
Angel Alderete-Chavez
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Vicente Espinosa Hernandez
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Enrique Ojeda Trejo
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Muhammad Ehsan
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Jesus Perez Moreno
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Victor Manuel Cetina Alcala
Department of Forestry, Postgraduate College of Agricultural Sciences, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Dante Arturo Rodriguez Trejo
Division of Forest Sciences, Autonomous University of Chapingo, Km 38.5 Carr. Mex-Texcoco, Texcoco, Edo. de Mexico, C.P. 56230, Mexico
Nancy De la Cruz-Landero
Department of Soil Science, Km 36.5 Carr. Mex-Texcoco, Montecillo, Edo. de Mexico C.P. 56230, Mexico
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 604-609







ABSTRACT
In the present study, a general description of 5 species of Lupinus (Lupinus leptophyllus, L. montanus, L. potosinus, L. uncinatus, L. versicolor) growing on the oriental face of Tlalóc mountain in Sierra Nevada, México, is presented where growth of different populations was found under diverse ecological conditions: crop fields, areas disturbed by tree felling, natural aperture of the canopy and areas disturbed by forest fires. The distribution of the different species as well as its altitudinal range was determined through geo-referencing. The reported species were found at altitudes of 2800-3700 m above sea level growing in soils that range from loamy to sandy loam and sandy clay loam, with different soil physical and chemical characteristics at each site, which indicates that the species are growing in slightly acidic soils.
PDF Abstract XML References Citation
How to cite this article
Angel Alderete-Chavez, Vicente Espinosa Hernandez, Enrique Ojeda Trejo, Muhammad Ehsan, Jesus Perez Moreno, Victor Manuel Cetina Alcala, Dante Arturo Rodriguez Trejo and Nancy De la Cruz-Landero, 2008. Natural Distribution and Principal Characteristics of Lupinus in the Oriental Face of Tlaloc Mountain in Sierra Nevada, Mexico. Journal of Biological Sciences, 8: 604-609.
DOI: 10.3923/jbs.2008.604.609
URL: https://scialert.net/abstract/?doi=jbs.2008.604.609
DOI: 10.3923/jbs.2008.604.609
URL: https://scialert.net/abstract/?doi=jbs.2008.604.609
INTRODUCTION
The Lupinus species are extensively distributed around the world, without certainly knowing its evolutionary origin; nevertheless, various authors have suggested different centres of its origin. Ruiz et al. (2006) and Walker et al. (2003) are of the opinion that its origin lies in South America. The reports for Mexico indicate that mostly Lupinus species are located in the central region of Mexico: in the Neovolcánic axis at the intersection of the Sierra Madre Oriental and the Sierra Madre Occidental. The species of this genus have been studied from different perspectives: Among them are the availability of P and N in soils low in nutrients and phytoremediation of polluted soils (Bahmanyar and Ranjbar, 2008; Falkengren-Grerup and Schöttelndreir, 2004). The abundance, distribution and life cycle of the populations of native plants are influenced by climatic factors that mark the differences along the altitudinal slope (Salem et al., 2007; Phillips et al., 2002). It has been discovered, hence, that the reduction in abundance is related to the summer average temperature and low temperatures before the commencement of the winter season (Sugiyama, 2003; Diekmann and Falkengren-Grerup, 2002; Tozer and Bradstock, 2002). The natural vegetation is constantly changing by human activities and the change in climatic factors, so it is important that the bio-geographical studies about the distribution and classification of the native flora should involve environmental information, which is expected to provide more precise data about natural distribution of native species (Jalilvand et al., 2007; Jonas et al., 2002; Toft and Elliott-Fisk, 2002; Townsend and Walsh, 2001). In recent studies about classification and ordering of the vegetation in Sierra Nevada, (Sánchez-González and López-Mata, 2003) L. montanus is reported as the only species forming part of the herbaceous vegetation structure, associated to different ecosystems present in this physiographical unit. Nevertheless, other authors (Graham and Turkington, 2000) have found diverse wild species of Lupinus growing and forming populations at slopes and the foothills of mountainous places that grow and adapt well to altitudes ranging from the sea level to 4000 m, being non-resistant to prolonged frosts. In the mountainous regions all around the world the variation in the temperature is very abrupt along the slopes with the increase in altitude, the reason why this factor can be decisive for the production of fruits and seeds, allowing the establishment of new populations of plants at such slopes through seed dissemination by birds (Baret et al., 2004; Dracup and Thomson, 2000). Vital importance is attached to the studies of native species to help obtain valuable information for the rehabilitation of natural vegetation (Robles et al., 2002). In this study we present a general description of five species of Lupinus growing on the oriental face of Tlalóc Mountain in Sierra Nevada, México.
MATERIALS AND METHODS
The study area is located on the oriental slope of the Sierra Nevada in the Tláloc mountain in the State of Mexico, Mexico, between 19° 23' 43'' and 19° 28' 37'' North and between 98° 42' 51'' and 98° 48' 12'' West in an altitudinal range of 2800-4120 masl (meters above sea level) (Fig. 1). The present study is based on the samplings carried out in various field visits to the study area between the months of August, 2006 to August, 2007, searching for sites having Lupinus plantations.
Different populations were found growing in diverse ecological conditions: Fields of annual crops (Zea mays, Hordeum vulgare and Canavalia ensiformis), areas disturbed by felling of trees, natural aperture of the canopy and areas disturbed by forest fires. The sites were geo-referenced locating the geographical coordinates to know the distribution of the different populations, as well as their altitudinal range. The recently developed taxonomical keys for the valley of Mexico were employed to identify different species of Lupinus (Calderón and Rzedowski, 2005). Moreover, experts of different national herbariums were consulted to correctly identify the plant samples collected. A random soil sampling was carried out, using three sub-samples (20x20x30 cm), to constitute a compound sample for each site with different species of Lupinus. These samples were placed in polyethylene bags of 4 kg and subsequently carried to the laboratory, for the analysis of some physical and chemical soil characteristics (Table 1). The data were subjected to one way ANOVA followed by Tukey test for multiple comparison of means (Significance level p<0.05) using STATISTICA (Stat-Soft Inc., 2003).
 | |
| Fig. 1: | The study area is located on the oriental slope of the Sierra Nevada in the Tláloc mountain in the State of Mexico, Mexico |
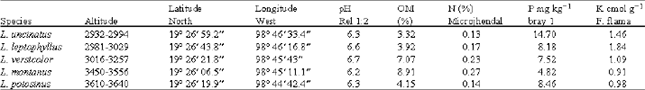
| Table 1: | Species, geographical coordinates and altitude, as well as the main characteristics of associated soils for each species |
 | |
RESULTS AND DISCUSSION
As it can be seen in Table 1, some species share the altitudinal range among them, even in the case where only dispersed individual plants of each one of them may exist. On the other hand different species of Lupinus are found growing on sites with slopes ranging from 2 to 73% where the slope is abrupt with very difficult access. The species reported in this study were found at altitudes of 2800-3700 masl.
Lupinus species found in the oriental part of the Tlalóc mountain in Sierra Nevada, Mexico: Lupinus leptophyllus Schlecht and Cham, can be found in plantations between 2981-3029 masl and at slopes of 2-30%, exactly in the area which is border limit between the cultivated fields and the plantations of L. uncinatus. L. leptophyllus grows at 3170 masl in the forests of Abies religiosa. It`s an annual plant, of short life having stems more than l m long with solid marrow, erect or ramified, purple stipule 5 to 12 mm long, 4 to 8 cm long leaf stalk, 6 to 8 leaflets (uptill 10 leaflets observed in this study) 4.5-8 cm long and 9-15 mm wide, widespread flowers with sharp and feathery apex, that disappears with the age, of purple colour, with whitish wings, the fruits are enclosed in hairy pods 4-5 cm long and 8-9 mm wide, containing 26 to 98 pods per branch and 2 to 9 mature seeds per pod, flowering initiates since the last days of January and the first mature seeds (dark to dark brown) appear in March (Fig. 2). This species is found in the soils with higher pH (6.6), the soil Organic Matter content (OM) is of 3.92%, contain 0.17% of total Nitrogen, 8.18 mg kg-1 of soil Phosphorus (P) and 1.84 cmol g-1 of soil Potassium (K) (Table 1).
Lupinus montanus H.B.K: Although this species can be found on the road side at an altitude since 3100 masl, grows at altitudes ranging between 3450-3556 masl in forests of Pinus hartwegii with a slope of 15-73% where its dense plantations can be found. It is apparently the most widespread species in the study area, forming compact plantations of about 10 ha or more, mostly in the forest of Pinus hartwegii subjected to intervention by forest fire. The plants of this species are annual, biannual, or perennial of more than 1.5 m length, hollow stems 4 to 15 mm wide, stipules 3 to 10 cm long, 10 to 14 leaflets (in the present study uptill 17 leaflets found), sharp apex, verticillate, purple, bigger flowers than other species, feathery apex, thick floral pedicel, at times bifurcated. Its fruits are pods, 4 to 5 cm long and 9 to 10 mm wide, containing about 45-73 pods per branch, 3-9 dotted to black mature seeds per pod, flowers at the end of May


(Fig. 3) Lupinus montanus is found in the soils with higher pH (6.2), the soil organic matter content (OM) of 8.91%, with 0.27% of total Nitrogen, 4.82 mg kg-1 of Phosphorus content (P) and 0.91 cmol g-1 of soil Potassium (K) (Table 1).
Lupinus potosinus rose: This plant is found rarely in the area, growing in isolated form at altitudes of 3610 to 3640 masl and slope of 5-17%, in disturbed forests of Pinus hartwegii and on the roadsides, its height ranges from 45-75 cm with pubescence in the stem, slightly ramified, petioles 5-7 cm long with 5 to 7 leaflets, pubescent pods with abundant hair, 3-4 cm long, 15 to 35 pods per branch and 4-7 seeds per pod, the flowers are little, of blue colour and white wings at the tip, flowers from May to August (Fig. 4). This species is found in the soils with higher pH (6.2), the soil Organic
 | |
| Fig. 4: | Lupinus potosinus Rose |
Matter (OM) content is of 4.15 %, with 0.14 % of total Nitrogen, 8.46 mg kg-1 of soil Phosphorus (P) and 0.98 K cmol g-1 of soil Potassium (K) (Table 1).
Lupinus uncinatus schlecht: Dense populations of this species were found in cultivated fields and on the roadside between 2932-2994 masl and slope of 2-23%. It is an annual, biannual, or perennial plant, more than 1 m tall, herbaceous, of hollow stems, stipules 7 to 9 mm long (not always), petiole of 1.5-12.5 cm length, 5 to 8 leaflets 3 to 4 cm long and 6 to 8 mm wide, slightly sharp apex, widespread deciduous flowers with feathery apex of dark purple colour, the fruits are dehiscent pods 4.5 to 5.5 cm long, containing 9 to 49 pods per branch and 1 to 7 mature seeds per pod, flowering starts since March and the first mature seeds (whitish to ash grey) appear in the middle of May, the seeds being of an orthodox type, rare in this genera (Fig. 5). It is found in the soils with higher pH (6.3), 3.32% soil Organic Matter (OM) content, 0.13% total Nitrogen, 14.7 mg kg-1 soil Phosphorus (P) and 1.46 cmol g-1 soil Potassium (K) (Table 1).
Lupinus versicolor sweet: It can be found at 3016-3257 masl and slope of 7 to 21% on the roadsides and in disturbed forests of Abies religiosa, its height is less than 1 m, with 3 to 3.5 cm long petioles and from 6-7 oblong to obtuse slightly hairy leaflets, verticillate flowers, 32 to 64 pods per branch and 4 to 9 seeds per pod of dark brown color, flowers in the months of April to August (Fig. 6) Lupinus versicolor is found in the soils with higher pH (6.7), the soil Organic Matter content (OM) of 7.07%, containing 0.23% of total Nitrogen, 7.52 mg kg-1 of soil Phosphorus content (P) and 1.09 cmol g-1 soil Potassium (K) (Table 1).


CONCLUSIONS
The Lupinus species are distributed widely in the oriental part of Tlaloc mountain found in an altitudinal range varying from 2932 to 3640 m above sea level. In two earlier studies regarding genus Lupinus in this area, two species have been reported: L. uncinatus (Ehsan et al., 2007) and L. montanus (Sánchez-González and López-Mata, 2003). In the present research, we described three additional species of this genus growing in the study area. The given species have well adapted to the climatic and edaphic conditions of the area including soils with varying contents of NPK and organic matter and are associated to various ecosystems with different degrees of intervention (Forest fire, tree felling, traditional cropping and animal grazing areas) (Reyes and Casal, 2004; Valbuena and Trabaud, 2001). The soils in the study area with Lupinus growth range from loamy to sandy loam and sandy clay loam where different soil chemical characteristics may be related to the presence of the diverse reported species in this study (Andrist-Rangel et al., 2007; Watt and Evans, 1999; Gardner et al., 1982). Future studies on different aspects of this genus including agronomy, breeding, phytochemistry and classification is required to fully harness its potential.
ACKNOWLEDGMENTS
The authors express their gratitude to the National Council of Science and technology (CONACYT) for the grant of the scholarship number 177441 and to the Program of Improvement for the Faculty for the grant of the scholarship for studies of high quality (PROMEP/103.5/06/1973) without which the present study would not have been possible.
REFERENCES
- Andrist-Rangel, Y., A.C. Edwards, S. Hillier and I.O. Born, 2007. Long-term K dynamics in organic and conventional mixed cropping systems as related to management and soil properties. Agric. Ecosyst. Environ., 122: 413-426.
CrossRefDirect Link - Bahmanyar, M.A. and G.A. Ranjbar, 2008. The role of potassium in improving growth indices and increasing amount of grain nutrient elements of wheat cultivars. J. Applied Sci., 8: 1280-1285.
CrossRefDirect Link - Baret, S., M. Sandrine, T. Le-Bourgeois and D. Strasberg, 2004. Altitudinal variation in fertility and vegetative growth in the invasive plant Rubus alceifolius poiret (Rosaceae), on Reunion Island. Plant Ecol., 172: 265-273.
Direct Link - Diekmann, M. and U. Falkengren-Grerup, 2002. Prediction of species responses to atmospheric nitrogen deposition. J. Ecol., 90: 108-120.
CrossRef - Dracup, M. and B. Thomson, 2000. Narrow-leafed lupins with restricted branching. Ann. Bot., 85: 29-35.
Direct Link - Ehsan, M., P.A. Molumeli, V.E. Hernández, A.B. Reyes and J.P. Moreno et al., 2007. Ontamination time effect on plant available fractions of cadmium and zinc in a Mexican clay loam soil. J. Applied Sci., 7: 2380-2384.
Direct Link - Falkengren-Grerup U. and M. Schöttelndreier, 2004. Vascular plants as indicators of nitrogen enrichment in soils. Plant Ecol., 172: 51-62.
CrossRefDirect Link - Graham, A.S. and R. Turkington, 2000. Population dynamics response of Lupinius arcticus to fertilization, clipping and neighbour removal in the understory of the boreal fores. Can. J. Bot., 78: 753-758.
Direct Link - Jalilvand, H., Y. Kooch, M.A. Bahamnyar and M.R. Pormajidian, 2007. Ecological species groups of hornbeam forest ecosystems in Southern Caspian (North of Iran). J. Biol., Sci., 7: 1504-1510.
CrossRefDirect Link - Jonas, E., E.J. Lawesson and F. Skov, 2002. The phytogeography of Denmark revisited. Plant Ecol., 158: 113-122.
Direct Link - Phillips, P.D., I. Yasman, T.E. Brash and P.R. Var Gardingen, 2002. Grouping tree species for analysis of forest data in Kalimantan (Indonesia Bormeo). For. Ecol. Manage., 157: 205-216.
Direct Link - Reyes, O. and M. Casal, 2004. Effects of forest fire ash on germination and early growth of four Pinus Species. Plant Ecol., 175: 81-89.
Direct Link - Robles, A., B. Allegretty and Passera, 2002. Coronilla juncea is both a nutritive fodder shrub and useful in the rehabilitation of abandoned Mediterranean marginal farm land. J. Arid Environ., 50: 381-392.
Direct Link - Ruiz, L.M.A., M.R. Rodriguez and P.S. Navarro, 2006. Evaluación químico-nutricional de Lupinus exaltatus Zucc, Del Nevado de Colima, México, como fuente potencial de forraje. Interciencia, 31: 758-760.
Direct Link - Salem, F.B., M. Tarhouni, B.A. Ouled and M. Neffati, 2007. Impact of drought on plant cover dynamics in two natural areas of Southern Tunisia. J. Biol. Sci., 7: 1539-1544.
CrossRefDirect Link - Sugiyama, S., 2003. Geographical distribution and phenotypic differentiation in populations of Dactylis Glomerata L. in Japan. Plant Ecol., 169: 295-305.
CrossRefDirect Link - Toft, C. and D. Elliott-Fisk, 2002. Patterns of vegetation along a spatiotemporal gradient on shoreline strands of a desert basin lake. Plant Ecol., 158: 21-39.
Direct Link - Townsend, A.P. and S.J. Walsh, 2001. . Remote sensing of forested wetlands: Application of multitemporal and multispectral satellite imagery to determine plant community composition and structure in Southeastern USA. Plant Ecol., 157: 129-149.
Direct Link - Tozer, M.G. and R.A. Bradstock, 2002. Fire-mediated effects of overstorey on plant species diversity and abundance in an Eastern Australian heath. Plant Ecol., 164: 213-223.
CrossRefDirect Link - Valbuena, L. and L. Trabaud, 2001. Contribution of the soil seed bank to post-fire recovery of a heathland. Plant Ecol., 152: 175-183.
Direct Link - Walker, L.R., B.D. Clarkson, W. Silvester and B.R. Clarkson, 2003. Facilitation outweighs inhibition in pos-volcanic primary succession in New Zealand. J. Veg., Sci., 14: 277-290.
CrossRef