
E. Farshadfar
College of Agriculture, Razi University, Kermanshah, Iran
S. Mahjouri
College of Agriculture, Razi University, Kermanshah, Iran
M. Aghaee
Seed and Plant Improvement Institute, Karaj, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 598-603







ABSTRACT
In order to detect gene action and epistatic effects for drought tolerance indicators, 22 diverse genotypes of durum wheat were used in a randomized complete block design with three replications under rainfed and irrigated conditions in the Agricultural Research Center of Kermanshah, Iran. Analysis of variance revealed enough genetic variation and possibility of selection between genotypes. Mean comparison, using Duncan multiple range test, indicated that genotypes 12, 13 and 21 were suitable in drought conditions. In three-dimensional plot, based on STI index, genotype 21 was grouped as a one cluster. Triple test cross analysis indicated epistatic effects for heading date, ruin irritability and proline content in the rainfed condition. Both additive and non-additive gene actions were significant for grain yield, proline content and ruin irritability in the water stress condition. For these traits, selection in segregating generations and hybridization breeding methods were offered. Only additive gene action was predominant for heading date in the rainfed condition. Thus, selection in early segregated generations and massive selection are considered. Higher D than H for all characters was showing incomplete dominance. Correlation coefficient(r) between sums and differences of lines were non-significant, indicating ambidirectional nature of dominant genes between lines.
PDF Abstract XML References Citation
How to cite this article
E. Farshadfar, S. Mahjouri and M. Aghaee, 2008. Detection of Epistasis and Estimation of Additive and Dominance Components of Genetic Variation for Drought Tolerance in Durum Wheat. Journal of Biological Sciences, 8: 598-603.
DOI: 10.3923/jbs.2008.598.603
URL: https://scialert.net/abstract/?doi=jbs.2008.598.603
DOI: 10.3923/jbs.2008.598.603
URL: https://scialert.net/abstract/?doi=jbs.2008.598.603
INTRODUCTION
Water deficit is the major constraint to rainfed durum wheat production worldwide (Kristin et al., 1997; Andrew et al., 2000).
Durum wheat (Triticum turgidum L. var. durum Desf.) as a self-pollinated tetraploid (2n = 4x = 28) is highly valued for production of semolina and pasta products. It is occupying about 30 million hectares and 8% of total production throughout the world (Sharma and Sain, 2004). Recently, it has been a renewed interest in durum cultivation and its breeding in Iran. Considering the lower potential yield in durum than that of bread wheat under both rainfed and irrigated conditions, more investigations and breeding programs for development of higher-yielding and best cultivars are necessary. Drought causes some deficits in plant characters like yield. The identification of drought tolerance as the ability of plants to grow satisfactorily when exposed to water defects has little direct applicability to either quantifying or breeding for the characters in crop species (Clarck et al., 1992).
To formulate on efficient breeding program for developing drought tolerant varieties, it is essential to understand the mode of inheritance (Farshadfar et al., 2000; Morgan, 1991). Primarily, biological variation present in the plant population is of three types, viz., phenotypic, genotypic and environmental. The genotypic variance consists of additive, dominance and epistatic components. There is much more evidence that study of epistatic gene action or non-allelic interaction in quantitative trait inheritance is important in making a decision for the plant breeding (Khattak et al., 2002; Sharma et al., 2003; Rebetzke et al., 2004; Sood et al., 2007). Mather and Jinks (1982) divided the phenotypic variance into three components, namely, (1) heritable fixable (additive variance), (2) heritable non-fixable (dominance and epistatic components) and (3) non-heritable non-fixable (environmental fraction). In fact, the heritable fixable component of phenotypic variance will include the additive x additive fraction of the epistatic variance as well. Further, the total phenotypic variance may be partitioned as (1) fixable (additive and additive x additive components) and (2) non-fixable (dominance, additive x dominance and dominance x dominance types of epistasis and environmental fraction) components (Singh and Narayanan, 1990, 1993; Singh and Chaudhary, 1999). Cockerham (1961) partitioned the epistatic effects into additive x additive, additive x dominance and dominance x dominance components.
In order to detect epistatic effects different methods viz., scaling test in the generation mean analysis, evaluation of Wr/Vr ratio in the diallel mating design and triple test cross analysis have been used (Sharma, 1998). Kearsey and Jinks (1968) suggested an extension of design III of Comstock and Robinson (1952) in such a manner that besides estimating additive and dominance variance, the presence or absence of gene interaction may be tested without any difficulty. This design is Known as Triple test cross method (Singh and Chaudhary, 1999).
The Triple Test Cross (TTC) is one of the multiple mating design that helps to estimate the genetic architecture of polygenic characters. Its power in detecting epistasis and flexibility in terms of accommodating large samples from the population are some of its advantages (Pooni et al., 1994; Kearsey and Jinks, 1968; Kearsey and Pooni, 1998).
This design is most flexible in that it can be applied to any population with any level of inbreeding, any gene frequency and degree of linkage disequilibrium or gene correlations. In the absence of epistasis this design provides a more efficient estimate of additive and dominance components (Viana, 2005; Roy, 2000).
The objectives of the present investigation were to determine the genetic analysis of drought tolerance criteria in durum wheat; to detection of non-allelic interaction for the characters under investigation and the estimation of additive and dominance components of genetic variance for the traits not influenced by epistasis.
MATERIALS AND METHODS
To evaluate gene action and detecting epistatic effects of drought tolerance indicators in durum wheat, 22 diverse plant genetic materials were used in a randomized complete block design with three replications in the Agricultural Research Institute of Kermanshah, Iran during 2000-2004.
Two homozygous parents (P1 and P2) having different allelic genes at two loci, unrelated and genetically divergent were crossed and their subsequent generations: F1(P1 x P2), F2(selfed F1), F3(selfed F2), BC1(P1 x F1), BC2(P2 x F1); (8-21) genotypes numbered 8-21 obtained from crosses between F1 and pure lines as testers. These genotypes plus durum wheat (Zardak) as a guard plant to eliminate border effect were analyzed. Twenty one genetic materials were cultivated in rows with 50 cm row to row and 20 cm plant to plant spacing. After harvesting, 5 seeds from each generation were randomly selected and transplanted into 20x25 cm pots in the framework of randomized complete block design with three replications under two different water regimes (irrigated and rainfed).
After ten days data of 5 randomly selected plants were recorded on Grain Yield (GY) per pot in the stress (YS) and non-stress (YP) conditions. Number of seed per spike (NSPS), number of spike per plant (NSPP), 100 seed weight (HSW), biomass, Harvest Index (HI), earliness (days to heading = DH), Proline Cumulating Content (PCC) and Ruin Irritability (RI) were determined on the leaves in the stress condition as described by Bates et al. (1973), Hasiao (1973) and Levitt (1980).
Stress Tolerance Index (STI) was calculated using the formula suggested by Fernandez (1992) as:
analysis of variance, mean comparison and three dimensional plot were done using statistical softwares MSTAT-C and SPSS.
Triple test cross analysis (Kearsey and Jinks, 1968; Kearsey and Pooni, 1998) was used to detect epistasis. The presence of epistasis was indicated when the mean squares for the deviations ![]() were significantly greater than the error. This analysis is valid regardless of gene frequency, degree of inbreeding and linkage relationships (Pooni et al., 1994).
were significantly greater than the error. This analysis is valid regardless of gene frequency, degree of inbreeding and linkage relationships (Pooni et al., 1994).
Additive and dominance components were estimated from the analysis of sums and differences assuming no epistasis. The Expected Mean Squares (EMS) of the sums and differences are as follows:
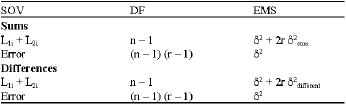 |
| The expectations for δ2sum and δ;2difference in the absence of epistasis and linkage is |
The average degree of dominance was estimated as:
The direction of dominance was determined as the correlation coefficient (rs,d) of the sums and differences (Mather and Jinks, 1982). Significant positive and negative correlation would indicate a predominant direction of dominance towards decreasing and increasing values of the trait.
RESULTS AND DISCUSSION
The results of analysis of variance (Table 1) under rainfed condition indicated highly significant differences for Yp, Ys, NSPS, NSPP, HSW, biomass, HI, earliness, proline content, ruin irritability and stress indicating the presence of genetic variation and possibility of selection for drought tolerant genotypes.
Mean comparison in the stress condition using Duncan's multiple range test (Table 2) revealed that genotypes 12, 13 and 21 had the highest grain yield, proline content and the lowest ruin irritability.
A three-dimensional plot between Yp, Ys and STI (Fig. 1) showed genotype 21 in group A i, e perform favorably in both stress and non-stress environments (Fernandez, 1992; Farshadfar et al., 2004; Farshadfar and Sutka, 2003) as there is no significant difference between genotype 12, 13 and 21 (Table 2), hence these genotypes are recommended as the most drought tolerant ones according to the results of this experiment.
Considerable genetic variation was observed among treatments for all traits (Table 3). Significant differences between parents (Table 3) for grain yield in both conditions, number of seed per spike, number of spike per plant, harvest index, proline content and ruin irritability indicated that parents were different from each other. So with absence of non-allelic interaction, the estimation of additive and dominance genetic components of variance would be unbiased. These results were in agreement with the results of Khattak et al. (2002) in mungbean. Triple test cross analysis (Table 4) exhibited significant overall
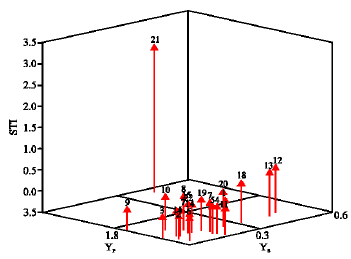 | |
| Fig. 1: | Three-dimensional plot between YP, YS and STI |
| Table 1: | Analysis of variance for studied characters under water stress condition |
 | |
| **Significant at 0.01 probability level | |
| Table 2: | Mean comparisons of YP and characters under water stress condition |
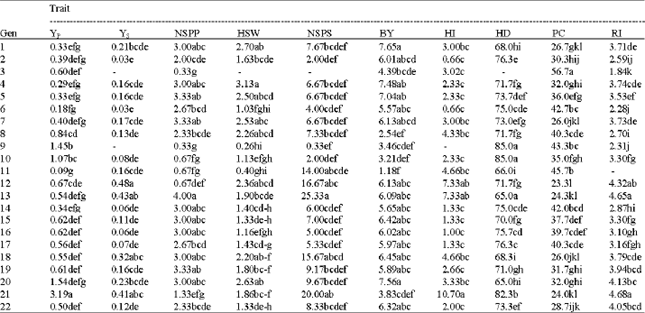 | |
| Mean values with the different letter(s) are not statically significant | |
| Table 3: | Analysis of variance showing mean squares for testing epistasis under stress condition |
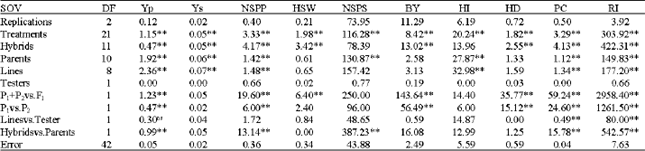 | |
| **; ns: Significant at 0.01 probability level and non-significant, respectively | |
| Table 4: | Analysis of variance showing mean squares for testing epistasis in stress |
| **: Significant at 0.01 probability level | |
| Table 5: | Mean Squares for sums and differences together with their errors for characters under stress condition |
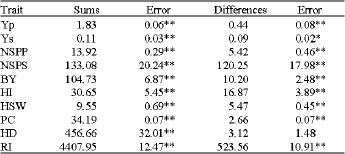 | |
| **, *: Significant at 0.01 probability level and at 0.05, respectively | |
epistasis effects for grain yield, earliness, proline content and ruin irritability in the stress condition. Sood et al. (2007) and Rebetzke et al. (2004) observed epistasis in some studied traits in Linum and some crosses in a wheat variety, respectively. Non-allelic interaction was reported by Sharma and Sain (2004) in a study of amelioration of grains per spike in durum wheat plants. These results indicated that epistasis could not be ignored in genetic models. Epistatic components related to a large number of interacting genes, lead to inefficient selection. But if the number of interacting genes will be reduced, selection will be efficient relied on the magnitude due to dominance and environmental variances.
Epistatic effects for quantitative traits were less stable due to strong interaction with environment as compared with additive (sums) and dominant (differences) effects (Upadhyaya and Nigma, 1998; Perkins and Jinks, 1971). Environmental interactions depend on the number of genes involved in the inheritance and as the number of genes increases; opportunities for environmental influences become greater (Gamble, 1962). This could explain the strong epistasis by environment interaction in this investigation. Therefore, population improvement through pedigree selection or single seed descent might give a good response (Jensen, 1970).
The analysis of variance for sums and differences between genotypes (Table 5) was used to detect additive and dominance effects, respectively in the presence of epistasis. Estimates of the sums revealed highly significant additive component for all studied characters (Table 5). The predominance of additive component suggested that selection in early segregating generations would be effective for improving these traits.
In the presence of epistasis, estimates of additive component (D) were affected by interactions (additive x additive and additive x dominance) in loci (Pooni et al., 1994). The analysis of differences (dominance component) was highly significant for most of the characters except heading date (earliness) in the stress condition. Therefore, only additive effects were important in the genetic control of earliness. Significant and non-significant dominance effects for some traits in the stress and non-stress conditions displayed a strong different environmental interaction (Upadhyaya and Nigma, 1999). The dominance variance estimated in the triple test cross is related to the different loci of the L1 and L2 testers (Parents). When the dominance component is non-significant, an additive component free of the dominance contamination is estimated irrespective of the number of loci by which L1 and L2 differ (Mather and Jinks, 1971).
For the traits with significant mean squares of sums and differences both additive and dominance components were involved in their genetic control. Therefore, hybridization breeding and selection in segregating generation are recommended for their improvement. As estimate of D was higher than H for all the characters, hence, the degree of dominance was less than unity indicating the existence of incomplete and partial dominance. Detection of components in Linum studied by Sood et al. (2007) using TTC, showed additive (D) type of
| Table 6: | Estimated of additive (D) and dominance (H) variance components, degree of dominance and correlation of sums and different for characters under stress condition |
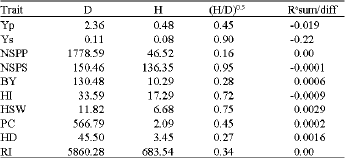 | |
gene action for the studied traits such as Plant height, capsule per plant, seed yield, 1000-seed weight, harvest index. In faba bean (Vicia faba), significant mean squares of additive and dominance components for the characters such as plant height, days to first flower, pods/plant, 100-seed weight and yield have also been reported (Bakheit et al., 2002).
Correlation of sums and difference (r) was non-significant for all the traits (Table 6), suggesting ambidirectional dominance (increasing and decreasing alleles) were more or less equally distributed among the genotypes. This result is similar to the results in maize reported by Gautaman (2003) and Bhatnagar (2004). Also, Kearsey et al. (2003) in similar research on 22 interesting traits in some generations of Arabidopsis reported epistasis and partial dominance for most of the traits.
For the characters (earliness) that both dominance (H) and correlation coefficient (r) were non-significant in the stress condition, dominance effects were not important or dominant genes had both positive and negative effects in its genetic control.
Genotype x environment interactions may mask the expression of epistatic effects (Tefra and Peat, 1997), therefore a series of experiments are required for efficient breeding procedures (Jinks et al., 1969; Subbaraman and Rangasamy, 1989). Epistasis has been reported in wheat for grain yield (Singh and Singh, 1976), tiller number and harvest index (Nanda et al., 1989) and in rice for tiller number and earliness (Subbaraman and Rangasamy, 1989).
In general additive, dominance and epistatic components are important in durum wheat, but as it is an autogamous plant only the additive component is important to develop pure breeding varieties from any hybridization program. This result is in agreement with the findings of Tefra and Peat (1997). In this study, it is shown that significant additive component (D) for the character under investigation is an advantageous for selection in advanced generations as homozygosity become fixable.
REFERENCES
- Borrell, A.K., G.L. Hammer and R.G. Henzell, 2000. Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci., 40: 1037-1048.
CrossRefDirect Link - Bakheit, B.R., M.Z. El-Hifny, M.M. Eissa and S.B. Ragheb, 2002. Triple test cross and six population techniques for partitioning the components of genetic variance in faba bean (Vicia faba). J. Agric. Sci., 139: 61-66.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bhatnagar, S., F.J. Betran and L.W. Rooney, 2004. Combining ability of quality protein maize inbreds Crop Sci., 44: 1997-2005.
Direct Link - Clark, J.M., R.M. DePauw and T.F. Townley-Smith, 1992. Evaluation of methods for quantification of drought tolerance in wheat. Crop Sci., 32: 723-728.
CrossRefDirect Link - Clark, J.M., R.M. DePauw and T.F. Townley-Smith, 1992. Evaluation of methods for quantification of drought tolerance in wheat. Crop Sci., 32: 723-728.
CrossRefDirect Link - Farshadfar, E., M. Farshadfar and J. Sutka, 2001. Combining ability analysis of drought tolerance in wheat over different water regimes. Acta Agron. Hung., 48: 353-361.
CrossRefDirect Link - Farshadfar, E. and J. Sutka, 2003. Locating QTLs controlling adaptation in wheat using AMMI model. Cereal Res. Commun., 31: 249-254.
Direct Link - Farshadfar, E., R. Mohammadi, M. Farshadfar and J. Sutka, 2004. Locating QTLs controlling field and laboratory predictors of drought tolerance in Agropyron using multiple selection index. Cereal Res. Commun. 32: 17-24.
Direct Link - Gamble, E.E., 1962. Gene effects in corn (Zea mays L.): III. Relative stability of the gene effects in different environments. Can. J. Plant Sci., 42: 626-634.
CrossRefDirect Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Jensen, F., 1970. A diallel selective mating system for cereal breeding. Crop Sci., 10: 629-635.
CrossRefDirect Link - Jinks, J.L., J.M. Perkins and E.L. Breese, 1969. A general method of detecting additive, dominance and epistatic variation for metrical traits II. Application to inbred lines Heredity, 24: 45-57.
CrossRefDirect Link - Jinks, J.L. and J.M. Perkins, 1970. A general method for the detection of additive, dominance and epistatic components of variation III. F2 and backcross populations. Heredity, 25: 419-429.
CrossRefDirect Link - Kearsey, M.J. and J.L. Jinks, 1968. A general method of detecting additive, dominance and epistatic variation for metrical traits I. Theory. I. Heredity, 23: 403-409.
CrossRefDirect Link - Kearsey, M.J., H.S. Pooni and N.H. Syed, 2003. Genetics of quantitative traits in Arabidopsis thaliana. Heredity, 91: 456-464.
CrossRefDirect Link - Khattak, G.S.S., M.A. Haq, M. Ashraf and G.R. Tahir, 2002. Triple test cross analysis for some morphological traits in mungbean (Vigna radiate (L.) Wilczek). Euphytica, 126: 413-420.
Direct Link - Kristin, A.S., M.E. Brothers and I.D. Kelly, 1997. Marker assisted selection to improve drought resistance in common bean. Crop Sci., 37: 51-60.
Direct Link - Morgan, M., 1991. A gene controlling difference in osmoregulation in wheat. Aust. J. Plant Physiol., 18: 249-257.
Direct Link - Nanda, G.S., G. Singh and K. Chand, 1990. Detection of components of genetic variation and prediction of the frequencies of transgressive segregants in bread wheat (Triticum aestivum L.). J. Genet. Breed., 44: 63-66.
Direct Link - Perkins, J.M. and J.L. Jinks, 1971. Analysis of genotype × environment interaction in triple test cross data. Heredity, 26: 203-209.
CrossRefDirect Link - Pooni, H.S., I. Kumar and G.S. Khush, 1994. A general method of detecting additive, dominance and epistatic variation for metrical traits. V. Triple test cross analysis of disomically inherited traits expressed in triploid tissues. Heredity, 72: 563-569.
CrossRefDirect Link - Rebetzke, G.J., T.L. Botwright, C.S. Moore, R.A. Richards and A.G. Condon, 2004. Genotypic variation in specific leaf area for genetic improvement of early vigor in wheat. Field Crops Res., 88: 179-189.
Direct Link - Sharma, S.N., R.S. Sain and R.K. Sharma, 2003. Genetics of spike length in durum wheat. Euphytica, 130: 155-161.
CrossRef - Sharma, S.N. and R.S. Sain, 2004. Genetics of grains per spike in durum wheat under normal and late planting conditions. Euphytica, 139: 1-7.
CrossRefDirect Link - Singh, S. and R.B. Singh, 1976. Triple test cross analysis in two wheat crosses. Heredity, 37: 173-177.
CrossRefDirect Link - Sood, S., N.R. Kalia, S. Bhateria and S. Kumar, 2006. Detection of genetic components of variation for some biometrical traits in Linum usitatssimum L. in sub-montain Himalayan region. Euphytica, 155: 107-115.
CrossRef - Subbaraman, N. and S.R.S. Rangasamy, 1989. Triple test cross analysis in rice. Euphytica, 42: 35-40.
CrossRefDirect Link - Tefra, H. and W.E. Peat, 1997. Genetics of grain yield and other agronomic characters in (Eragrostis tef Zucctrotter ). II the triple test cross. Euphytica, 96: 193-202.
Direct Link - Upadhyaya, H.D. and S. Nigam, 1998. Epistasis for vegetative and reproductive traits in peanut. Crop Sci., 38: 44-49.
CrossRefDirect Link - Upadhyaya, H.D. and S.N. Nigma, 1999. Detection of epistasis for protein and oil contents and oil quality parameters in peanut. Crop Sci., 39: 115-118.
Direct Link - Viana, J.M.S., 2005. Dominance, epistasis, heritability and expected genetic gains. Genet. Mol. Biol., 28: 67-74.
Direct Link