
E.O. Oyekanmi
Department of Crop Protection and Environmental Biology, University of Ibadan, Ibadan, Nigeria
D.L. Coyne
International Institute of Tropical Agriculture, Oyo Road, Ibadan, Nigeria
B. Fawole
Department of Crop Protection and Environmental Biology, University of Ibadan, Ibadan, Nigeria
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 4 | Page No.: 746-752







ABSTRACT
Some selected microorganisms isolated from rhizosphere of crops, were screened for their abilities to enhance crop growth and suppress plant parasitic nematodes; in screenhouse experiments. Two promising hybrids of soybean genotypes: TGx 1448-2E (medium duration) and TGx 1485-1D (early maturing) and a high yielding hybrid of maize genotype Oba Super 1 were used. Microorganisms assessed included the fungi: Trichoderma pseudokoningii, Trichoderma viride, Paecilomyces lilacinus, Aspergillus niger, Glomus mosseae and the rhizobacteria Pseudomonas fluorescens and Pseudomonas putida. One thousand juveniles of Meloidogyne incognita were applied to soybean, while five thousand Pratylenchus zeae (juveniles and adults) were applied to maize. Fungal spores and rhizobacteria cells approximately 2.5x106 mL-1 concentration were applied per plant, for maize experiment. For soybean experiment, Bradyrhizobium japonicum (106 cells), Trichoderma pseudokoningii (6.8x106 spores) and G. mosseae (200 spores) were inoculated per plant. The effect of microorganism`s application was compared with a nematicide (carbofuran) treatment, untreated control and a nematode only control. The parameters measured were nematode density, root damage due to plant parasitic nematode infection, relative leaf chlorophyll content and plant growth parameters. The inoculated beneficial microorganisms in most cases significantly (p≤0.05) reduced the nematode density across the treatments in test crops by up to 79.6% and improved plant growth by up to 46.0%, when treatments performances were compared across treatments. Most of the beneficial microorganisms assessed have prospects of enhancing nematode management through nematode density reduction and improving crop production.
PDF Abstract XML References Citation
How to cite this article
E.O. Oyekanmi, D.L. Coyne and B. Fawole, 2008. Utilization of the Potentials of Selected Microorganisms as Biocontrol and Biofertilizer for Enhanced Crop Improvement. Journal of Biological Sciences, 8: 746-752.
DOI: 10.3923/jbs.2008.746.752
URL: https://scialert.net/abstract/?doi=jbs.2008.746.752
DOI: 10.3923/jbs.2008.746.752
URL: https://scialert.net/abstract/?doi=jbs.2008.746.752
INTRODUCTION
The global call to reduce pesticides residues in food and priority for organically produced agricultural products, make crop scientists` and farmers` interest in organic farming to increase. The use of beneficial microorganisms to reduce plant parasitic nematode damage and/or enhance the growth and yield of crops is an environmental-friendly and healthy option (Holderness et al., 2000; Khan et al., 2002; Oyekanmi et al., 2007). The utilization of nematode-antagonistic fungi and rhizobacteria as Plant Growth Promoting Organisms (PGPOs) and as possible alternatives to synthetic nematicides and inorganic fertilizers have been suggested by Stirling (1991), Khan et al. (2002) and Nwaga et al. (2004). The nematode-antagonistic fungi: Trichoderma pseudokoningii, Trichoderma harzianum, Paecilomyces lilacinus, Dactylaria sp., Aspergillus sp., Verticillium chlamydosporium, may suppress nematodes through predation and/or parasitism (Sikora and Hoffman-Hergarten, 1993; Meyer and Roberts, 2002). Trichoderma spp., for instance, have long been recognised for their potential as biocontrol agents of plant diseases and nematodes are currently being used commercially (Jensen and Wolffhechel, 1995; Kerry, 1998). According to Coyne and Oyekanmi (2007), Bradyrhizobium japonicum enhances nodulation and Symbiotic Nitrogen Fixation (SNF) of soybean in the presence of root knot nematode which reduces nodulation and SNF in most cases and thus improved soybean yield.
The use of such beneficial microorganisms for nematode management and crop improvement is environmentally-friendly and could be an alternative to synthetic nematicides, most of which pose a hazard to the environment. PGPOs such as arbuscular mycorrhizal fungi, Pseudomonas fluorescens, Azotobacter chroococum, Bacillus subtilis solubilise phosphorus and/or nitrogen in the soil and supply them to plant roots in much greater amounts than in their absence (Gaur, 1990; Stirling, 1991; Siddiqui, 2002). Some of these microorganisms affect nematodes through the production of nematodes toxic compounds (Stirling, 1991; Sikora and Hoffman-Hergarten, 1993). Other beneficial microorganisms influence crops physiology which confer immunity on crops against pests through induced resistance responses (Jensen and Wolffhechel, 1995; Oyekanmi et al., 2005).
Most of the pesticides and inorganic fertilizers are not environmentally-friendly, apart from the fact that health hazards may loom as a result of the consumption of their residue in food (Fassuliotis, 1985; Kerry, 1987; Holderness et al., 2000). Also, these agrochemicals are expensive and may not be available for farmers use when needed. Likewise there are legislations against massive use of agrochemical in crop protection, therefore, a search for bionematicide and biofertilizer agents are of global interest. The current study was undertaken to assess the potential of selected microorganisms for nematode suppression and crop improvement.
MATERIALS AND METHODS
Site and experimental details: These experiment were conducted in the screenhouse of the International Institute of Tropical Agriculture (IITA) Ibadan, Nigeria (7°30`N, 3°5`E); a derived savanna agroecology in the year 2005 and 2006. Eight microorganisms isolated from the soil were screened for potential of plant parasitic nematodes management and ability to improve crop growth. The microbes were inoculated separately or in combinations to the soybean plants, while only single inoculation was performed for maize plants. Three kilogram (dry weight) non-sterilized sandy loam soil, with pH 5.98, available N 0.7 g kg-1 and P 2.96 mg kg-1, were used in the screenhouse. Two promising IITA-bred soybean genotypes: TGx 1448-2E (medium duration) and TGx 1485-1D (early maturing) and an IITA-bred high yielding maize genotype, Oba Super 1 were used. Four seeds were planted per pot and thinned to one at seven days after emergence. Pots were maintained in a plastic-covered screen house and irrigated daily. Experimental design was Randomized Complete Block Design (RCBD) with four replications.
Raising and inoculation of nematodes: Nematodes were extracted from roots of Celosia and maize respectively and quantified using standard methodologies (Hooper, 1990). Pure cultures of the nematodes were raised in sterile soil on which tomato and maize were planted respectively for M. incognita and P. zeae. Seven days after planting, soybean seedlings (except in untreated control pots) were inoculated with 1000 M. incognita second stage juveniles (J2) in 2 mL water suspension, the suspension was applied using a micropipette, the suspension was applied to the base of the seedling via two holes 4 cm deep. For maize, 5000 Pratylenchus zeae adults and juveniles, obtained from maize roots were inoculated onto maize seedlings.
Treatments: The seeds used for this experiment were surface-sterilized with 95% ethanol for 3 min, rinsed 7 times with sterile distilled water, prior to inoculation. Yeast mannitol agar was used for the culturing of B. japonicum in the presence of bromothymol blue indicator incubated at 28°C for 2 days. B. japonicum density of 106 cell per mL was used to coat soybean seeds with the aid of boiled starch solution (sticker solution); 5 mL of bacterial suspension and 15 mL of sticker solution were used to coat 60 g of soybean seeds.
In treatments involving G. mosseae, 200 spores/65 mL of soil were inoculated per plant. Fungal spores and rhizobacteria cells approximately 2.5x106 mL-1 concentration were applied per plant, for maize experiment. For soybean experiment, Bradyrhizobium japonicum (106 cells) and Trichoderma pseudokoningii (6.8x106 spores) were inoculated per plant. The pure cultures of the fungi isolated were used at density approximately 2.5x106 mL-1. The organisms were cultured on media; each maize seedling was inoculated with 1 mL of the spore suspension. The fungi (except G. mosseae) were cultured on potato dextrose agar; (Difco Laboratories Detroit, MI, USA) to which 0.5 g streptomycin L-1 of agar medium was added. G. mosseae was cultured in an equal volume of sterile soil and sterile sea sand, on which two maize seedlings were planted. P. fluorescens and P. putida used, were cultured on nutrient agar and bacterial suspensions were made with sterile distilled water. Fungal antagonists` and all bacteria cells were quantified with a haemocytometer. The spores were inoculated at the rate of approximately 2.5x106 spores mL-1 in 5 mL of quantified spore suspension, combined with 15 mL of starch solution as a sticker, per 60 g sterilized seeds. The effectiveness of beneficial microorganisms was compared against nematicide ([carbofuran] Furadan 3G®), applied at the equivalent of 100 kg active ingredient ha-1 incorporated into the soil (0.5 g kg-1 soil).
Nematode density and mycorrhiza colonization assessments: Meloidogyne incognita densities (J2 and adult females) were assessed from a 2 g subsample of each root after staining in hot lactoglycerol with acid fuchsin stain (Bridge et al., 1982). Root galling index was assessed on a scale of 1-5 where: 1 = no galling, 2 = 1-25% of roots with galls, 3 = 26-50% with galls, 4 = 51-75% with galls and 5 ≥75% root galling (Benjamin and Grover, 1987). While Pratylenchus zeae in maize was assessed from 5 g sub sample taken at harvest, using piepan extraction technique (Hooper, 1990). Mycorrhizal colonization of roots was assessed from a 5 g plant-1 root subsample. Roots were cleared with 10% KOH, in an autoclave liquid cycle at 121°C for 15 min. Roots were then rinsed with 5 changes of tap water, bleached with 3% H2O2 rinsed as described above and stained with chlorazol black E stain composition (Brundrett et al., 1994). Colonization of roots was estimated by the grid-line-intersect method (Giovenneti and Mosse, 1980).
Data collection and analysis: Plant height was determined by meter rule, leaf number was visually counted, Leaf Area (LA) was calculated from: length x breadth of leaf x 0.75 for maize and 0.50 for soybean (Alagarswamy and Wani, 1999). Stem girth was measured by venier caliper and Relative Leaf Chlorophyll Content (RLCC) with chlorophyll meter, at weekly intervals. Root weight, root length, cob number (maize only) and nematode density in root were assessed at harvest 16 weeks after planting (WAP). Statistical analysis of data was performed with analysis of variance, using the general linear model procedure on Statistical Analysis Systems Institute (1999). Mean separation was carried out using standard error.
RESULTS
In most cases the inoculated microorganisms significantly (p≤0.05) reduced the nematode density across the treatments and improved crop growth, when treatment performances were compared with nematode control only. Aspergillus treatment had the lowest mean P. zeae density (147.0) that was significantly (p≤0.05) different from the highest (5550.0) density obtained from P. putida treatment (Table 1). P. zeae mean density obtained following Paecilomyces lilacinus (237.5) and T. pseudokoningii (201.8) inoculation were significantly lower when compared with the mean P. zeae density obtained from P. zeae treatments. Maize cob weight and number were not significantly different, between nematode and no nematode treatments, however mean cob weight was highest in Trichoderma viride (3.5 g) and lowest in Paecilomyces sp., treatment (1.9 g). The mean cob weight across the treatments was (2.8 g). Maize cob numbers were higher in both Aspergillus niger and T. pseudokoningii lower cob numbers were obtained in the three treatments: Trichoderma viride, P. fluorescens and P. putida (Table 1).
| Table 1: | Effect of microorganism application on mean Pratylenchus zeae density, Cob weight and Cob number of maize, inoculated with P. zeae in a pot study harvested at 16 WAP. |
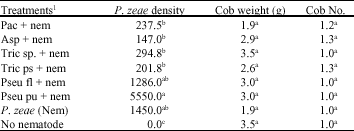 | |
| Means within a column followed by the same superscript letter(s) are not significantly different according to a standard error test (p≤0.05). 1Treatments: Pac = Paecilomyces lilacinus, Asp = Aspergillus niger, Tric sp. = Trichoderma viride, Tric ps = Trichoderma pseudokoningii, Pseu fl = Pseudomonas fluorescens, Pseu pu = Pseudomonas putida, Nem = Nematode (Pratylenchus zeae; 5000 juveniles + adults), No nematode = Control. | |
| Table 2: | Effect of microorganisms application on mean plant height, leaf number and stem girth of maize, inoculated with P. zeae in a pot study harvested at 16 WAP. |
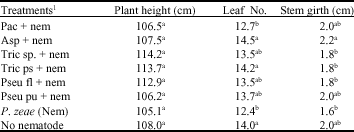 | |
| Means within a column followed by the same superscript letter(s) are not significantly different according to a standard error test (p≤0.05). 1Treatments: Pac = Paecilomyces lilacinus, Asp = Aspergillus niger, Tric sp. = Trichoderma viride. Tric ps = Trichoderma pseudokoningii, Pseu fl = Pseudomonas fluorescens, Pseu pu = Pseudomonas putida, Nem = Nematode (Pratylenchus zeae; 5000 juveniles + adults), No nematode = Control. | |
There was no significant difference in maize height across the treatments at harvest in pots. However, plants in P. zeae treatment were relatively shorter had the shortest height (105.1 cm) and plants in Trichoderma sp., treatment had the highest height (114.2 cm) which was about 9% higher in height than the mean plant height across treatment.
Mean leaf number of maize plants involving nematode treatments and no nematode treatment were significant (p≤0.05) when compared (Table 2). Aspergillus niger, treatment had more mean leaves (14.5) and Paecilomyces lilacinus, plants had lower (12.7). Except for the Paecilomyces sp. plants, all other treatments had their mean leaf number above or very close to (13.7) the mean leaf number across all the treatments. In the no nematode treatment more leaves were obtained which was significantly higher than the nematode control. Stem girth varied across treatments, ranging between 1.6 cm and 2.2 cm. Mean stem girth across treatments was 1.8 cm with the thickest stem girth observed in Aspergillus niger treated plants (1.1 cm).
| Table 3: | Effect of microorganisms application on mean leaf area, relative leaf chlorophyll content (RLCC) 4 and 8 WAP on maize, inoculated with P. zeae in a pot study harvested at 16 WAP. |
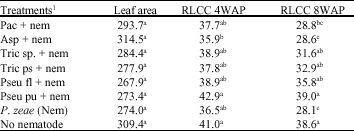 | |
| Means within a column followed by the same superscript letter(s) are not significantly different according to a standard error test (p≤0.05). 1Treatments: Pac = Paecilomyces lilacinus, Asp = Aspergillus niger, Tric sp. = Trichoderma viride. Tric ps = Trichoderma pseudokoningii, Pseu fl = Pseudomonas fluorescens, Pseu pu = Pseudomonas putida, Nem = Nematode (Pratylenchus zeae; 5000 juveniles + adults), No nematode = Control. | |
Relative Leaf Chlorophyll Content (RLCC) of maize plants was significantly different across treatments (Table 3). RLCC at 4 WAP was significant across the treatments, P. putida treatment had the highest RLCC (42.9), while treatments Trichoderma viride and P. fluorescens had values of RLCC slightly higher than the mean (38.7) RLCC across treatments. RLCC at 8WAP was lowest in Aspergillus sp., treatment (28.6) and highest in P. putida (39.0). Treatments Trichoderma sp. and Pseudomonas fluorescens had RLCC above the means RLCC at 8WAP across the treatments. Although Leaf Area (LA) was not significant the highest mean leaf area was obtained in Aspergillus treatment, while the lowest was recorded in Pseudomonas fluorescens treatment. Paecilomyces and Aspergillus niger treatments had leaf area values greater than the mean (285.3) LA across the treatments.
Pratylenchus zeae reduced maize shoot dry weight and root fresh weight by 52 and 90%, respectively across treatments as shown in Fig. 1. However, an increase of 40 and 44%, respectively in maize shoot and root was obtained when microbial antagonistic such as Trichoderma sp. was applied to P. zeae infested pots. Shoot dry weight was significant across the treatments, Paecilomyces lilacinus treatment had the lowest shoot dry weight and, the highest was Trichoderma treatment. Root fresh weight was also significantly different across the treatments. Pseudomonas fluorescens had the lowest root fresh weight and T. pseudokoningii had the highest fresh root weight value. Significant differences were observed when means of shoot dry weight and root fresh weight represented in Fig. 1 was separated by the standard error across the treatments. Meloidogyne incognita density percentage reduction in soybean was significantly reduced across treatments. In most cases, single species of the beneficial soil microbiota reduced M. incognita density across the two genotypes used.
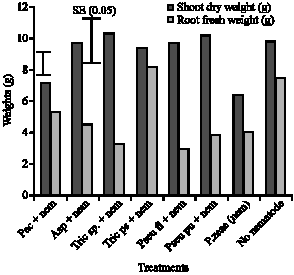 | |
| Fig. 1: | Treatments effect on mean shoot weight and root fresh weight on Oba Super 1 maize genotype in a pot study harvested at 16 WAP. Treatments: Pac = Paecilomyces lilacinus, Asp = Aspergillus niger, Tric sp. = Trichoderma viride, Tric ps = Trichoderma pseudokoningii, Pseu fl = Pseudomonas fluorescens, Pseu pu = Pseudomonas putida, Nem = Nematode (Pratylenchus zeae; 5000 juveniles + adults), No nematode = Control. |
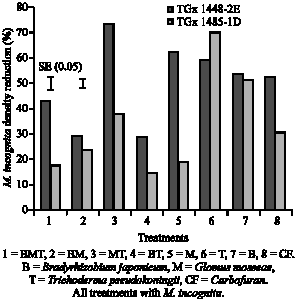 | |
| Fig. 2: | Percentage reduction of M. incognita density in soybean root harvested at eight weeks after planting following various treatments in Meloidogyne incognita infested soil. |
A significantly high reduction was obtained when G. mosseae was combined with T. pseudokoningii. However, this did not lead to better plant growth (Fig. 2).
DISCUSSION
The beneficial microorganisms, T. pseudokoningii and Trichoderma viride, could be antagonistic or nematicidal to P. zeae and root knot nematode, because they suppressed the densities of the nematodes and in some cases improved growth and biomass of the test crops. Paecilomyces sp., could be nematicidal as it further lowered P. zeae density significantly, but in some cases did not improve growth and biomass of crops. The P. fluorescens and Glomus mosseae improved growth of the test crops, without necessarily lowering the density of nematodes significantly. The strain of the P. putida used may not be appropriate organism for biocontrol of P. zeae, as there was no significant reduction (p≤0.05) in P. zeae density but in some instances improved maize growth compared to P. zeae only inoculated.
The use of microorganisms for the management of plant parasitic nematode provides an environmentally safe approach that does not pose any danger to health. Beneficial microorganisms are not harmful to humans, the re-establishment of the soil biodiversity is encouraged and the basis of setting up a preventive biological nematode control program could be achieved (Kerry, 1987; Stirling, 1991; Coyne and Oyekanmi, 2007). Beneficial soil microorganisms do not adversely affect the healthy root system of the plant. The implication of this in biological control and crop improvement as far as end users are concerned is wonderful, because the harvested crops will not contain unacceptable residues. Also, the farmer can bargain on the fact that nematodes cannot develop resistance against antagonistic or nematode parasitic microorganisms. The beneficial soil microorganisms are not subject to the accelerated microbial degradation process in the soil and they are natural inhabitants of the soil they are therefore safe to handle during their application (Kerry, 1987; Meyer and Roberts, 2002).
In the current study, some of the microorganisms were combined together; this was initiated to prevent inconsistencies in effectiveness under varying environmental conditions and investigate best practice of the method of application of the microorganisms against the target pathogen. This was in line with the findings of other work in field of biological pest management (Meyer and Roberts, 2002; Oyekanmi et al., 2007).
There were studies in which no enhanced benefits were observed with combinations of biocontrol agents. For example, the fungi Hirsulella rhossiliensis and Verticillium chlamydosporium, applied against Meloidogyne hapla on lettuce, were effective in reducing J2 numbers in seedlings when applied alone or in combination. These microorganisms could perform well either in single species or as mixed species. However, in most cases, single species application perform better than combination of beneficial microbes in soybean (Gaur, 1990; Rao, 1990; Viaene and Abawi, 2000; Oyekanmi et al., 2007). In this current study the inoculation of G. mosseae and T. pseudokoningii together in combination reduced root knot density in the root of soybean. However, this did not lead to better plant growth. Incompatibility between G. mosseae and T. pseudokoningii could be responsible for the inability of the two organisms to synergistically lead to improved crop growth. On soybean single species of microorganisms performed better in the management of M. incognita and yield enhancement than mixed species of microorganisms (Oyekanmi et al., 2007).
The use of microorganisms in crop production and pest management will have the potential to boost crop yield quality and quantity as there are legislations restricting the use of agrochemicals in the production of crops (Fassuliotis, 1985; Kerry, 1987; Meyer and Roberts, 2002). Most crops form effective mutualistic association with beneficial microorganisms such as indigenous AMF. Studies revealed biomass increase up to 56 and 40% in yield of maize, when AMF was applied to Pratylenchus zeae infested maize plants (Hayman, 1987; Nwaga et al., 2004; Oyekanmi et al., 2005).
Arbuscular mycorrhizal fungi has been shown to reduce the level of damage inflicted by root knot nematodes on plant root systems of Musa spp., which depended on the AMF interaction, in other to enhance crop resistance to Musa nematodes and improve crop growth (Elsen et al., 2003; Fogain and Njifenjou, 2003; Jaizme-Vega et al., 1997). While it is easy to culture and multiply Paecilomyces lilacinus, Trichoderma viride and Pseudomonas fluorescens in large quantity. It might be difficult to produce the promising arbuscular mycorrhiza fungus inoculants in pure culture and use them in Nigeria and in most African countries. This may be due to under development of inoculants technology in the African sub-regions, compared to Europe and America, where inoculation technology are well developed. However, there are organic farming initiatives springing up in Africa and this study will be helpful to them. Such initiatives will help to tackle the challenges of producing the promising mycorrhiza pure inoculant and making them accessible and affordable to resource-poor organic farmers. There is a call to further research into technological development and advancement in order to promote and harness the biodiversity of the beneficial soil microorganisms that perform these ecosystem services of biological nematode management and crop improvement.
In conclusion, from the current study T. pseudokoningii, T. viride, Paecilomyces lilacinus, G. mosseae and Pseudomonas fluorescens have prospects of enhancing nematode management through nematode density reduction and enhanced crop improvement. These selected beneficial microorganisms could be effective alternatives to the toxic nematicides and inorganic fertilizers used in crop production.
ACKNOWLEDGMENT
The authors gratefully acknowledge the financial assistance provided by IITA.
REFERENCES
- Alagarswamy, G. and S.P. Wani, 1999. Plant Observations. In: Methods and Management of Data for Watershed Research: Technical Manual for the Training Workshop, Wani, S.P., P. Singh and P. Pathak (Eds.). ICRISAT Centre, Patancheru, Patancheru, Andhra Pradesh, International Crops Research Institute for the Semi-Arid Tropics, India, pp: 324-502.
- Benjamin, D. and S.C. Jr. Grover, 1987. Biological control of Meloidogyne incognita by Paecilomyces lilacinus and Pasteuria penetrans. J. Nematol., 19: 222-227.
Direct Link - Coyne, D.L. and E.O. Oyekanmi, 2007. Symbiotic nitrogen fixation and nodulation of two soybean genotypes as affected by root knot nematode and microsymbionts. J. Biol. Sci., 7: 1221-1226.
CrossRefDirect Link - Elsen, A., H. Baimey, R. Swennen and D. De Waele, 2003. Relative mycorrhizal dependency and mycorrhiza-nematode interaction in nematode susceptibility. Plant Soil, 256: 303-313.
Direct Link - Fogain, R. and S. Njifenjou, 2003. Effect of a mycorrhizal Glomus sp., on growth of plantain and on the development of Radopholus similis under controlled conditions. Afr. Plant Prot., 9: 27-30.
Direct Link - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Holderness, M., S. Sharrock, E. Frison and M. Kairo, 2000. Organic banana, Towards an organic banana initiative in the Caribbean. In: Report of the International Workshop on the Production and Marketing of Organic Bananas by Smallholders Farmers, October 31-November 4, 1999, International Network for the Improvement of Banana and Plantain, Montpellier France, pp: 173.
- Jaizme-Vega, M.C., P. Tenoury, J. Pinochet and M. Jaumot, 1997. Interaction between the root-knot nematode Meloidogyne incognita and Glomus mosseae in banana. Plant Soil, 196: 27-35.
Direct Link - Kerry, B.R., 1998. Progress towards biological control strategies for plant-parasitic nematodes. The Brighton Crop Protection Conference. Pest Dis., 3: 739-746.
Direct Link - Khan, M.R., K. Kounsar and A. Hamid, 2002. Effect of certain rhizobacteria and antagonistic fungi on root-nodulation and root-knot nematode disease of green gram. Nematol. Medit., 30: 85-89.
Direct Link - Meyer, S.L.F. and D.P. Roberts, 2002. Combinations of biocontrol agents for management of plant-parasitic nematodes and soilborne plant-pathogenic fungi. J. Nematol., 34: 1-8.
Direct Link - Nwaga, D., C. The, R. Ambassa-Kiki, E.L. Ngonkeu Mangaptché and C. Tchiegang-Megueni, 2004. Selection of Arbuscular Mycorrhizal Fungi for Inoculating Maize and Sorghum Grown in Oxisol/Utisol and Vertisol in Cameroon. In: Managing Nutrient Cycles to Sustain Soil Fertility in Sub-Saharan Africa. André Bationo (Ed.). Academy Science Publishers (ASP), Nairobi, pp: 467-486.
- Oyekanmi, E.O., D.L. Coyne, O.E. Fagade and O. Osonubi, 2007. Improving root-knot nematode management of two soybean genotypes through the application of Bradyrhizobium japonicum, Trichoderma pseudokoningii and Glomus mosseae in full factorial combinations. Crop Prot., 26: 1006-1012.
Direct Link - Sikora, R.A. and S. Hoffmann-Hergarten, 1993. Biological Control of Plant-parasitic Nematodes with Plant-Health Promoting Rhizobacteria. In: Pest Management: Biologically Based Technologies, Proceedings of Beltsville Symposium XVIII. Lumsden, R.D. and J.L. Vaughn (Eds.). Washington, DC: America Chemical Society, America, pp: 166-172.
- Vianene, N.M. and G.S. Abawi, 2000. Hirsutella rhossiliensis and Verticillium chlamydosporium as biocontrol agents of the root-knot nematode Meloidogyne hapla on lettuce. J. Nematol., 32: 85-100.
Direct Link