
A. Mehrvar
Department of Plant Protection, Maragheh Higher Education Complex, University of Tabriz, Tabriz-5166616471, Iran
R.J. Rabindra
Project Directorate of Biological Control (ICAR), P.B. No. 2491, Bellary Road, Hebbal, Bangalore-560024, Karnataka, India
K. Veenakumari
Project Directorate of Biological Control (ICAR), P.B. No. 2491, Bellary Road, Hebbal, Bangalore-560024, Karnataka, India
G.B. Narabenchi
Pest Control (India) (PCI): Bio-Control Research Laboratories (BCRL), Bangalore-560064, Karnataka, India
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 534-541







ABSTRACT
Experiments conducted to study the effect of various adjuvants on the larval mortality of Helicoverpa armigera NPV under simulated sunlight revealed that all the adjuvants (either alone or in combination) resulted in significantly higher larval mortality as compared to the virus alone. A combination of three adjuvants, viz., molasses (5.0%) + Tinopal (0.2%) + lampblack (0.1%) showed the highest larval mortality (94.2%) under simulated sunlight with the lowest LT50 values (99.6 h). It was also observed that the persistence of HearNPV was higher when molasses (5.0%) + Tinopal (0.2%) + lampblack (0.1%) was used as adjuvant with an OAR (original activity remaining) of 86 and 36.2% after 1 and 6 days after treatment, respectively under open weather conditions on tomato plants.
PDF Abstract XML References Citation
How to cite this article
A. Mehrvar, R.J. Rabindra, K. Veenakumari and G.B. Narabenchi, 2008. Evaluation of Adjuvants for Increased Efficacy of HearNPV Against Helicoverpa armigera (Hubner) Using Suntest Machine. Journal of Biological Sciences, 8: 534-541.
DOI: 10.3923/jbs.2008.534.541
URL: https://scialert.net/abstract/?doi=jbs.2008.534.541
DOI: 10.3923/jbs.2008.534.541
URL: https://scialert.net/abstract/?doi=jbs.2008.534.541
INTRODUCTION
Adjuvants are known to increase the persistence of nucleopolyhedroviruses in the environment. Persistence generally refers to the ability of a pathogen to remain viable in the environment. Being obligate pathogens, viruses cannot multiply in outside the environment of the host insect and have to remain in a viable state before they are ingested by the host insect (Jacques, 1985). Inactivation of viruses on foliage of plants has been a major problem in the development of formulations of viral insecticides for use in insect management systems. Persistence of viruses on foliage is influenced by sunlight, pH, temperature and moisture. Sunlight is the primary factor limiting viral persistence in the environment. Virus inactivation by sunlight is caused primarily by ultraviolet light (UV) (Young, 2000). The medium wave or erythermal UV band (UV-B of 280-320 nm) is the most important photoinactivator of baculoviruses, with considerable but slower effect in the near-UV region (320-400 nm) and in some cases some effect above this (Griego et al., 1985; Martignoni and Iwai, 1985; Killick, 1986; Jones et al., 1993). A combination of UV-B (280-320 nm) and UV-A (320-400 nm) wavelengths reaches the surface of the earth. Since pathogens are exposed only to longer wavelengths of UV light, (above 290 nm) any UV inactivation must be due to these higher wavelengths (Young, 2000).
Use of adjuvants increases the efficacy of the virus. This is usually attributed to the improved field persistence of the virus or due to increased consumption of the virus by the pest (Luttrell et al., 1983). A variety of adjuvants have been examined for their potential to slow inactivation of viruses and prolong their persistence (Young, 2000). Evaluation of adjuvant materials such as stickers, wetters, phagostimulants and sunscreens in baculovirus formulations has been well documented by Rabindra and Jayaraj (1988, 1992, 1995), Behle et al. (2003), Martinez et al. (2004), Arthures et al. (2005) and Dougherty et al. (2006). Rabindra and Jayaraj (1989) reported that Robin blue and Tinopal when used as adjuvants increased the persistence of the virus on foliage and suggest that this could be attributed to the UV protection. Shapiro et al. (1983) evaluated different adjuvants and concluded that molasses, Shade and Coax served as UV protectants at 5% concentration.
However, expansion of baculovirus use will depend on further research on the identification of different adjuvants which will improve their environmental persistence. The type of adjuvants used will depend on the different ecosystems in which it has to be used and we need to consider the availability, cost and ease of application of these adjuvants under local conditions.
MATERIALS AND METHODS
The nucleus culture of H. armigera NPV (HearNPV) was obtained from the microbial repository of the Project Directorate of Biological Control (PDBC), Bangalore. Various adjuvants either alone or in combination were evaluated for their ability to increase the larval mortality of H. armigera. The different adjuvants and the concentrations tested are shown in Table 1. The adjuvants tested locally against natural sunlight under Indian open weather conditions (Asokan, 1998; Rabindra and Jayaraj, 1988, 1989, 1992, 1995) were selected and tested in this experiment for their efficacy under controlled conditions using a suntest machine in PDBC during the years 2005 and 2006. Among these adjuvants chosen, a few were selected based on their ability to increase larval mortality, were cheaper and easily available locally. They were used to test the persistence of the virus on tomato plants in PDBC during the year 2006.
Preparation of adjuvants: Cotton Seed Kernel Extract (CSKE) was prepared by homogenizing dehusked cotton seeds in a blender and sieving 2-3 times through a muslin cloth. Suspensions of soya flour, starch, maize flour, molasses, crude sugar, turmeric, food colours and lampblack were prepared in distilled water. Polyflavonoids from mango and guava leaves were extracted using 80% methanol and then lyophilized in a freeze-drier (Asokan, 1998). All the adjuvants were locally sourced expect for Tinopal (Ranipal®, Pidlite Industries Led, Mumbai) and Boric acid (HiMedia RM 1224-500G).
To evaluate the single adjuvants 100 μL of virus (1x108 OB mL-1) was added to 900 μL of adjuvant to attain a concentration of 1x107 OB mL-1. Teepol was also added at 0.1% to all the suspensions. Five hundred microlitter of each of these suspensions was applied onto plastic sheets (6x12 cm) using a micropipette and spread uniformly with the blunt end of a sterile 6 mm polished glass rod. After air drying, treated sheets were irradiated in a Suntest machine (Atlas Suntest CPS+/XLS+, Atlas Material Testing Technology GmbH, Vogelsbergstra βe, 22) at 500 W m-2 for 90 min.
Studies were conducted to compare the irradiance (W m-2) in different categories of natural sunlight (different hours of the day) with that of the simulated light from an Atlas CPS suntest machine (McGuire et al., 2000). They reported that the irradiance for UV wavelength (300-400 nm) of the natural sunlight ranged from
| Table 1: | Effect of simulated sunlight (at the rate of 500 W m-2 for 90 min) on mortality of second instars of H. armigera caused by HearNPV‡ when applied with and without adjuvants |
 | |
In each lot, means followed by the same letter are not significantly different (p = 0.05) by DMRT. ‡The virus concentration was 1x105 OB mL-1. xAll materials were compared with the irradiated virus alone as standard in the same lot and the results named as relative efficiency | |
4.0-29.3 W m-2 which was comparable with that of 24.3-26.5 W m-2 in suntest machine. For visible light (400-800 nm) the irradiance of the natural sunlight ranged from 75.5-487.8 W m-2 as compared to 321.2-384.6 W m-2 in suntest machine. Whereas the irradiance for the infrared (800-1100 nm) the irradiance of the natural sunlight ranged from 33.7-206.8 W m-2 as compared to 444.2- 480.4 W m-2 in the suntest machine. On the basis of this we selected an upper limit of 500 W m-2 as irradiance level to test the inactivation of virus by sunlight although it is known that only the UV wavelength inactivates the virus.
The irradiated virus was eluted immediately with 500 μL of distilled water and the suspension was collected in microfuge tubes, labeled and re-enumerated. Bioassays were performed with these irradiated viruses (1x105 OB mL-1) using second instars of H. armigera on semi-synthetic diet (Shorey and Hale, 1965). Ten microlitter of the suspension was dispensed on the semi-synthetic diet filled in 5 mL glass vials. Virus suspension was distributed uniformly over the entire diet surface using the polished blunt end of a glass rod (6 mm). The larvae after inoculation were incubated at 25 ± 1 °C in an incubator. Thirty larvae were used per treatment. Virus (alone) both irradiated and unirradiated served as control. In addition to this, untreated control was also maintained. All treatments were replicated thrice. Mortality was recorded daily after 3rd day for a period of 10 days.
Similar procedure was adopted to study the effect of combination of various adjuvants on the persistence of HearNPV.
Evaluation of adjuvants on tomato plants: Tomato seedlings (cv: All-round) were raised in pots. Ten microlitter of different treatments (Table 3) containing 1x105 OB mL-1 in 0.1% Teepol was smeared onto the 20 mm diameter area labeled with a marker pen on the upper surface of leaves. The suspensions were distributed uniformly by smearing with a polished blunt end of a glass rod (6 mm) and the leaves were exposed to open weather conditions. Virus with different adjuvants (alone and in combinations) was applied on the leaves in the similar fashion for 7 days, as a result of which on the 7th day, viral deposits exposed to weathering for 0, 1, 2, 3, 4, 5 and 6 days were available for studying the effect of various adjuvants on the persistence of virus on tomato leaves. On the 7th day, leaves from the treated plants were removed and placed individually with the treated surface upward. Ten newly moulted second instars of uniform age (3.45 ± 0.51 days) and weight (1.23 ± 0.18 mg) were released on each leaf. Then a sterilized glass vial (50x20 mm) was inverted on the treated area of the leaf thus confining the larvae and restricting their food consumption to the upper treated surface of the leaf. This was replicated thrice. After 24 h of feeding, the larvae were individually transferred to semi-synthetic diet lacking formaldehyde and plugged with sterile cotton. Mortality was recorded daily from the 3rd to the 10th day. The Relative Efficiency (RE) of each adjuvant was computed by:
This ratio can be used as a comparative measure to rate the efficacies of the adjuvants.
Data analysis: The data was subjected to Analysis of Variance (ANOVA) using SAS software version 6.12 and the means were separated by Duncan`s Multiple Range Test (DMRT). Probit analysis using Statistical Package for Social Sciences (SPSS) version 10.0 for Windows was carried out.
RESULTS
Laboratory evaluation of different adjuvants: Molasses (5.0%), CSKE (10%) and soya flour (10%) resulted in larval mortality of 91.7% indicating their efficiency in increasing kill as opposed to 51.3% with irradiated virus without any adjuvant. This was followed by Tinopal (0.2%), lampblack (1.0%), turmeric (2.0%), mango leaf extract (1.0%), boric acid (1.0%) and guava leaf extract (1.0%) resulting in a larval mortalities of 77.0-85.2% (Table 1). The maximum relative efficiency (1.8) was achieved with molasses (5.0%), CSKE (10%) and soya flour (10%). It was also observed that the unirradiated virus resulted in 98-99% larval mortality when compared to 50% of irradiated virus indicating the relative efficiency of unirradiated virus to be twice that of the irradiated virus. However, almost all the adjuvants that were tested resulted in increased larval mortality when compared to the control. The time-mortality responses of second instars of H. armigera revealed that all the adjuvants except food colour green, saffron and blue ink could significantly reduce the LT50. Lampblack (1.0%) had the shortest LT50 value (101.2 h) followed by turmeric (2.0%) and boric acid (1.0%), viz., 101.3 and 101.4 h, respectively (Table 2).
Considering cost and easy availability, different combinations of adjuvants along with HearNPV (Table 3) were tested for their efficacy in increasing the larval mortality of H. armigera. Bioassays conducted with different combinations of adjuvants revealed that molasses (5.0%) + Tinopal (0.2%) + lampblack (0.1%) showed the highest larval mortality (94.2%). However, crude sugar (5%) + Tinopal (0.2%) and molasses (5.0%) + Tinopal (0.2%) also resulted in a larval mortality of 93.9 and 93.8%,, respectively as compared to 52.3% with irradiated virus (Table 3).
All the adjuvants and their combinations significantly protected the virus when exposed to simulated sunlight. Molasses (5.0%) + Tinopal (0.2%) + lampblack (0.1%), crude sugar (5%) + Tinopal (0.2%) and molasses (5.0%) + Tinopal (0.2%) offered the maximum protection in terms of LT50 (Table 4).
Pot culture evaluation of different adjuvants for persistence of HearNPV on tomato: The activity of the virus applied without any adjuvant on tomato plants steadily decreased with time. The activity of the virus applied along with adjuvants also decreased with time but the rate of inactivation was significantly lower when compared to virus applied alone. From the second day onwards, significantly higher mortality rates were recorded when adjuvants were used either alone or in combination (Table 5).
| Table 2: | Time-mortality response of second instar larvae of H. armigera to HearNPV* applied with single adjuvants after exposure to simulated sunlight at 500 W m-2 for 90 min |
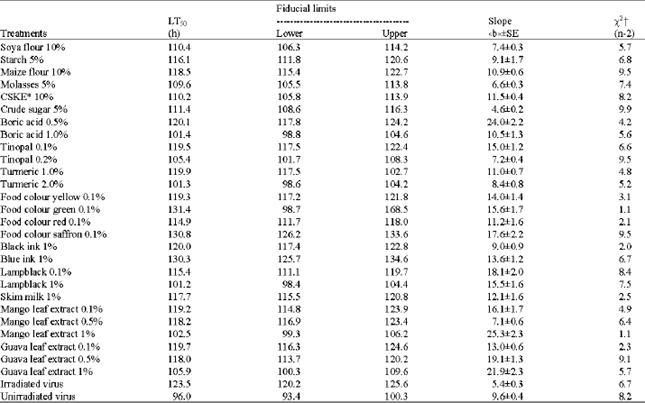 | |
| * The virus concentration was 1x105 OB mL-1. xAll lines are a significantly good fit (p<0.05) | |
| Table 3: | Evaluation of combined adjuvants on persistence of HearNPV after exposure to simulated sunlight at 500 W m-2 for 90 min against second instars of H. armigera |
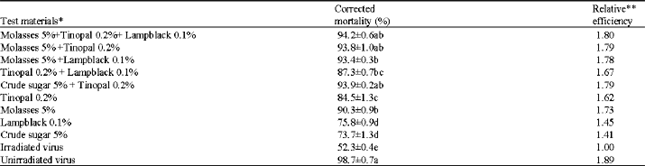 | |
*All the treatments contained NPV at 1x105 OB mL-1. **All materials were compared with the irradiated virus alone as standard and the results named as relative efficiency | |
A combination of molasses % + Tinopal 0.2% + lampblack 0.1% recorded highest OAR on all the days ranging from 85.94 to 36.27% for first and 6th days, respectively. Even though Tinopal 0.2% + Lampblack 0.1% recorded higher OAR 1 day after treatment but it was lower on all other days when compared with crude sugar 5% + Tinopal 0.2% which recorded higher OAR for the 2-6 days after treatment. On the 5th day only 28.1% of the Original Activity Remained (OAR) in virus applied alone, whereas in a combination of virus with molasses, lampblack and Tinopal, the OAR recorded was to the extent of 44.4% which was on par with molasses + Tinopal and crude sugar + Tinopal. The same trend was seen on the 6th day also. So using molasses 5% + Tinopal 0.2% + lampblack 0.1% increased the persistence of virus on all the days.
The LT50 recorded in different virus-adjuvant combinations was also significantly lower than in virus applied alone on all the 6 days (Table 6). In general, the combinations of adjuvants were found to be significantly better than single adjuvants.
| Table 4: | Time-mortality response of second instar larvae of H. armigera to HearNPV applied with combination of adjuvants after exposure to simulated sunlight at 500 W m-2 for 90 min |
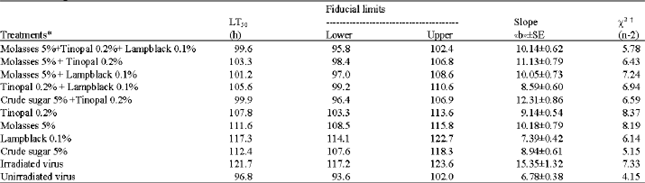 | |
| *All the treatments contained NPV at 1x105 OB mL-1. xAll lines are a significantly good fit (p<0.05) | |
| Table 5: | Persistence of HearNPV on tomato plants when applied with different adjuvants against second instar larvae of H. armigera |
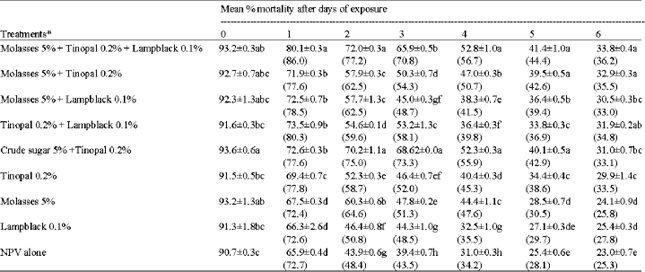 | |
*All the treatments contained NPV at 1x105 OB mL-1. Values in parenthesis represent percent original activity remaining. Means followed by the same letter in a column are not significantly different (p = 0.05) by DMRT | |
| Table 6: | Time-mortality response of second instar larvae of H. armigera to NPV applied with different adjuvants after days of application on tomato |
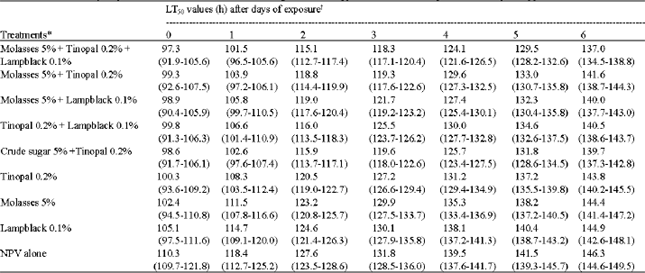 | |
*All the treatments contained NPV at 1x105 OB mL-1. xAll lines are a significantly good fit (p<0.05). Values in the parenthesis are representing the fiducial limits of each value | |
DISCUSSION
Of the adjuvants evaluated molasses (5.0%), cotton seed kernel extract (10%), soya flour (10%) and Tinopal (0.2%) significantly increased larval mortality. Turmeric (2.0%), lampblack (1.0%), boric acid (1.0%), mango leaf extract (1.0%) and guava leaf extract (1.0%) could also enhance larval mortality (Table 1). A combination of the highly promising adjuvants (5.0% molasses, 0.2% Tinopal and 0.1% lampblack) showed the highest larval mortality (94.2%) under simulated sunlight with the lowest LT50 values (99.6 h) (Table 3). The present findings that molasses increases the persistence of NPV corroborate with those of Smith et al. (1980) who reported an increase in potency of H. zea NPV when applied with molasses (10%) under simulated sunlight. Shapiro et al. (1983) also have reported that when molasses (5%) was used as adjuvant it offered UV protection to the virus. Farrar et al. (1995) increased gypsy moth larval feeding around 2-fold by using molasses (12.5%) on lettuce leaf discs, thereby increasing the acquisition of the virus and subsequent kill of the host insects.
Rabindra and Jayaraj (1989) reported that 4% larval extracts of H. armigera, Spodoptera litura and Corcyra cephalonica, 20% whole milk, 10% whole egg homogenate, 10% yellow of egg, 10% egg white, 20% tender coconut water and 20% crude sugar increased the efficacy of NPV against second instars of H. armigera. Rabindra and Jayaraj (1992) found that 10% water extracts of sorghum and pearl millet grains and sunflower seeds, cotton bolls and squares and tomato fruits significantly increased H. armigera larval mortality when added to HearNPV suspensions. Measurement of leaf area consumed showed that these adjuvants had acted as phagostimulants. In concurrence with these findings we also feel that in the present study molasses as well as crude sugar could have acted also as phagostimulants (in addition to act as UV protectant) thus increasing the uptake of virus which in turn increased the larval mortality.
Jacques (1971) tested 29 materials as UV protectants of Trichoplusia ni NPV either alone or in combination in the laboratory and under field conditions. He found that in the laboratory, addition of one of the following additives to the viral suspension could substantially prolong the persistence of the NPV, viz., India ink, charcoal, yeast extract, brewers` yeast, peptonized milk, soya hydrolyze, yellow stain or the autoclaved supernatant of a crude viral suspension. Addition of soybean, or cottonseed components and sugars to water-based formulations containing H. zea NPV produced significantly higher mortalities in laboratory tests against 24-h-old bollworms (Smith et al., 1982).
Tinopal, an optical brightener was found to enhance the persistence of HearNPV in the present study (Table 1). Fluorescent brighteners significantly lowered the LC50 and LT50 values in a variety of nucleopolyhedroviruses (Shapiro, 1992; Shapiro and Robertson, 1992; Webb et al., 1994; Shapiro and Dougherty, 1994; Shapiro and Argauer, 1995; Arivudainambi et al., 2000; Shapiro and Argauer, 2001; Murillo et al., 2003; Farrar et al., 2003; Dougherty et al., 2006). Also a combination of 5% molasses, 0.2% Tinopal and 0.1% lampblack was found to be the most effective in increasing the persistence of HearNPV with increased mortality of H. armigera and provided further kill in open weather conditions. Topper et al. (1984) found that combination of molasses (10%) + Teepol (0.5%) + Tinopal RBS200 (0.001%) was the best additive to enhance the field efficacy of S. littoralis NPV on cotton. One per cent leaf extracts of mango and guava were able to provide UV protection to the NPV of H. armigera in the present studies (Table 1). Polyflavonoids from mango and guava leaves was found to protect S. litura NPV from inactivation by sunlight (Asokan, 1998).
An adjuvant or a combination of adjuvants which provides UV protection increases the persistence of the virus. Deposition and persistence of the virus on the foliage as well as increased consumption of treated leaves would lead to improved kill of the larvae. Therefore, a proper choice of adjuvants would be crucial, particularly in terms of cost for successful use of these substances. Thus using molasses 5% + Tinopal 0.2% + lampblack 0.1% can be recommended as an effective adjuvant for management of H. armigera in tomato fields for increasing the persistence of virus on the plant which in turn will increase the larval mortality.
REFERENCES
- Arivudainambi, S., V. Selvanarayanan and A. Vikash, 2000. Enhancing the efficacy and persistency of Spodoptera litura (Fab.) nuclear polyhedrosis virus using UV irradiation protectants. Indian J. Exp. Biol., 38: 1175-1176.
Direct Link - Arthures, S.P., L.A. Lacey and R. Fritts, 2005. Optimizing use of codling moth granulovirus: Effects of application rate and spraying frequency on control of codling moth larvae in Pacific Northwest apple orchards. J. Econ. Entomol., 98: 1459-1468.
Direct Link - Behle, R.W., P. Tamez-Guerra and M.R. McGurie, 2003. Field activity and storage stability of Anagrapha falcifera NPV in spray-dried lignin-based formulations. J. Econ. Entomol., 96: 1066-1075.
Direct Link - Dougherty, E.M., N. Narang, M. Leob, D.E. Lynn and M. Shapiro, 2006. Fluorescent brightener inhibits apoptosis in baculovirus-infected gypsy moth larval midgut cells in vitro. Biocontrol Sci. Technol., 16: 157-168.
Direct Link - Farrar, Jr. R.R., R.L. Ridgway, S.P. Cook, K.W. Thorpe and R.E. Webb, 1995. Nuclear polyhedrosis virus of the gypsy moth (Lepidoptera: Lymantriidae): Potency and effects of selected adjuvants on insect feeding behavior. J. Entomol. Sci., 30: 417-428.
Direct Link - Farrar, R.R., M. Shapiro and I. Javaid, 2003. Photostabilized titanium dioxide and a fluorescent brightener as adjuvants for a nucleopolyhedrovirus. Biocontrol, 48: 543-560.
Direct Link - Griego, V.M., M.E. Martignoni and A.E. Claycomb, 1985. Inactivation of nuclear polyhedrosis virus (Baculovirus subgroup A) by monochromatic UV radiation. Applied Environ. Microbiol., 49: 709-710.
Direct Link - Jacques, R.P., 1971. Tests on protectants for foliar deposits of a polyhedrosis virus. J. Invertebr. Pathol., 17: 9-16.
CrossRef - Luttrell, R.G., W.C. Yearian and S.Y. Young, 1983. Effect of spray adjuvants on Heliothis zea (Lepidoptera: Noctuidae) nuclear-polyhedrosis virus efficacy. J. Econ. Entomol., 76: 162-167.
Direct Link - Martignoni, M.E. and P.J. Iwai, 1985. Laboratory evaluation of new ultraviolet absorbers for protection of Douglas-fir Tussock moth (Lepidoptera: Lymantriidae) baculovirus. J. Econ. Entomol., 78: 982-987.
CrossRefDirect Link - Martinez, A.M., P. Caballero, M. Villanueva, N. Miralles, I.S. Martin, E. Lopez and T. Williams, 2004. Formulation with an optical brightener does not increase probability of development resistance to Spodoptera frugiperda nucleopolyhedrovirus in the laboratory. J. Econ. Entomol., 97: 1202-1208.
Direct Link - McGuire, M.R., R.W. Behle, H.N. Goebel and T. Fry, 2000. Calibration of sunlight simulator for determining solar stability of Bacillus thuringiensis and Anagrapha falcifera nuclear polyhedrovirus. Environ. Entomol., 29: 1070-1074.
Direct Link - Murillo, R., R. Lasa, D. Goulson, T. Williams, D. Munoz and P. Caballero, 2003. Effect of Tinopal LPW on the insecticidal properties and genetic stability of the nucleopolyhedrovirus of Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol., 96: 1668-1674.
CrossRefDirect Link - Rabindra, R.J. and S. Jayaraj, 1988. Efficacy of nuclear polyhedrosis virus with adjuvants as high volume and ultra low volume applications against Heliothis armigera Hbn. on chickpea. Trop. Pest Manage., 34: 441-444.
CrossRefDirect Link - Rabindra, R.J. and S. Jayaraj, 1992. Efficacy of extracts of certain host plants as adjuvants for nuclear polyhedrosis virus of Helicoverpa armigera Hhn. and its dust formulation. J. Biol. Control, 6: 80-83.
Direct Link - Rabindra, R.J. and S. Jayaraj, 1995. Management of Helicoverpa armigera with nuclear polyhedrosis virus on cotton using different spray equipment and adjuvants. J. Biol. Control, 9: 34-36.
Direct Link - Shapiro, M., P.P. Agin and R.A. Bell, 1983. Ultraviolet protectants of the gypsy moth (Lepidoptera: Lymantriidae) nucleopolyhedrosis virus. Enviorn. Entomol., 12: 982-985.
CrossRefDirect Link - Shapiro, M., 1992. Use of optical brighteners as radiation protectants for gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus. J. Econ. Entomol., 85: 1682-1686.
CrossRefDirect Link - Shapiro, M. and J.L. Robertson, 1992. Enhancement of the gypsy moth (Lepidoptera: Lymantriidae) by an optical brightener. J. Econ. Entomol., 87: 361-365.
Direct Link - Shapiro, M. and E.M. Dougherty, 1994. Enhancement in activity of homologous and heterologous viruses against the gypsy moth (Lepidoptera: Lymantriidae) by an optical brightener. J. Econ. Entomol., 87: 361-365.
CrossRefDirect Link - Shapiro, M. and R. Argauer, 1995. Effects of pH, temperature and ultraviolet radiation on the activity of an optical brightener as a viral enhancer for the gypsy moth (Lepidoptera: Lymantriidae) baculovirus. J. Econ. Entomol., 88: 1602-1605.
CrossRefDirect Link - Shapiro, M. and R. Argauer, 2001. Relative effectiveness of selected stilbene optical brighteners as enhancers of the beet armyworm (Lepidoptera: Noctuidae) nuclear polyhedrosis virus. J. Econ. Entomol., 94: 339-343.
CrossRefDirect Link - Shorey, H.H. and R.L. Hale, 1965. Mass rearing of the larvae of nine noctuid species on a simple artificial medium. J. Econ. Entomol., 58: 522-524.
CrossRefDirect Link - Smith, D.B., D.L. Hostetter and R.E. Pinnell, 1980. Laboratory formulation comparisons for a bacterial (Bacillus thuringiensis) and a viral (Baculovirus heliothis) insecticide. J. Econ. Entomol., 73: 18-21.
CrossRefDirect Link - Smith, D.B., D.L. Hostetter, R.E. Pinnell and C.M. Ignoffo, 1982. Laboratory studies of viral adjuvants: Formulation development. J. Econ. Entomol., 75: 16-20.
CrossRefDirect Link - Topper, C., G. Moawad, D. McKinley, M. Hosny and K. Jones et al., 1984. Field trials with a nuclear polyhedrosis virus against Spodoptera littoralis on cotton in Egypt. Trop. Pest Manage., 30: 372-378.
CrossRefDirect Link - Webb, R.E., M. Shapiro, J.D. Podgwaite, R.R. Ridgway, L. Venables, G.B. White, R.J. Argauer, D.L. Cohen, J. Witcosky, K.M. Kester and K.W. Thorpe, 1994. Effect of optical brighteners on the efficacy of gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus in forest plots with high or low levels of natural virus. J. Econ. Entomol., 87: 134-143.
Direct Link