
Mohammad I. Soliman
Department of Zoology, Faculty of Science, Ain Shams University, Abbasia, Cairo, Egypt
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 4 | Page No.: 738-745







ABSTRACT
Ultrastructural changes in Schistosoma mansoni isolated from mice up to 2 weeks after dosing with 50 mg kg1 body weight Ro-354 were investigated. The testicular follicles were completely disorganized. Signs of degeneration of non-germinal cells, vacuolization and swelling of spermatogonia, degeneration of spermatocytes and regression of spermatids were also observed. All stages of spermiogenesis were regressed and developing spermatozoa showed abnormal morphology. As to gastrodermis, a dramatic decrease in the number of surface amplifications (microvilli) was observed. Blebbing, formation of lipid granules, degeneration and prominent flattening of the gastrodermis followed by a general erosion of the tissue to the basal lamina.
PDF Abstract XML References Citation
How to cite this article
Mohammad I. Soliman, 2008. Ultrastructural Alterations in Testis and Gastrodermis of Schistosoma mansoni Due to Treatment of Infected Mice with the New Rhodanine Derivative Ro-354. Journal of Biological Sciences, 8: 738-745.
DOI: 10.3923/jbs.2008.738.745
URL: https://scialert.net/abstract/?doi=jbs.2008.738.745
DOI: 10.3923/jbs.2008.738.745
URL: https://scialert.net/abstract/?doi=jbs.2008.738.745
INTRODUCTION
Studies by Taha and Soliman (2007) have shown that Ro-354 (3-substituted-5- (2-aryl-2-oxoethyl)-2,4-dioxo-1,3-thiazolidine), administered as a single oral dose, has a high antischistosomal activity against Schistosoma mansoni in experimentally infected animals. Diverse biological activities such as anti-inflammatory, antiproteolytic, antibacterial, antiviral and antihelminthic have been found to be associated with thiazolidinone derivatives (Singh et al., 1981).
The use of new antischistosomal drugs is necessary nowadays because many of the present drugs develop drug resistance (Cioli et al., 1995, 2004; Cioli, 1998; Doenhoff et al., 2000; WHO, 1999a, b; Liang et al., 2001; Botros et al., 2005; Mattos et al., 2007). Also reduced cure rates and the failure of treatment after praziquantel (PZQ) treatment have been reported in patients (Fallon et al., 1995; Ismail et al., 1996; Lawn et al., 2003; Magnussen, 2003).
Drug related testis and gastrodermis alterations of schistosomes and other helminthes have received little attention (Bang and Hairston, 1946; Khayyal, 1964; Molokhia and Smith, 1968; Stammers, 1975; Otubanjo, 1981; Irie et al., 1989; Basch and Clemens, 1989; You et al., 1992; Mohamed, 1999; Fawzi et al., 2001).
On the other hand, the schistosome digestive tract has been more or less ignored in such investigatory efforts, although it long has been recognized, because of its role in the physiology and development of the organism, as an ideal system for study (Senft, 1969; Erasmus, 1977). There have been a few reports on drug action relative to this system. For instance, Yarinsky et al. (1970), Bogitsh (1975) and Clarkson and Erasmus (1984) have shown that either in vitro or in vivo administration of the drugs Astiban, Hycanthone, Lucanthone and Niridizole can precipitate morphological changes in the gastrodermis of schistosomes similar to changes resulting from starvation (Bogitsh, 1975).
In the present study, the effect of treatment with 3-substituted-5-(2-aryl-2-oxoethyl)-2,4-dioxo-1,3-thiazolidine (Ro-354) on the ultrastructure of testis and gastrodermis of adult Schistosoma mansoni worms has been examined using transmission electron microscopy.
MATERIALS AND METHODS
Experimental animals and infection: On March 2007, twenty Schistosoma mansoni experimentally infected male mice were supplied by the Schistosome Biological Supply Program (SBSP) at Thoedor Bilharz Research Institute, Giza, Egypt. Thirty days post-infection, mice were divided into two groups of 10 animals. The first group was considered as non-treated infected control. Group 2 was treated intragastrically with 50 mg kg1 body weight of Ro-354 (3-substituted-5- (2-aryl-2-oxoethyl) -2,4- dioxo- 1,3-thiazolidine). Two weeks later, the worms were portally perfused from untreated and drug treated mice by the perfusion technique described by Smithers and Terry (1965).
Transmission electron microscopy: The male worms were carefully cut into small pieces, dropped immediately into 3% phosphate buffered glutaraldehyde (pH 7.3) for 2 h. Tissues were rinsed in 1% buffered osmium tetroxide for 1-2 h at 4 °C, dehydrated in ascending grades of ethanol, embedded in Spurr`s resin. Ultra-thin sections (60-90 nm) were mounted on copper grids and stained with uranyl acetate and lead citrate. Stained grids were then examined by JEOL 1200 EX2-transmission electron microscope (60 kV) at the Central Lab., Faculty of Science, Ain Shams University.
RESULTS
Testicular tissues: Ultrastructural examination of the male reproductive system of schistosomes, revealed that the testicular follicles of S. mansoni are formed of germinal and non-germinal cells bounded by a basal lamina enclosed by a thick coat of circularly arranged muscle fibers. These cells are randomly arranged in the testicular follicle. The developing germ cells appeared electron lucent while the non-germ cells were electron dense. The spermatogonia, which are located peripherally near the basal lamina, possessed oval or rounded nuclei with one large and spherical nucleolus and patches of chromatin bodies; these cells were considerably larger than the other germ cells (Fig. 1).
In treated testis, a significant alterations in testicular structures was noticed everywhere. The muscle layer that surrounds the follicles, increased in size and detached in some places. Also change in direction of muscle fibers and deformation were noticed (Fig. 2).
The non germ cells were the most sensitive testicular component to the drug. These cells exhibited different damage and some of them had completely disintegrated. The cytoplasm became dark and the nucleus disappeared completely, large intercellular spaces were noticed containing debris of degenerating cells. Membranous bodies were also observed (Fig. 2, 3).
The spermatogonia appeared swelling developing very large vacuoles that squeezing nucleus to the cell side. Chromatin patches disappeared and the nucleus became condensed. Lipid granules were observed in the cytoplasm while mitochondria appeared as dark bodies with indistinct cristae (Fig. 4).
Degenerative changes occurred in spermatocytes. A prolonged swelling was observed in cytoplasm and nucleus. The swollen nuclei lost characteristic chromatin patches. Mitochondria exhibited varying morphological changes: some lost their cristae and appeared distended, whereas others contained membrane whorls (Fig. 5). Degenerating spermatocytes sometimes contained membrane whorls which lay directly below the cell surface and around other cytoplasmic organelles (Fig. 5).
Rosette of early spermatids appeared pale with numerous mitochondria accumulated at nuclear anterior end. Lipid granules were observed in the cytoplasm. Nuclei became very pale with very few chromatin patches while some nuclei degenerated completely (Fig. 6).
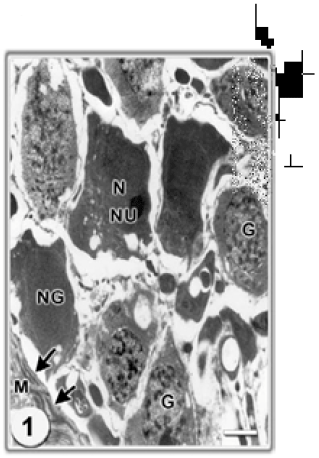 | |
| Fig. 1: | Electron micrograph of a section of testis of S. mansoni collected from control mice showing germinal cells (G), non-germinal cells (NG), nucleus (N), nucleolus (NU), basal lamina (arrows) and circular muscle (M). Bar = 2 μm |
Degeneration and regression of late spermatids were also noted in testicular follicles of treated worms. Cytoplasm develops vacuolization (Fig. 7) while other appears normal in the testicular follicle (Fig. 7).
As to spermatozoa, some of them showed abnormal morphology and their nuclei lost characteristic electron-lucent areas (Fig. 4, 8).
The cytoplasm of parenchyma cells surround the vas deferens swollen. Moreover, plasma membrane of these cells fragmented before the cell completely degenerated. Nuclei swollen and became pale in colour lost their chromatin patches (Fig. 9).
Gastrodermis: The first signs of anomaly appeared as blebbing of small components from the apical surface of the gastrodermis into the lumen and a dramatic decrease in the number of surface amplifications (cytoplasmic extensions) became evident (Fig. 11) as compared to control (Fig. 10). Moreover, an accumulation of many small, Golgi-derived vesicles, a diminution in the Golgi apparatus and the presence of larger, membrane-bound vacuoles, a number of which contained remnants of rough
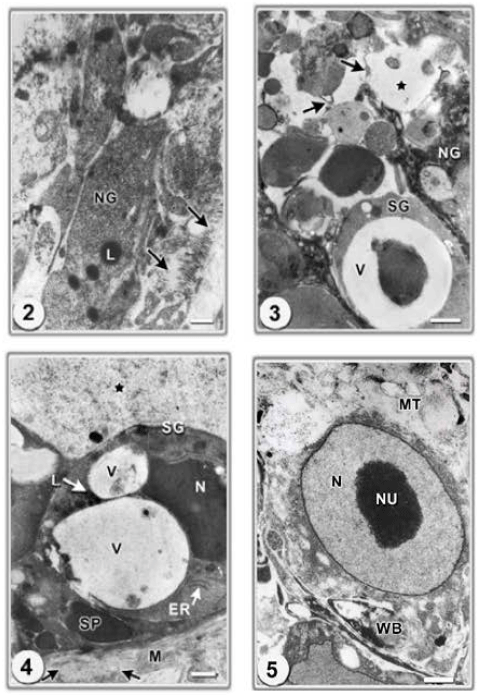 |
| Fig. 2-5: | Electron micrograph of a section of testis of S. mansoni collected from mice treated with 50 mg kg?1 body weight of Ro-354. | |
| 2: | Degeneration of non-germinal cells (NG), aggregation of lipid granules (L). Note also disruption of follicular wall (arrows). Bar = 500 nm | |
| 3: | Showing disorganized testicular tissue. Note swollen spermatogonia (SG) developing pronounced vacuolization (V), non-germinal cell (NG) degeneration leaving a large intercellular spaces (*), membranous materials (arrows). Bar = 1 µm | |
| 4: | Showing abnormal swollen spermatogonia (SG) with large vacuoles (V) that squeezing nucleus (N) sideward, cytoplasm full of lipid granules (L) and fragmented endoplasmic reticulum (ER). Note also large intercellular spaces (*) and regressed spermatozoa (SP). The circular muscles (M) detached from the basal lamina in some places (arrows). Bar = 500 nm | |
| 5: | Showing degenerated spermatocyte with abnormally enlarged nucleus (N) and nucleolus (NU). Note whorled bodies (WB) in degenerated cytoplasm. Mitochondria swollen and lost their cristae (MT). Bar = 1 µm | |
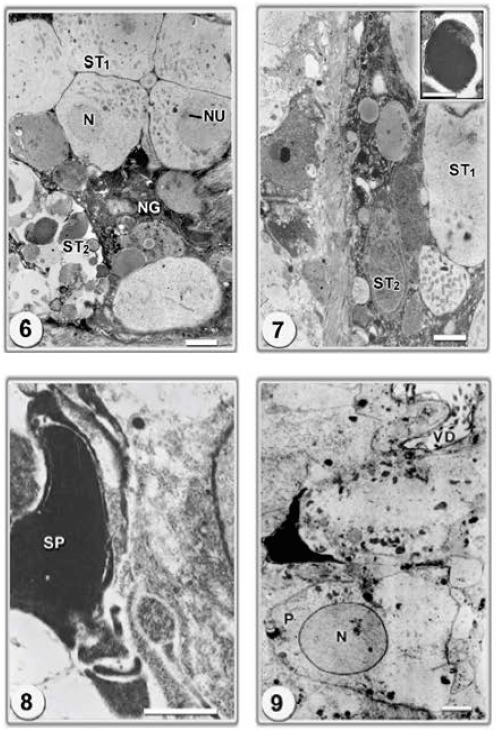 |
| Fig. 6-9: | Electron micrograph of a section of testis of S. mansoni collected from mice treated with 50 mg kg?1 body weight of Ro-354. | |
| 6: | Showing rosette of early spermatids (ST1), with pale cytoplasm containing numerous mitochondria, pale swollen nucleus (N) with nucleolus (NU) and few chromatin patches. Note degenerated non-germinal cells (NG) and late spermatids (ST2) some appeared normal and others degenerated. Bar = 2 µm | |
| 7: | Showing regressed early spermatids (ST1) and late spermatids (ST2). Inset: normal spermatid Bar = 2 m | |
| 8: | Showing spermatozoa (SP) with abnormal morphology and nucleus lost characteristic electron-lucent area. Bar = 1 µm | |
| 9: | Degenerated parenchyma cells (P) surround vas deferens (VD). Note degenerated cytoplasm and swelling nuclei. Bar = 2 µm | |
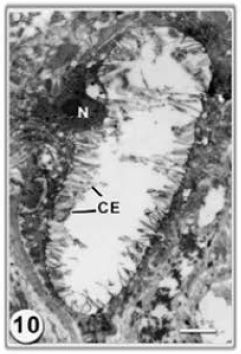 | |
| Fig. 10: | Section of gastrodermis of S. mansoni collected from control mice showing syncytial gastrodermal cells with nucleus (N) and numerous cytoplasmic extensions (CE). Bar = 2 µm |
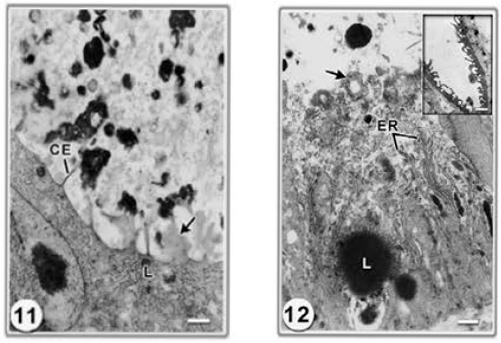 |
| Fig. 11 and 12: | Electron micrographs of sections of gastrodermis of S . mansoni collected from mice treated with 50 mg kg?1 body weight of Ro-354. | |
| 11: | Showing disorganized gastrodermis containing large lipid droplets (L) and blebbing (b) of small components from the apical surface (arrows). Note extensively decreased cytoplasmic extensions (CE). Bar = 500nm | |
| 12: | Another region showing degenerated gastrodermis. Lysis of outer membrane and the released cell content, lipid granules (L), degenerated endoplasmic reticulum (ER). Note membrane bound vacuoles a number of which contained remnants of ER (arrows). Bar = 500 nm inset: showing flattening and erosion of gastrodermis. Bar = 2 µm | |
endoplasmic reticulum causing them to resemble autophagic vacuoles (Fig. 12).
However, the most dramatic changes were twofold. Initially there was a very prominent flattening of the gastrodermis, followed by a general erosion of the tissue to the basal lamina and the gastrodermis was completely degraded (Fig. 12).
DISCUSSION
Testicular tissues: Initially, variability and severity of Ro-354-induced structural changes between different male worms and between different regions of the same worm were seen. Similar observations have been made for S. mansoni treated with praziquantel (Shaw and Erasmus, 1983) and oxamniquine (Popiel and Erasmus, 1984). However, as we saw no evidence that the extracellular space was formed as a result of cytolysis and cell deletion, the drug-induced physiological and morphological changes which gave rise to this bizarre condition is unknown.
Various alterations in testicular structure of S. mansoni have been recorded in the present study as a result of treatment with Ro-354. Such alterations included the disintegration of non-germinal cells, cellular swelling, distortion and disorganization of germ cells, vacuolization within the testis and regression of spermatocytes.
Non-germinal cells were completely disintegrated. Similar observations have been reported in the testis of S. mansoni after treatment with Astiban (Otubanjo, 1981) and Ro15-5458 (Mohamed, 1999). Moreover, vacuolation of the non-germinal cells was reported in testis of S. haematobium treated with Ro15-5458 (Fawzi et al., 2001). On the other hand, non-germinal cells of S. mansoni showed increased phagocytic activity in Astiban-induced testicular damage (Otubanjo, 1981).
In the present study, the germinal cells show swelling and contained very large vacuoles. Nucleus squeezed by these vacuoles. Chromatin patches disappeared and the nucleus became condensed. Cytoplasm developed lipid granules while mitochondria abnormally increased and appeared as dark bodies with indistinct cristae. Leitch and Probert (1984) similarly reported abnormal increase of mitochondria as evident after treatment of S. haematobium with amoscanate. Mohamed (1999) reported shrinkage of germinal cells leads to separation and intercellular spaces after Ro15-5458 administration in case of S. mansoni. Similarly, Fawzi et al. (2001) found that shrinkage and cell separation in S. haematobium. On the other hand, Jiraungkoorskul et al. (2005) reported swelling of the testicular tissue followed by degeneration leaving several hollow spaces in Eurytrema pancreaticum after using PZQ and triclabendazole.
Vacuolation within the testis and regression of spermatocytes observed in the present study are comparable to those recorded by Irie et al. (1989) in their work on S. mansoni treated with PZQ and dextro-PZQ and Mohamed (1999) in S. mansoni treated with Ro15-5458. The anticancer drug procarbazine was found to be profoundly damaging to the primary and secondary spermatocytes and spermatids which were replaced by amorphous granular material (Basch and Clemens, 1989).
Degeneration and atrophy of the testis of S. japonicum was also demonstrated by You et al. (1992) after artemether administration. Otubanjo (1981) mentioned that testicular disorganization was prominent initially in spermatozoa and spermatids but became more generalized with drug accumulation.
Basch and Clemens (1989) reported disruption of meiotic process, spermatocytes and spermatids were destroyed and replaces by amorphous granular material in S. mansoni by procarbazine. The testicular cells were damaged also due to the use of enaminone derivative against S. mansoni (El-Shenawy et al., 2007). The swelling of parenchyma cells found in the present study is similar to that reported by Xiao et al. (2002) after treatment of S. mansoni with artemether.
Gastrodermis: Ultrastructural changes in the caecal cytoplasm following Ro-354 treatment may interfere with digestion and, consequently, have a determental effect on nutrient assimilation. The subsequent degeneration of the gastrodermis may be attributed to further action of Ro-354 on the luminal membrane of the gastrodermis. Howells et al. (1983) attributed to suramin degenerative changes in the gastrodermis of Brugia pahangi which are similar to those observed in S. mansoni in present investigation. The drug may also labializes lysosomal and other membranes, since early investigations (Williamson and MacAdam, 1965) indicate that lysosomal membranes are altered by the drug to a point that acid hydrolases may diffuse out and degrade cytoplasmic organelles, such as ribosomes. The initial effects of Ro-354 on the gastrodermis of S. mansoni are reminiscent of the early stages of starvation induced by various drugs (Yarinsky et al., 1970; Bogitsh, 1975; Clarkson and Erasmus, 1984; Popiel and Erasmus, 1984; Xiao et al., 2002). The increase in number of gastrodermal vacuoles has been documented by Bogitsh (1975) and Clarkson and Erasmus (1984) and is considered to be consistent with changes occurring in the gastrodermis of S. mansoni when subjected to stress conditions, such as starvation or drug treatment, either in vitro or in vivo. Following this initial period, rapid degeneration of the gastrodermis and, eventually, of the surrounding parenchyma occurs. It is not until these last stages of degeneration that changes appear in the tegument, indicating that the drug is most likely affecting the organism from within, e.g., the cecum, while areas such as the tegument display changes reflecting general systemic effects (Xiao et al., 2006; Taha, 2007).
Moreover, focal and extensive lysis, decrease in granular endoplasmic reticulum, vacuolization and degeneration of mitochondria were observed in gut epithelium of S. haematobium harbored in mice following amoscanate (Lietch and Probert, 1984), artemether administration (Xiao et al., 2002, 2006).
Last of all, it would seem that Ro-354 is capable of inducing a range of ultrastructural abnormalities in both the testis and gastrodermal tissues. However, the exact mechanism by which it brings about these changes must await further information from both biochemical and physiological studies.
REFERENCES
- Bang, F.B. and N.G. Hairston, 1946. Studies on Schistosomiasis japonicum. IV. Chemotherapy of experimental Schistosomiasis japonica. Am. J. Hyg., 44: 348-366.
Direct Link - Bogitsh, B.J., 1975. Cytochemistry of gastrodermal autophagy following starvation in Schistosoma mansoni. J. Parasitol., 61: 237-248.
PubMed - Botros, S., H. Sayed, N. Amer, M. El-Ghannam, J.L. Bennett and T.A. Day, 2005. Current status of sensitivity to praziquantel in a focus of potential drug resistance in Egypt. Int. J. Parasitol., 35: 787-791.
CrossRefPubMedDirect Link - Cioli, D., L. Pica-Mattoccia and S. Archer, 1995. Antischistosomal drugs: Past, present and future? Parmacol. Ther., 68: 35-85.
CrossRef - Cioli, D., 2000. Praziquantel: is there real resistance and are there alternartives? Curr. Opin. Inf. Dis., 13: 659-663.
PubMed - El-Shennawy A.M., A.H. Mohamed and M. Abass, 2007. Studies on parasitologic and haematologic activities of an enaminone derivative of 4-hydroxyquinolin-2(1H)-one against murine Schistosomiasis mansoni. Med. Gen. Med., 9: 15-15.
PubMed - Erasmus, D.A., 1977. The host-parasite interface of trematodes. Adv. Parasitol., 15: 201-242.
CrossRef - Fallon, P.G., R.F. Sturrock, A.C. Niang and M.J. Doenhoff, 1995. Short report: Diminished susceptibility to praziquantel in a Senegal isolate of Schistosoma mansoni. Am. J. Trop. Med. Hyg., 53: 61-62.
PubMedDirect Link - Irie, Y., H. Utsunomiya, M. Tanaka, H. Ohmae, T. Nara and K. Yasuraoka, 1989. Schistosoma japonicum and S. mansoni ultrastructural damage in the tegument and reproductive organs after treatment with levo- and dextro-praziquantel. Am. J. Trop. Med. Hyg., 41: 204-211.
PubMed - Ismail, M., A. Metwally, A. Farghally, J. Bruce, L.F. Tao and J.L. Bennett, 1996. Characterization of isolates of Schistosoma mansoni from Egyptian villagers that tolerate high doses of praziquantel. Am. J. Trop. Med. Hyg., 55: 214-218.
PubMed - Jiraungkoorskul, W., S. Sahaphong, P. Sobhon, S. Riengrojpitak and N. Kangwanrangsan, 2005. Effects of praziquantel and artesunate on the tegument of adult Schistosoma mekongi harboured in mice. Parasitol. Int., 54: 177-183.
CrossRefPubMedDirect Link - Khayyal, M.T., 1964. The effects of antimony uptake on the location and pairing of Schistosoma mansoni. Br. J. Pharmacol., 22: 342-348.
PubMed - Lawn, S.D., S.B. Lucas and P.L. Chiodini, 2003. Case report: Schistosoma mansoni infection: Failure of standard treatment with praziquantel in a returned traveller. Trans. R. Soc. Trop. Med. Hyg., 97: 100-101.
PubMed - Leitch, B. and A.J. Probert, 1984. Schistosoma haematobium: Amoscanate and adult worm ultrastructure. Exp. Parasitol., 58: 278-289.
CrossRef - Magnussen, P., 2003. Treatment and re-treatment strategies for schistosomiasis control in different epidemiological settings: A review of 10 years' experiences. Acta Tropica, 86: 243-254.
CrossRef - Mattos, A.C.A., G.C. Pereira, L.K. Jannotti-Passos, J.R. Kusel and P.M.Z. Coelho, 2007. Evaluation of the effect of oxamniquine, praziquantel and a combination of both drugs on the intramolluscan phase of Schistosoma mansoni. Acta Tropica, 102: 84-91.
CrossRef - Mohamed, S.H., 1999. Schistosoma mansoni: Testis alterations induced by antischistosomal drug Ro15-5458. J. Egypt. Soc. Parasitol., 29: 325-334.
PubMed - Molokhia, M.M. and H. Smith, 1968. Antimony uptake by schistosomes. Ann. Trop. Med. Parasitol., 62: 158-163.
CrossRefDirect Link - Otubanjo, O.A., 1981. Schistosoma mansoni: The sustentacular cells of the testis. Parasitology, 82: 125-130.
CrossRef - Popiel, I. and D.A. Erasmus, 1984. Schistosoma mansoni: Ultrastructure of adults from mice treated with oxamniquine. Exp. Parasitol., 58: 254-262.
CrossRef - Senft, A.W., 1969. Considerations of schistosome physiology in the search for anti-bulharziasis drugs. Ann. N.Y. Acad. Sci., 160: 571-592.
CrossRef - Shaw, M.K. and D.A. Erasmus, 1983. Schistosoma mansoni: The effects of a subcurative dose of praziquantel on the ultrastructure of worms in vivo. Z. Parasitenkd, 69: 73-90.
PubMed - Singh, S.P., S.S. Parmar, K. Raman and V.I. Stenberg, 1981. Chemistry and biological activity of thiazolidinones. Chem. Rev., 81: 175-203.
CrossRef - Smithers, S.R. and R.J. Terry, 1965. The infection of laborastory hosts with cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasitology, 55: 695-700.
PubMed - Taha, H.A. and M.I. Soliman, 2007. Antischistosomal activity of 3-substituted-5-(2-aryl-2-oxoethyl)-2,4-dioxo-1,3 thiazolidine (Ro-354). Int. J. Agric. Biol., 9: 87-93.
Direct Link - Xiao, S., B.G. Shen, J. Utzinger, J. Chollet, M. Tanner, 2002. Transmission electron microscopic observations on the ultrastructural damage in juvenile Schistosoma mansoni caused by artemether. Acta Trop., 81: 53-61.
CrossRef - Xiao S., J. Utzinger, B.G. Shen, M. Tanner and J. Chollet, 2006. Ultrastructural alterations of adult Schistosoma haematobium harbored in mice following artemether administration. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi., 24: 321-328.
PubMed - Yarinsky, A.P., P. Hernandez and E.W. Dennis, 1970. The uptake of tritiated hycanthone by male and female Schistosoma mansoni worms and distribution of the drug in plasma and whole blood of mice following a single intramuscular injection. Bull. World Health Org., 42: 445-449.
PubMed - You, J.Q., J.Y. Mei and S.H. Xiao, 1992. Effect of artemether against Schistosoma japonicum. Zhongguo Yao Li Xue Bao., 13: 280-284.
PubMed