
Masoud Tabari
Faculty of Natural Resources and Marine Sciences, Tarbiat Modares University, Noor, Mazandaran, Iran
Mahmoud Asri
Faculty of Natural Resources and Marine Sciences, Tarbiat Modares University, Noor, Mazandaran, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 628-633







ABSTRACT
In order to determine the initial growth and development of direct-seeded Caucasian oak (Quercus castaneifolia C.A. Meyer) seedling an experiment was carried out using the completely randomized design on three soils, of low (soil L), medium (soil M) and high (soil H) nutrient contents in elm-oak stands of the northern Iran. The results at the end of the first growing season demonstrated that values in most of characteristics measured statistically differed among soils. Survival rate, stem length, root length, apical shoot length, node number on the apical shoot, stem dry weight, leaf area and specific leaf area were greatest on soils H. Collar diameter and growth flush number did not differ among different soils. Likewise, nutrient elements in stem, root and leaf were mostly greatest on soil H. Generally, from this research it is deduced that in the elm-oak stands higher fertility soils are more favorable for growth and development of oak seedlings when using seed sowing.
PDF Abstract XML References Citation
How to cite this article
Masoud Tabari and Mahmoud Asri, 2008. Early Growth of Direct-Seeded Quercus castaneifolia (C.A. Meyer) Seedlings on Different Soils of Elm-Oak Stands. Journal of Biological Sciences, 8: 628-633.
DOI: 10.3923/jbs.2008.628.633
URL: https://scialert.net/abstract/?doi=jbs.2008.628.633
DOI: 10.3923/jbs.2008.628.633
URL: https://scialert.net/abstract/?doi=jbs.2008.628.633
INTRODUCTION
Restoration of oak forests is a problem in temperate regions of the northern hemisphere (Lorimer et al., 1994; Ziegenhagen and Kausch, 1995) and the low regeneration success of the oak genus is still a question for ecologists and silviculturists (Lorimer, 1989). Caucasian oak (Quercus castaneifolia C.A. Mey.) is one of main broadleaved species growing throughout the south of Caspian Sea, in length of about 1000 km from coasts to elevations of 2300 m on Elborz mountains. It is favored for its superior form and excellent wood quality. Within its range, Caucasian oak is often found in association with several other hardwood tree species forming stands with multi-storied canopies. It comprises approximately 8% of the standing volume and 7.7% of the trees in the Caspian forests (Sagheb-Talebi et al., 2003). In recent years, a large area of Q. castaneifolia forests has been devastated by humans and livestock and rehabilitation by afforestation and reforestation is necessary. Overall, seed sowing of oaks is preferred to seedling plantation and this important is considered highly, particularly in United States and Europe (Gardiner and Hodges, 1998; Lof and Welander, 2004; Madsen and Lof, 2005). In Denmark and southern Sweden, transplanting of 2-3-year-old bare-rooted seedlings at a stock density of about 2500-5000 per hectare is the most common practice used for afforestation with broadleaves and particularly with oak of former farm land (Henriksen, 1988). It is an expensive method. The cost per transplanted seedling is about 0.4-0.7 Euro including the seedling and the transplanting only. Thus, the development of alternatives is needed. Sowing of oak is an old regeneration technique (Thirgood, 1971) that has lately been revived (Willoughby et al., 1996; Kussner and Wickel, 1998; Ammer et al., 2002). The regeneration cost is one-third to one-half of the cost of transplanting seedlings (Bullard et al., 1992). For 1 Euro it is possible to get about 20-100 pretreated acorns or 100-200 beechnuts. The periodical large crops may result in even lower prices. Consequently, sowing has potential for reaching high stock densities at low costs, resulting in high stand quality.
In Iran seed sowing is implemented mostly in northern (Caspian) forests (Mohadjer, 1998; Jalali et al., 2004) and in western (Zagros) forests (Hesami, 1999) but however, no special research has been still reported with direct seeding of Caucasian oak on different soils. In the current investigation oak acorns are direct seeded in three deforested elm-oak stands with different nutrient available soils to determine how soil influences growth, biomass distribution and nutrient content of Q. castaneifolia seedling during early establishment.
MATERIALS AND METHODS
Study area: The study was conducted in the lowland elm-oak forests of Noor town, north of Iran (Fig. 1). Mean annual precipitation is about 1040 mm and the mean
 | |
| Fig. 1: | Geographical location of the study area in watersheds of northern Iran covered by the Caspian forests |
| Table 1: | Physical and chemical properties of soils in the elm-oak stands |
 | |
| aL, low fertility; M, medium fertility; H, high fertility | |
annual temperature 17 °C. Rainfall distribution is irregular, the maximum occurring in December (211.6 mm) and the minimum in July (19.3 mm). The mean daily temperature in July and August is 25.4 and 23.4 °C, respectively and in January and February 7.4 and 9.6 °C, respectively. The mean maximum temperature of the warmest month is 30 °C and the mean minimum temperature of the coldest month is 3.7 °C (meteorological data referred to the period 1990-2004, Babolsar Station). Based on Emberger classification (1932) climate is moist with mild winters, the dry days (xerothermique index) are 55 and Pluviothermique index (Q2) is 143.6.
Experimental design: In this investigation, Caucasian oak acorns were examined as completely randomized design (replicated three times) in the open fields of three deforested elm-oak stands, each one with distinct soil, such as low fertility (L), medium fertility (M) and high fertility (H) (Table 1). In December 2003 uniform-sized acorns collected from an adjacent middle-aged oak were direct seeded at 5 cm depth in plots 2x2 m (sum of plots = 9 in each stand or on each soil type). The weight of 1000 seeds amounted to 9400 g, the degree of purity was about 100% and the proportion of hollow seeds was 0%. In early spring of 2004 before germination time, herbaceous vegetation was removed in plots. No irrigation was carried out after seedling emergence during the research period.
On all soils on average more than 90% of acorns germinated till April. Near the end of the first growing season (15 November 2004) seedling mortality was noted and all seedlings from each plot were measured for stem length to the nearest cm and root-collar diameter to the nearest mm. Apical shoot length (cm), growth flush number per plant and node number on the apical shoot were also registered and root/shoot ratio (cm/cm) calculated. Three randomly selected seedlings from each plot were chosen to determine root length and stem length. Roots were excavated from the soil by a spade and rinsed free of soil. Main root length (taproot) as well as stem length was recorded by a ruler. Besides, all biomass components were oven-dried at 70 °C to constant weight before weighing with an analytical balance. After drying the biomass components, leaf weight ratio (leaf weight/plant), stem weight ratio (stem weight/plant weight) and root/stem ratio (root weight/stem weight) were calculated for each seedling. Leaf area was measured with a Li-Cor LI-3000 meter and specific leaf weight (leaf biomass/leaf area) calculated. Leaves dry matter of seedling after weighing was averaged regarding to leaves number to obtain the mean dry weight of any leaf. Either of the dried plant components (i.e., leaf, stem and root) after grinding was examined for the determination of nutrient concentration. Nitrogen was determined by the Kjeldahl method, after block digestion using a sulphuric acid-hydrogen peroxide solution. Phosphorus was determined using spectrophotometer and potassium by atomic absorption, following wet digestion of the oven-dried tissue in concentrated HNO3.
Data analysis: Analysis of variance according to a completely randomized design was performed on means of seedling responses calculated in each plot. Significant effects were separated with Duncan`s Multiple Range test (p = 0.05). Means of growth flush number and node number on the apical shoot were analyzed by Mann-Whitney test, following Kruskal-Wallis test. All data analyses were performed using the statistical package of SPSS, version 12.5.
RESULTS
Survival and growth: The survival rate differed among soils. It was lower in soils L and M (40 and 48%, respectively) than in soil H (72%) (Table 2). Collar diameter was not strongly influenced by soil type during the first growing season. It did not exceed 4.5 mm at different soils. Stem length was significantly different among treatments; in soil H it was more than twice as long as (45 cm) those in soils L and M. Taproot responded to soil; it was significantly affected by soil type and was greatest in soil H (26.7 cm) and smallest in soil L (16.0 cm). Apical shoot length was dependent of soil type and was much taller in soil H (13.7 cm) than in soil L (1.4 cm) and soil M (2.5 cm). In reality by the end of period, this term in soil H was approx. five times and ten times greater than those in soils M and L, respectively. Growth flush number was independent of soil fertility. However, each seedling indicated nearly two growth flushes at the end of the first growing season. Seedlings exhibited about 10, 7 and 5 nodes on apical shoot length in soils H, M and L, respectively. Root/shoot ratio varied among the soils and was smallest (0.59) in soil H.
Biomass accumulation: The various soil types significantly affected seedling biomass. Root dry matter, root/shoot ratio and specific leaf area decreased in high fertile soil (H) (Table 3). Root biomass ranged from 1.1 g on high fertility to 2.2 on low fertility; for the same soils, root/shoot ratio ranged from 0.4 to 1.0 g g-1, respectively and specific leaf area from 0.06 to 0.10 mg cm-2. Conversely, leaf dry matter as well as leaf area increased on high nutrient content. Leaf dry matter varied from 0.06 to 0.09 g and leaf area from 6 to 14 cm2, where the bed was low in fertility and high in fertility, respectively. Stem dry matter was greater (2.9 g) in high fertility; it was smaller (1.5 g) in moderate fertility and intermediate (2.1 g) in low fertility.
Nutrient content: Nutrient contents in seedling organs indicated increases with increasing soil fertility (Table 4). N root content ranged from 0.54% in soil M to 84 and 0.97%, respectively in soils L and H. P root content was 0.004% in soils L and M and 0.100% in soil H; for the same
| Table 2: | Average values of Q. castaneifolia seedling survival and growth at different soils of the elm-oak stands |
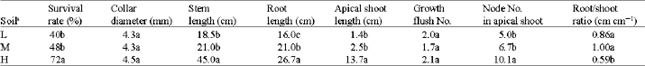 | |
| aFor explanation of abbreviations see Table 1. Within columns, values followed by different letter(s) are significantly different at p<0.05 | |
| Table 3: | Average values of Q. castaneifolia seedling biomass and leaf area at different soils of the elm-oak stands |
 | |
| aFor explanation of abbreviations see Table 1. Within columns, values followed by different letter(s) are significantly different at p<0.05 | |
| Table 4: | Average values of N, P and K percentage of roots, stems and leaves of Q.castaneifolia seedlings at different soils of the elm-oak stands |
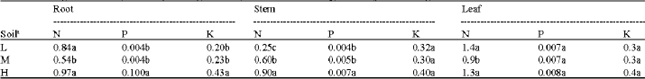 | |
| aFor explanation of abbreviations see Table 1. Within columns, values followed by different letter(s) are significantly different at p<0.05 | |
beds, K root content was 0.20-0.23 and 0.43%. Soil had significant effect on N and P stem content but not on K stem content whereas stem N and P content was increased (0.90 and 0.007%, respectively) in soil H. Only N leaf content was influenced by soil type, values being generally higher in high (1.3%) and low (1.4%) than in medium (0.9%) fertility. This soil effect was not evident for P and K leaf content; however, the values never exceeded 0.008% for P and 0.4% for K in the highest fertility.
DISCUSSION
Seedling survival and growth: In the current study, oak seedling survival was linked to soil status whereas a rather high survival rate was found on high nutrient content (H) and a low-moderate rate on medium (M) and low (L) nutrient contents. In contrast, Kelly (2002, on Q. petraea) and Madsen and Larsen (1997, on Fagus sylvatica) reported negative effect of medium on survival. Generally, effect of nutrient availability on the growth and development of tree seedlings is well known (Jose et al., 2002; Tabari et al., 1999). Although Berger and Glatzel (2001, on Q. petraea) believe that collar diameter increases with increased nutrient availability but in our research no clear difference of this characteristic was detected on sites with different nutrient.
In the present study the total length of the apical shoot was closely linked to soil bed whereas it was higher on high fertile site, a finding also reported by Harmer (1989) for Q. petraea seedlings.
We did not found significant difference with growth flush number at soil treatments. In the best conditions (on soil H) it did not exceed 2 units per seedling. Of course, in natural conditions sessile oak may produce up to four or five growth flushes in a growing season; nevertheless the limiting growing conditions may often confine production to only one or two flushes, thus restricting the full growth potential (Lavarenne-Allery, 1965; Longman and Cutts, 1974). Collet et al. (1997) working on Q. petraea showed that in year 1 or 2, growth flushes did not exceed one or two even where the seedlings were irrigated, but it was counted three to four where the ground vegetation was eliminated. Similarly it expects that in this research growth flushes of Q. castaneifolia would also elevate to four or five if herbal vegetation were removed following the seedling emergence. Our observations revealed that node number in apical shoot reached about 10/growth unit where the seedlings were grown on high fertility without watering. This is while the same result was obtained for 1-year-old Q. petraea seedlings when irrigated and cleaned from ground vegetation (Collet et al., 1997). This implies that this term may also promote two times (20/growth unit) in Caucasian oak seedlings when irrigated and released from weed.
Generally, nutrient deficiency strongly hinders seed germination and development of primary roots (Gehrmann and Ulrich, 1983). Likewise, reduction of root length is often due to the nutrient deficiency (particularly N and P) of soil (Brisette and Chamber, 1992). The confirmation of this matter can be seen in our results such that with decreasing fertility smaller root length (taproot) was produced.
Biomass accumulation: As findings of Van Hees (1997, on Q.robur and F. sylvatica) and Collet et al. (1997, on Q. petraea), in the present study, the superior stem growth of oak seedlings raised on the high nutrient-rich soil (H) was associated to a greater biomass accumulation. In contrast, the reduced stem growth of seedlings receiving lower nutrient content (L and M) was associated to a lower level of biomass accumulation. The likely explanation for the reduced stem growth and low biomass accumulation observed on nutrient-poor soil with sandy texture (L) may involve moisture stress, as it is cited that field capacity is higher in the fine texture than in the coarse texture.
Seedling development in term of root dry matter was indirectly proportional to fertility. This is while that for beech seedling Minotta and Pinazuti (1996) found no significant difference of root biomass in low and high fertilities (under high light environment). In our study, on sandy soil (L), benefited from the low nutrient and moisture, the small taproot and rootlets expanded in superficial layer exhibited greater root biomass. Similarly, Van Hees (1997) proves that seedlings of F. sylvatica and Q. pedunculata produce bigger root biomass where the moisture content is poor. The same finding is observed in working of Walters and Retch (1997) on maple, too.
As finding of Minotta and Pinazuti (1996), in our examination root/shoot biomass decreased on high nutrient site. This is while that both investigations demonstrated bigger leaf dry matter and larger leaf area on such site. The reduced leaf area could be also caused by soil moisture shortage (Van Hees, 1997). In the present review, such an occurrence with leaf area was appeared in soil L being benefited from a coarse texture.
Specific leaf area decreased on high fertile soil. This is in inconsistent with line of Minotta and Pinzauti`s (1996), who revealed this attribute was declined on nutrient-rich soil (under high radiation supply). In literature it looks like that influence of light more than soil has been reported with this term; it often decreases due to the increase of light intensity (Walters and Retch, 1997; Tabari et al., 2005).
Nutrient content: The soil effect was significant for N, P and K content of root. Potassium content of stem and leaf as phosphorus content of leaf seemed to be independent of soil fertility. With increasing fertility, N and P content increased in root and stem. Where the nutrient availability was raised (soil H) K content increased in root but it did not vary in stem and leaf. In total, it can be noted that the richer substrates seem to have promoted the plant`s uptake of N, P and K as the similar result for N and K, not for P, is observed with European beech (Minotta and Pinzauti, 1996). In other words, it is accepted that nutrient concentration in seedlings elevates with increasing soil fertility (Marler and Strom, 2003). Accordingly, Milberg and Lamont (1997) declare that P content of seedlings is higher in high nutrient available soils.
CONCLUSION
In conclusion, the early results of this investigation reveal that in the elm-oak stands most of the characteristics measured, particularly survival and early growth, with Q.castaneifolia seedling sown by acorn are best on high nutrient site. This is while that the growth suitability is produced to some extent on medium nutrient content site (M), too. However it may be better that for rehabilitation of Caspian lowland forests where the elm-oak stands have been devastated since some years ago direct seeding of Q.castaneifolia to be carried out on the higher fertile sites.
REFERENCES
- Ammer, C., R. Mosandl and H. El Kateb, 2002. Direct seeding of beech (Fagus sylvatica L.) in Norway spruce (Picea abies L. Karst.) stands-effect of canopy density and fine root biomass on seed germination. For. Ecol. Mange., 159: 59-72.
Direct Link - Berger, T.W. and G. Glatzel, 2001. Response of Quercus petraea seedlings to N fertilization. For. Ecol. Manage., 149: 1-14.
Direct Link - Brisette, J.C. and J.L. Chamber, 1992. Leaf water status and root system water flux of shortleaf pine (Pinus echinata Mill.) seedlings in relation to new root growth after transplanting. Tree Physiol., 11: 289-303.
CrossRefDirect Link - Bullard, S., J.D. Hodges, R.L. Johnson and T.J. Straka, 1992. Economics of direct seeding and planting for establishing oak stands on old-field sites in the South. South J. Applied For., 16: 34-40.
Direct Link - Collet, C., F. Colin and F. Bernier, 1997. Height growth, shoot elongation and branch development of young Quercus petraea grown under different levels of resource availability. Ann. Sci. For., 54: 65-81.
CrossRefDirect Link - Gardiner, E.S. and J.D. Hodges, 1998. Growth and biomass distribution of cherrybark oak (Quercus pagoda Raf.) seedlings as influenced by light availability. For. Ecol. Mange., 108: 127-134.
CrossRef - Harmer, R., 1989. The effect of mineral nutrients on growth, flushing, apical dominance and branching in Quercus petraea (Matt.) Liebl. Forestry, 62: 383-395.
CrossRefDirect Link - Jose, S., S. Merritt and C.L. Ramsey, 2002. Growth, nutrition, photosynthesis and transpiration responses of longleaf seedling to light, water and nitrogen. For. Ecol. Mange., 180: 335-344.
CrossRef - Kelly, D.L., 2002. The regeneration of Quercus petraea (sessile oak) in southwest Ireland: A 25-years experimental study. For. Ecol. Manage., 166: 207-226.
CrossRefDirect Link - Lof, M. and N.T. Welander, 2004. Influence of herbaceous competitors on early growth in direct seeded Fagus sylvatica L. and Quercus robur L. Ann. For. Sci., 61: 781-788.
Direct Link - Lorimer, C., G.J.W. Chapman and W.D. Lambert, 1994. Tall understorey vegetation as a factor in the poor development of oak seedlings beneath mature stands. J. Ecol., 82: 227-237.
CrossRefDirect Link - Madsen, P. and J.B. Larsen, 1997. Natural regeneration of beech (Fagus sylvatica L.) with respect to canopy density, soil moisture and soil carbon content. For. Ecol. Mange., 97: 95-105.
CrossRef - Madsen, P. and M. Lof, 2005. Reforestation in Southern Scandinavia using direct seeding of oak. Forestry, 78: 55-64.
CrossRef - Marler, R.J. and J.C. Strom, 2001. Growth response of Populus fremontii, Salix goodtingii and Tamarix ramosiissims seedling under different nitrogen and phosphorus concentration. J. Arid Environ., 49: 133-146.
CrossRefDirect Link - Milberg, P. and B.B. Lamont, 1997. Seed cotyledon size and nutrient content play a major role in early performance of species on nutrient-poor soil. New Phytal., 137: 665-672.
Direct Link - Minotta, G. and S. Pinazuti, 1996. Effects of light and soil fertility on growth, leaf chlorophyll content and nutrient use efficiency of beech (Fagus sylvatica L.) seedling. For. Ecol. Manage., 86: 61-71.
CrossRef - Tabari, M., P. Fayaz, K. Espahbodi, J. Staelens and L. Nachtergale, 2005. Response of oriental beech (Fagus orientalis Lipsky) seedlings to canopy gap size. J. For., 78: 443-450.
CrossRefDirect Link - Van Hees, A.F.M., 1997. Growth and morphology of pedunculate oak (Quercus robur L) and beech (Fagus sylvatica L) seedlings in relation to shading and drought. Ann. Sci. For., 54: 9-18.
CrossRefDirect Link - Walters, M.C. and P.B. Retch, 1997. Growth of Acer saccharum seedlings in deeply shaded understories of northern Wisconsin: Effects of nitrogen and water availability. Can. J. For. Res., 27: 237-247.
CrossRefDirect Link - Ziegenhagen, B. and W. Kausch, 1995. Productivity of young shaded oaks (Quercus robur L.) as corresponding to shoot morphology and leaf anatomy. For. Ecol. Mange., 72: 97-108.
CrossRefDirect Link