
Zaidi Che Cob
School of Environmental and Natural Resource Science, Faculty of Science and Technology,
National University of Malaysia, 43600 Bangi, Selangor, Malaysia
Aziz Arshad
Department of Biology, Faculty of Science, Universiti Putra Malaysia,
43400 UPM Serdang, Selangor, Malaysia
Japar Sidik Bujang
Department of Biology, Faculty of Science, Universiti Putra Malaysia,
43400 UPM Serdang, Selangor, Malaysia
Mazlan Abd. Ghaffar
School of Environmental and Natural Resource Science, Faculty of Science and Technology,
National University of Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 616-621







ABSTRACT
The study was conducted at the Merambong Shoal, Johor Straits, Malaysia. Strombus canarium was the most abundant herbivorous mollusc within the study site, highly prized as seafood and contributes to the economics of the locals. Ontogenetic development of sex characters and sexual maturity were determined via logistic curve analysis, using shell-length and/or lip thickness as predictors. The minimum shell-length at which sex characters appeared was at 30 mm in males and 32 mm in females and both sexes can definitely be determined at more than 41 mm shell-length. The SX50 (probability of individuals can be sexed is 0.5) was at 38.33 ± 0.41 mm for male and at 37.15 ± 0.31 mm for female. The SL50 (length at which the probability of individuals are matured is 0.5) was at 54.14 ± 0.86 mm shell-lengths for males and at 58.51 ± 1.02 mm shell-lengths for females. The LIP50 (lip thickness at which the probability of individuals are matured is 0.5) values on the other hand was about 0.69 ± 0.0003 and 0.80 ± 0.014 mm for males and females respectively. The findings indicate that sexual dimorphisms occurred at very early stage in S. canarium life history.
PDF Abstract XML References Citation
How to cite this article
Zaidi Che Cob, Aziz Arshad, Japar Sidik Bujang and Mazlan Abd. Ghaffar, 2008. Sexual Maturity and Sex Determination in Strombus canarium Linnaeus, 1758 (Gastropoda: Strombidae). Journal of Biological Sciences, 8: 616-621.
DOI: 10.3923/jbs.2008.616.621
URL: https://scialert.net/abstract/?doi=jbs.2008.616.621
DOI: 10.3923/jbs.2008.616.621
URL: https://scialert.net/abstract/?doi=jbs.2008.616.621
INTRODUCTION
Strombus canarium, as with other Strombus species, is colonial and abundant wherever it occurs. The species normally associated with sandy mud bottoms and seagrass beds (Abbott, 1960; Erlambang and Siregar, 1995; Zaidi et al., 2005). It was the most abundant herbivorous mollusc within the study site and might contribute to the maintenance and well being of the seagrass bed ecosystem (Zaidi et al., 2005). The species was highly prized as seafood in Malaysia and other areas within the region (Chuang, 1961; Erlambang and Siregar, 1995; Zaidi et al., 2005).
The gleaning activity has been continued for ages and very seasonal in nature. Each year the species appeared in large number during breeding season, which starts from late November to early March (Zaidi et al., 2005; Japar Sidik et al., 2006), which the locals take full advantage by harvesting them. There was currently no regulation in Strombus fishery activity, which prompts the current study. Preliminary surveys in local market found very wide range of conch age was harvested (personal observations), from juvenile to reproductively active females and to very old individuals.
The objective of this study is to determine the minimum shell-length at which they matured and become reproductively active. In addition the minimum length at which sexual character appeared was also investigated. The information is very important for better understanding and better management of this commercially important fishery species.
MATERIALS AND METHODS
Study sites and sample collection: The study was conducted at the Merambong Shoal (01° 19.979’N, 103° 35.965’E) in Johor Straits, Malaysia (Fig. 1), from January to June 2005. It is a subtidal shoal that was only accessible during period of extreme low tide. Sample collections were conducted using transect line approach where all conch within 2 m areas of a 50 m transect line were collected. In laboratory the shells were cleaned and fouling organisms were scrapped-off and the animals were separated from their shell.
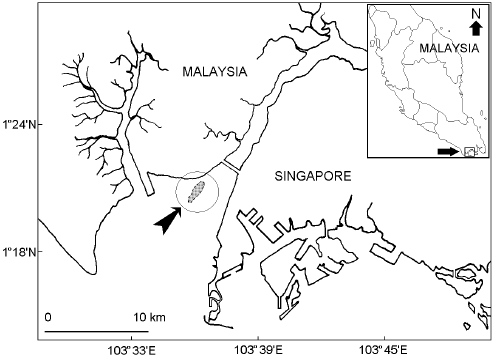 | |
| Fig. 1: | West Johor Straits, Malaysia showing the sampling site i.e., Merambong Shoal (encircle), which was only exposed during extreme low tide period |
Ontogenetic development of sex characters: Shell-length and lip thickness of juvenile of various length sizes were measured and the presence of its external reproductive genitalia i.e. the verge in males and egg groove in females, were noted. Individuals were then group into 1 mm length classes and the proportions of sexed animals against unidentified sex were determined for both sexes. A logistic-curve was then fitted to the data using the software ORIGIN® (Microcal Software Inc.), using the equation of:
(1) |
where, Pi is the probability that individual-i can be sexed. A and B are coefficients describing the intercept and slope of the logistic-curve and Li is independent variable (growth in length) of individual-i. The non-linear curve fitting was conducted using Levenberg-Marquardt least-squares method, incorporated in the ORIGIN® software package. The SX50, which indicated the probability individuals could be sexed is 0.5, was then determined for male and female S. canarium.
Sexual maturity: The juvenile and adult stages of S. canarium are clearly defined through the thickened and flared shell-lip of the latter. However, the onset of maturity and rates at which they matured are still not known. Therefore maturity stages were determined, which were based on: (1) Observation of penis development in males and egg groove in females; (2) Flaring and thickening of the lips; (3) Macroscopic observation on gonad coloration and appearance (e.g., vesicle seminalis in males) under dissecting microscope and (4) Microscopic observation on gonad of maturing or newly matured individuals, using compound microscope.
Maturity determination and analyses were then conducted based on shell-length and shell-lip thickness, two main shell characters that are highly correlated with maturity of this species (Abbott, 1960; Appeldoorn, 1988). The proportion of mature animals in each size class was computed and a logistic-curve was fitted, again using the Eq. 1 but Pi is the probability that individual-i is mature; A and B are coefficients describing the intercept and slope of the curve and Li is growth in length or shell-lip thickness of individual-i. The onset of maturity was then expressed as SL50 and LIP50, which indicated the probability of an individual being mature is 0.5, at the respective Shell-Length (SL) and shell-lip thickness (LIP) values. The non-linear curve fitting was conducted using Levenberg-Marquardt least-squares method.
RESULTS
General anatomy of the gonad: At macroscopic levels, gonad of the adult S. canarium can be differentiated between males and females. Testes and seminal vesicle were clearly visible in males, while the ovary, oviduct and fallopian tube present in females (Fig. 2A, B). However,
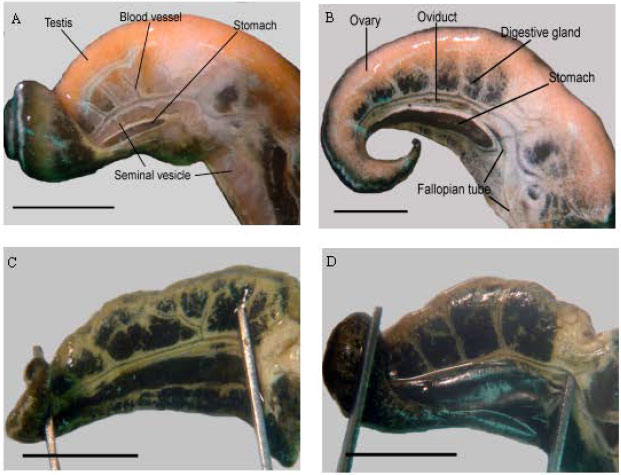 | |
| Fig. 2: | Strombus canarium gonad: (A) immature male; (B) Immature female; (C) Mature male; (D) Mature female. Scale bar = 1 cm |
the male and female gonads were almost identical during juvenile stage (Fig. 2C, D), which appeared as thin, pale-brown tissues overlaying the digestive gland. There was subsequent change in color of the gonad as they matured: from blackish to cream or pale yellow to brownish yellow or bright orange at mature stage.
Ontogenetic development of sex characters: The minimum shell-length that the sex characters appeared was at 30 mm shell-length for male and 32 mm shell-length for female and both sexes can definitely be determined when S. canarium attained more than 41 mm shell-length. Using the logistic curve analysis the SX50 (where the probability individuals can be sexed is 0.5) was at 38.33 ± 0.41 mm (logistic curve analysis: χ2 = 0.011, R2 = 0.94, N = 406) for male and at 37.15 ± 0.31 mm (logistic curve analysis: χ2 = 0.0064, R2 = 0.97, N = 569) for female (Fig. 3, 4, respectively).
The verge of a matured male consists of a grooved stalk with extended prongs at terminal. Important features of a matured S. canarium verge include: auxiliary prongs
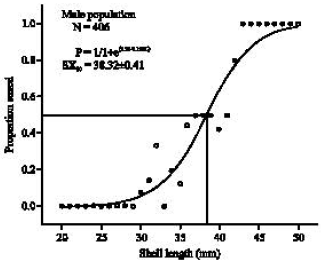 | |
| Fig. 3: | Logistic curve plot of sex determination in male S. canarium. P = probability of successful sex determination; SL = shell-length (mm) |
(anterior and lateral), accessory pad and keel. Figure 5 showed a schematic diagram of a typical adult verge and
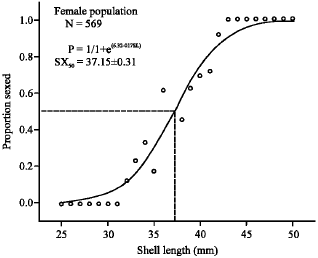 | |
| Fig. 4: | Logistic curve plot of sex determination in female S. canarium. P = probability of successful sex determination; SL = shell-length (mm) |
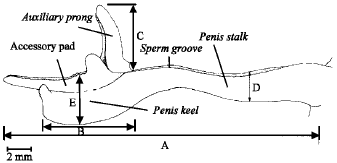 | |
| Fig. 5: | Schematic line drawing of a matured S. canarium verge. A = verge total length; B = keel length; C = auxiliary prong length; D = stalk width; E = keel width |
measurements that were used to investigate variability in the verge structure and to compare with other conch species. For a narcotized (MgCl2) adult, S. canarium verge length ranged from 20.18 to 31.51 mm; stalk width ranged from 2.2 to 3.72 mm; keel length from 7.93 to 13.96; penis keel width from 4.88 to 9.76 mm; accessory pad width from 1.43 to 4.07 mm and the main auxiliary prong length from 3.45 to 7.28 mm (Table 1).
Sexual maturity: Male conch matured at lower shell-length and lower shell-lip thickness compared with females. The SL50 (length at which the probability of individuals are matured is 0.5) for males was 54.14 ± 0.86 mm (logistic-curve analysis: χ2 =0.021, R2 = 0.81, N = 257) lengths (Fig. 6), whilst SL50 for female was 58.51 ± 1.02 mm lengths (logistic-curve analysis: χ2 = 0.028, R2 = 0.74, N = 497) (Fig. 7). The LIP50 value on the other hand was about 0.69 ± 0.0003 mm (logistic-curve analysis:
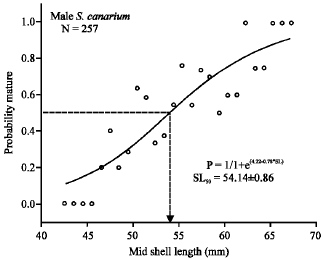 | |
| Fig. 6: | Logistic curve plot of percentage of mature S. canarium males for the respective shell-length class. Abbreviations: P, probability of mature males; SL, shell-length (mm) |
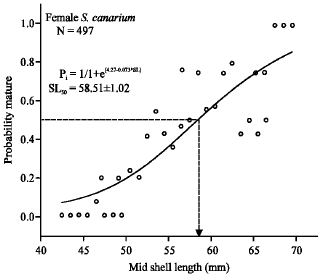 | |
| Fig. 7: | Logistic curve plot of percentage of mature females for the respective shell-length class. Abbreviations: P, probability of mature males; SL, shell-length (mm) |
| Table 1: | Morphometric measurement of verge characters of adult S. canarium |
 | |
χ2 = 0.00002, R2 = 0.99, N = 257) for males (Fig. 8), compared with 0.80 ± 0.014 mm (logistic-curve analysis: χ2 = 0.01, R2 = 0.95, N = 497) in females (Fig. 9)
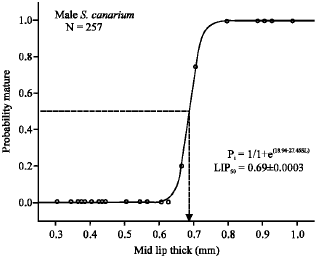 | |
| Fig. 8: | Logistic curve plot of probability of mature males for the respective lip thick class. Abbreviations: P, probability of mature males; LIP, shell-lip thickness (mm). |
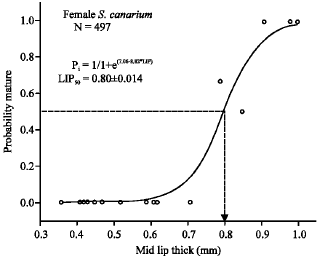 | |
| Fig. 9: | Logistic curve plot of probability of mature females for the respective lip thick class. Abbreviations: P, probability of mature females; LIP, shell-lip thickness (mm) |
DISCUSSION
The general anatomy of S. canarium was strikingly similar to other Strombus species except for the verge structure. For the first time detailed examination has been conducted on the morphology of S. canarium verge. Abbott (1960) only briefly described the verge as simple, with broad swollen distal end. This study however found the verge was rather complex, with few extensions and prongs, which make it unique from that of other Strombus species present within the study area, i.e., Strombus marginatus, Strombus urceus and Strombus vittatus
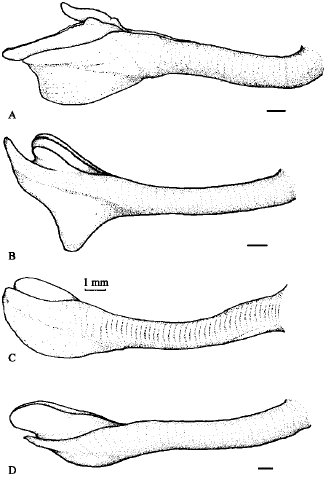 | |
| Fig. 10: | Schematic line drawing of Strombus verge, drawn from fresh specimen taken from Merambong Shoal: A. S. canarium; B. S. marginatus; C. S. urceus; D. S. vittatus. Scale bar = 1 mm |
(Fig. 10). Important characters that differentiate S. canarium verge from other Strombus species were the presence of an elongated auxiliary prong, an extended accessory pad and a much wider keel section.
Sex characters first appeared at lower shell-length in males than in females. Male conch also matured at lower shell-length than the females. The findings indicate that sexual dimorphisms occurred in the species and it happens very early in their life stage. Polymorphism has indeed been reported in few other Strombus species (Kuwamura et al., 1983; Reed, 1992, 1993a, b, 1995). Recently Zaidi et al. (2008) showed clear sexual dimorphism within S. canarium population, which however were only apparent when the conch reached adult stage where flaring and thickening of shell-lip was prominent.
The current study was generally in agreement with previous studies on other Strombus species where males are smaller than females, e.g. Abbott (1949) on S. gibberulus gibbosus Linnaeus, 1758, Randall (1964) on S. gigas Linnaeus, 1758, Kuwamura et al. (1983), Reed (1995) on S. luhuanus Linnaeus, 1758 and Reed (1993b) on S. pugilis Linnaeus, 1758. Zaidi et al. (2008) conclude this condition as a general characteristic of Strombidae.
CONCLUSION
The verge structure of S. canarium is unique for the species, clearly different compared with other Strombus present in the study area. The sexual characters of S. canarium appeared at lower shell-length in males compared to females. The males also matured at lower shell-length than females. The findings indicate that sexual dimorphisms occurred very early in their life history, even before the ornamentation of the shells took place.
ACKNOWLEDGMENTS
The authors would like to thanks the deanery and staffs of Biology Department, Faculty of Science, UPM and School of Environmental and Natural Resource Science, UKM for technical support and laboratory facilities provided. Special thanks to the Ministry of Science, Technology and Environment (MOSTE), Malaysia, for the research grant (04-01-02-SF0124) and JPA Malaysia for the scholarship awards, which make this study possible.
REFERENCES
- Appeldoorn, R.S., 1988. Age determination, growth, mortality and age of first reproduction in adult queen conch, Strombus gigas L., off Puerto Rico. Fish. Res., 6: 363-378.
CrossRefDirect Link - Bujang, J.S., M.H. Zakaria and A. Arshad, 2006. Distribution and significance of seagrass ecosystems in Malaysia. Aquat. Ecosyst. Health Manage., 9: 203-214.
CrossRefDirect Link - Kuwamura, T., R. Fukao, M. Nishida, K. Wada and Y. Yanagisawa, 1983. Reproductive biology of the gastropod Strombus luhuanus (Strombidae). Publ. Seto Mar. Biol. Lab., 28: 433-443.
Direct Link - Randall, J.E., 1964. Contributions to the biology of the queen conch, Strombus gigas. B. Mar. Sci., 14: 246-295.
Direct Link - Zaidi, C.C., B. Japar Sidik, A.G. Mazlan and A. Arshad, 2005. Diversity and Population Structure Characteristics of Strombus (Mesogastropod, Strombidae) in Johor Straits. In: Natural Resource Utilization and Environmental Preservation: Issues and Challenge, Sahibin, A.R. (Ed.). National University of Malaysia, Malaysia, USA., pp: 198-205.
- Cob, Z.C., A. Arshad, H.M. Idris, B. Japar Sidik and A.G. Mazlan, 2008. Sexual polymorphisms in a population of Strombus canarium Linnaeus, 1758 (Mollusca: Gastropoda) at Merambong shoal, Malaysia. Zool. Stud., 47: 318-325.
Direct Link