whose ADG was 17.6, -1.8 and -7.3 g day-1 for the control, low and high infected groups.
The mean faecal egg count was significantly affected by breed and by treatment as shown in Table 2. The almost absence of eggs in the faeces in the non-infective control goats and the presence of helminth eggs in the infected groups is an evidence of the effectiveness of maintaining the infection throughout the experiment. The Sahelian had significantly (p < 0.05) higher (1477 ± 153.4) mean epg than the Red Sokoto goats (607 ± 147.5). Faecal eggs were first detected from the Sahelian, 3 weeks Post Infection (PI) and the values rose progressively to about 3500 epg by week 8 before dropping to about 2000 epg in week 10 (Fig. 1). There was a slight rise by week 12, after which the dropped continued to the end of the experiment. Meanwhile eggs were detected in the Red Sokoto 4 weeks post infection, slowly rising to a peak of about 1500 epg at week 12 after which it continued to drop until the end of the experiment (Fig. 1).
Table 2 also showed the mean PCV, which did not significantly differ (p > 0.05) between breed but varied significantly between treatment (p < 0.05). In general the PCV started to drop one week post-infection reaching its lowest level of 20% in 4th week post infection for the Red Sokoto and for the Sahelian the lowest level of 20% was obtained at the 6th week post infection, thereafter the PCV steadily increased but never attained the mean pre-infection level. The drop in PCV was also noticed in all treatment groups, for the control group with an initial mean PCV of 27%, the recorded PCV was 24% at the end of the study showing a difference of 3%. The corresponding drop in PCV values for the low and high level infection group were 8.1 and 11.6%, respectively. The initial PCV for the Sahelian and Red Sokoto were 27.5 and 30.5%, the drop to the lowest level was from 28.4 to 20.6 for the Red Sokoto in week 9 and from 30.5 to 19.8% for the Sahelian in the 6th week (Fig. 2).
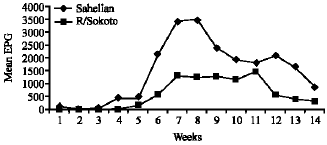 | |
| Fig. 1: | Egg Per Gram (EPG) of feaces recovered from infected Sahelian and Red Sokoto goats |
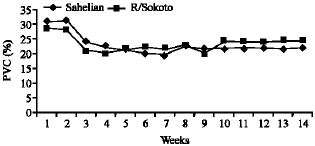 | |
| Fig. 2: | Effect of H. contortus infection on the PVC of Sahelian and Red Sokoto goats |
Thereafter, the PCV increased gradually at different rates in the two breeds, with the Sahelian recording the lower value compared to the Red Sokoto goats, hence the difference between the initial and final PCV were 4.3 and 8.9% for the Red Sokoto and Sahelian breeds, respectively.
The mean total protein is also shown in Table 2, the difference was statistically more significant for breed but not for treatment. Generally there was an increase in the total protein from week 1 post-infection to the end of the study. The Red Sokoto had significantly higher (p < 0.05) 64.6 ± 1.6 g dL-1 than the Sahelian 58.5 ± 1.6 g dL-1), although the initial value for total protein was higher for the Red Sokoto (64.6 g dL-1) than the Sahelian (49.9 g dL-1). The total proteins for the Red Sokoto showed a slight increase from week 1 post infection to week 5 after which it dropped briefly before rising to pre-infective levels. The Sahelian however, continued on the rise from the beginning of the experiment to the end. The difference in total protein between the final and initial were 4.1 and 12.7 for the Red Sokoto and Sahelian goats.
The mean albumin concentration shown in Table 2 showed no significant difference between breed or infection levels. The mean concentration for Red Sokoto and Sahelian breed were 32.7 ± 0.86 and 29.2 ± 0.82 g dL-1, Table 2 also shows the fluctuating pattern of albumin concentration for the 2 breeds post infection, the 2 breeds showed a 50% increase in the albumin concentration week 2 post-infection, thereafter there was an undulating pattern of increment, but the Sahelian had higher albumin at the end of the experiment than at the beginning of the trial. Red Sokoto however had slightly lower concentration at the end of the study than at the beginning. The differences in initial and final mean albumin concentration during the study were -2 and 9 for Red Sokoto and Sahelian breeds.
Similarly the mean globulin concentration is shown in Table 2, indicates no significant difference (p > 0.05) for breed or levels of infection. However, the Red Sokoto had a higher globulin compared to the Sahelian, while globulin concentration decreased with increasing level of infection. Generally, there was a drastic drop in globulin concentration 2 weeks post-infection followed by a gradual increase until the end of the experiment. The recorded difference in the initial and final mean globulin concentrations were 6.1 and 4.6, for Red Sokoto and Sahelian goats.
DISCUSSION
The results of this study showed the difference between the Sahelian and the Red Sokoto in their susceptibility to trickle infection with H. contortus. The delayed establishment of the parasite as indicated by the late appearance of helminth eggs may have been as a result of the high protein in the diet (Nnadi et al., 2007). The Faecal Egg Count (FEC) was higher and more sustained in the Sahelian than in the Red Sokoto. This high and sustained faecal egg count was partly due to the inability to development resistance in the Sahelian (Kahn et al., 2003; Pralomkarn et al., 1997). The research of Baker et al. (1998), found differences in the two indigenous breed of goats, where the small East African goats were more resistant to gastrointestinal nematode infection than the Galla because the later showed significantly higher FEC and lower PCV, with a marked and persistent peri-parturient rise in FEC than the small East African goat. Similar research by Mugambi et al. (1997) showed that sheep also exhibit resistance between breed, they showed that the black-headed Dorper Somalian sheep was more susceptible to H. contortus on the basis of it higher egg count and lower PCV than the Red Masai sheep.
The significantly higher faecal egg count in the present study by the Sahelian breeds of goats was accompanied by a slightly larger difference in the change in PCV values from the start of the experiment and when it terminated. Whereas the change in PCV even though not significant, showed that the drop in the Red Sokoto was from 27 to 21% and that of the Sahelian was from 30 to 21%. This slightly higher drop in PCV, which is an evidence of anaemia demonstrated the fact that the Sahelian were less capable of containing the infection, but for the high protein level of the diet (Abbott et al., 1985) this difference was not statistical.
The initial sudden fall in total protein in the Red Sokoto goats may be due to Hypoproteinaemia; a common feature of helminths infection (Steel et al., 1982), which is sign of haemodilution; a compensatory mechanism for abomasal haemorrhage caused by invading larvae and later large quantities of serum protein lost into the gastrointestinal tract (Sharma et al., 2001). This present study showed that both breeds of goats show increased serum protein level and this research in agreement with those of Mugambi et al. (1996), where total protein increased, but in contrast with that of Abbott et al. (1985) and Haile et al. (2002) where the finn Dorset lambs showed no change in serum protein. This may be due to the higher protein levels fed to the experimental animals. A diet of 16% crude protein CP has been recommended to be adequate for goats, to provide the necessary ingredients for immune response to the infection (Blackburn et al., 1991).
The protein content plays an important role particularly in growing animals like those in this experiment, since protein is very essential for the growth and development of weaned animals (Santra and Karim, 1999) and this diet provided enough dietary protein required to replace that which has been lost due to helminth ingestion and loss into the gastrointestinal tract. In this experiment the animals were post-weaned kids requiring protein for growth and defense against the experimental infection. However, the dietary protein provided (20%) was beyond the recommended (16%) protein in the diet therefore providing adequate nutrition to protect against the devastating effect of the parasite. This is evident by the increase in the total protein and serum globulin recorded during the experiment. It had earlier been suggested by Torres-Acosta et al. (2006), Kahn et al. (2003), Haile et al. (2002) and Kyriazakis et al. (1994) that lambs that were supplemented were able to better withstand the pathogenic effect of infection by maintaining high serum protein and PCV than unsupplemented lambs. Also the increase in globulin concentration may be linked to immune response of the animals to infection, as serum globulin, is a component of the total protein, therefore the increase in the concentrate of the serum globulin observed in the current studies may be linked to the response to the infection. Globulin has been shown to contain immunoglobulin which is necessary for defense against parasitic infection. Abrahams-Sandi et al. (2005), Balic et al. (2002) and Kyriazakis et al. (1996) demonstrated an increase in both cellular and humoral response following helmintic challenge, which go on to suggest the increase the serum globulin in this current study. Therefore, it can be deduced that these responses in this present study may be an immunological response (Gomez-Munoz et al., 1999) against the H. contortus challenge. The results showed that the Red Sokoto goats exhibited dramatic increase in globulin concentration suggesting that their immunological response was more than that exhibited by the Sahelian goats.
It is pertinent to note that the Red Sokoto goats were able to tolerate H. contortus infection because they exhibited high PCV, total protein, serum globulin, weight gain and lower faecal egg count than the Sahelian. Generally the 2 breeds were able to tolerate the infection without any severe weight loss. This is similar to the research of Chiejina et al. (2002), who did not observe any weight loss in the West African Dwarf (WAD) goat when inoculated with between 260-450 L3 kg-1 body weight of the native helminth. The WAD were able to develop protective immunity which was expressed by mild changes in the PCV, little or no effect (Torres-Acosta et al., 2006) on the weight changes, but when in an earlier study they (Fakae et al., 1999) used an exotic strain of the helminth in West African Dwarf goats, their response was severe anaemia and weight loss.
The weight gain in this present trial does not depict the normal weight change for these breeds of goats within this ecological zone. Indeed Makun et al. (2006) recorded a higher average daily gain of 66 and 40 g day-1 for the Red Sokoto and Sahelian kids respectively within the same experimental environment. Therefore the present average daily gain of 11.2 and 5.7 g day-1 is below that for the Red Sokoto and Sahelian respectively fed the same diet, providing a basis for the conclusion that the lower weight gain was due to the deleterious effect of the H. contortus.
It has been reported that following infection with helminths there is usually a 10% reduction in voluntary feed intake and a 30% decrease in live weight gain (Kyriazakis et al., 1994) but the host circumvents this by making selecting between available and appropriate feeds to maintain its requirements. In their study Kyriazakis et al. (1996) showed that even though T. colubriformis parasitized lambs had reduced voluntary feed intake, by selecting their diet between the low, medium and high protein diet, the parasitised lambs were able to consume more protein to obtain similar live weight as the control lambs. It therefore goes to show that since the CP in the present study was above the recommended CP, the goats in the study may not have suffered any lack of protein, which is considered the most limiting ingredient in most livestock diet in developing countries (Devendra, 1999).
The Red Sokoto goats have evolved well within the northern guinea Savanna and this is usually attributed to its survival ability against prevalent disease causing agents within the ecological zone. In this study, the Red Sokoto has demonstrated its ability to be less susceptibility to infection against H. contortus than the Sahelian breeds of goats. This is evident by the lower faecal egg count, higher total proteins, higher average daily gain and minimally changed packed cell volume reported for the Red Sokoto goats compared to the Sahelian goats.
CONCLUSION
The performance of the Red Sokoto was superior to the Sahelian when they were infected with Haemonchus contortus. The Red Sokoto kid were able to relatively resist the infection compared to the Sahelian as indicated by the higher average daily gain, less severe anaemia indicated by higher mean packed cell volume, lower faecal egg count as well as a low worm count in the abomasum. These findings go on to suggest that the Red Sokoto goats were more tolerant and therefore they may be an important breed in breeding and selection using parasite as an indicator.
REFERENCES
- Abbott, E.M., J.J. Parkins and P.H. Holmes, 1985. Influence of dietary protein on parasite establishment and pathogenesis in Finn Dorset and Scottish Blackface Lambs given a single moderate infection of Haemonchus contortus. Res. Vet. Sci., 38: 6-13.
PubMed - Abrahams-Sandi, E., S.M. Geiger, K. Fernandez-Quesada and H. Schulz-Key, 2005. Specific antibody production against different life cycle stages during an experimental A. costaricensis infection in mice Rev. Biomed., 16: 239-246.
Direct Link - Athanasiadou, S., J. Houdijk and I. Kyriazakis, 2008. Exploiting synergisms and interaction in the nutritional approaches to parasitic control in sheep production systems Small Ruminant Res., 76: 2-11.
Direct Link - Baker, R.L., D.M. Mwamachi, J.O. Audho, E.O. Aduda and W. Thorpe, 1998. Resistance of Galla and small East African goats in the sub humid tropics to gastrointestinal nematodes infections and peri-parturient rise in faecal egg counts. Vet. Parasitol., 79: 53-64.
CrossRef - Balic, A., V.M. Bowles and E.N. Meeusen, 2002. Mechanisms of immunity to Haemonchus contortus infection in sheep Parasitic Immunol., 24: 39-46.
Direct Link - Blackburn, H.D., J.L. Rocha, E.P. Figuei-Redo, M.E. Berne, L.S. Viera, A.R. Calvalcante and J.S. Rosa, 1991. Interaction of parasitism and nutrition and their effects on production and clinical parameters in goats. Vet. Parasitol., 40: 99-112.
CrossRefPubMedDirect Link - Bourn, D., W. Wint, R. Blench and E. Woolley, 1994. Nigerian livestock resources survey. World Anim. Rev., 78: 49-58.
Direct Link - Chandrawathani, P., P.J. Waller, M. Adrain and J. Hoglund, 2003. Evolution of high-level, multiple anthelmintic resistance on a sheep farm in Malaysia. Trop. Anim. Health Prod., 35: 17-25.
CrossRef - Chiejina, S.N., B.B. Fakae, J.N. Behnke, P.A. Nnadi, G.A. Musongong and D. Wakelin, 2002. Expression of acquired immunity to a local isolate of Haemonchus contortus by the Nigerian West African Dwarf goat. Vet. Parasitol., 104: 229-242.
CrossRefDirect Link - Devendra, C., 1999. Goat: Challenges for increased productively and improved livelihoods. Outlook Agric., 28: 215-226.
Direct Link - Gomez-Munoz, M.T., M. Cuquerella, L.A. Gomez-Iglesias, S. Mendez, F.J. Fernandez-Perez, C. de la Fuente and J.M. Alunda, 1999. Serum antibody response of Castellana sheep to Haemonchus contortus infection and challenge: Relationship to abomasal worm burdens. Vet. Parasitol., 81: 281-293.
CrossRefPubMed - Haile, A., S. Tembely, D.O. Anindo, E. Mukasa-Mugerwa, J.E.O. Rege, A. Yami and R.L. Baker, 2002. Effects of breed and dietary protein supplementation on the responses to gastrointestinal nematode infections in Ethiopian sheep. Small Ruminant Res., 44: 247-261.
CrossRefDirect Link - Jaitner, J., J. Sowe, E. Secka-Njie and L. Dempfle, 2001. Ownership pattern and management practices of small ruminants in The Gambia-implication for a breeding programme. Small Ruminant Res., 40: 101-108.
Direct Link - Kahn, L.P., M.R. Knox, S.W. Walkden-Brown and J.M. Lea, 2003. Regulations of the resistance of nematode parasites of single and twin bearing Merino ewes through nutrition and genetic selection Vet. Parasitol., 114: 15-31.
Direct Link - Kyriazakis, I., J.D. Oldham, R.L. Coop and F. Jackson, 1994. The effect of subclinical intestinal nematode infection on the diet selection of growing sheep. Br. J. Nutr., 72: 665-677.
PubMedDirect Link - Lebbie, S.H.B., 2004. Goats under household conditions. Small Rumin. Res., 51: 131-136.
CrossRefDirect Link - Malau-Aduli, B.S., L.O. Eduvie, C.A.M. Lakpini and A.E. Malau-Aduli, 2003. Variations in live weight gains, milk yield and composition of Red Sokoto goats fed crop-residue-based supplements in the subhumid zone of Nigeria Livestock Prod. Sci., 83: 63-71.
Direct Link - Makun, H.J., I.R. Mohammad, S.A.S. Olorunju, S.M. Otaru and C.U. Osuhor, 2006. Growth performance of Sahelian and Red Sokoto kids fed Digitaria simutsii hay supplemented with concentrate Nig. Vet. J., 27: 1-7.
Direct Link - Mugambi, J.M., R.K. Bain, S.W. Wanyangu, M.A. Ihiga, J.L. Duncan, M. Murray and M.J. Stear, 1997. Resistance of four breeds of natural and subsequent artificial Haemonchus contortus infection. Vet. Parasitol., 69: 265-273.
Direct Link - Nnadi, P.A., T.N. Kamalu and D.N. Onah, 2007. The effect of dietary protein supplementation of the pathophysiology of Haemonchus contortus infection in West African Dwarf goats. Vet. Parasitol., 148: 256-261.
CrossRefDirect Link - Romjali, E., V.S. Pandey, A. Batubara, R.M. Gatenby and A. Verhulst, 1996. Comparison of resistance of four genotypes of rms to experimental infection with Haemonchus contortus Vet. Parasitol., 65: 127-137.
CrossRef - Santra, A. and S.A. Karim, 1999. Effect of protein levels in creep mixture on nutrient utilization and growth performance of pre-weaner Lambs. Small Ruminant Res., 33: 131-136.
Direct Link - Sharma, D.K., P.P.S. Chauhan and R.D. Agrawal, 2001. Changes in the levels of serum enzymens and total protein during experimental haemonchosis in Barbari goats. Small Ruminant Res., 42: 119-123.
Direct Link - Tembely, S., A. Lahlou-Kassi, S. Sovani, J.E.O. Rege, R.L. Baker and E. Mukasa-Mugerwa, 1996. Small ruminant management practices and control of helminthosis under traditional production systems in the cool Ethiopian highlands. Small ruminant research and development in Africa Proceedings of the 3rd Biennial Conference of the African Small Ruminant Research Network, December 5-9, 1994, UICC, Kampala, Uganda, 5-9 December 1994. International Livestock Research Institute (ILRI), Nairobi, Kenya, pp: 326.
- Torres-Acosta, J.F.J., D.E. Jacobs, A.J. Aguilar-Caballero, C. Sandoval-Castro, L. Cob-Galera and M. May-Martinez, 2006. Improving resilience against natural gastrointestinal nematode infections in browsing kids during the dry season in tropical Mexico Vet. Parasitol., 135: 163-173.
CrossRefDirect Link - Wallace, D.S., K. Bairden, Y.L. Duncan, G. Fishwick, M. Gill, P.H. Holmes, W.A. Mckellar, M. Murray, J.J. Parkins and M. Stear, 1996. Influence of Soya bean meal supplementation on resistance of Scottish blackface lambs to haemonchosis. Res. Vet. Sci., 60: 138-143.
CrossRef