
Shafiqur Rahman
Bangladesh Fisheries Research Institute, Mymensingh-2201, Bangladesh
Mohd. Zakaria-Ismail
Institute of Biological Science, Faculty of Science, University of Malaya, 50603, Kuala Lumpur, Malaysia
Pek Yee Tang
Faculty of Engineering and Science, University Tunku Abdul Rahman, Malaysia
Sekaran Muniandy
Department of Molecular Medicine, Faculty of Medicine, University of Malaya, 50603, Kuala Lumpur, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 517-525







ABSTRACT
The genetic variability of wild populations of green arowana (Scleropages formosus) from Tasek Bera Lake and Endau River and a captive stock of Malaysian golden arowana as an out-group population was studied. The average number of alleles per locus was low, ranging from 2.33 in the Endau River population to 2.56 in the Tasek Bera Lake population. Departures from Hardy-Weinberg equilibrium were observed in these three populations depending on the locus. Among nine loci, seven significantly deviated from Hardy-Weinberg Equilibrium (p<0.05) in the Tasek Bera Lake population whereas five significantly departed from HWE (p<0.05) in the Endau River population. A deficit of heterozygosity was observed from the analysis of these nine loci. This genetic data indicate the wild population may be under stress due to inbreeding, over fishing and deterioration in the breeding environment. The estimates of FST and RST were statistically greater than zero for each comparison and this suggested the existence of three genetically distinct populations. However, the values of FST is low between Tasek Bera Lake and Endau River populations, reflecting historical patterns of connections between river systems in Peninsular Malaysia.
PDF Abstract XML References Citation
How to cite this article
Shafiqur Rahman, Mohd. Zakaria-Ismail, Pek Yee Tang and Sekaran Muniandy, 2008. Microsatellite Analysis of Wild and Captive Populations of Asian Arowana (Scleropages formosus) in Peninsular Malaysia. Journal of Biological Sciences, 8: 517-525.
DOI: 10.3923/jbs.2008.517.525
URL: https://scialert.net/abstract/?doi=jbs.2008.517.525
DOI: 10.3923/jbs.2008.517.525
URL: https://scialert.net/abstract/?doi=jbs.2008.517.525
INTRODUCTION
The river systems in Malaysia are rich in flora and fauna. Tasek Bera Lake is a lowland alluvial riparian swamp system, in central Peninsular Malaysia (Fig. 1). It lies within the catchments of Pahang River, the Peninsula`s largest river. This wetland is nominated Malaysian`s first RAMSAR site of November 1994. It is an excellent example of a blackwater ecosystem, which includes open water, a reed swamp area and swamp forest with grasslands on the periphery. The lake contains a large number (95 species of fish) of the country's freshwater fish species, including both popular aquarium and sport fishes most of which are indigenous and include the endangered Asian arowana, Scleropages formosus (Sim, 2002). The Endau-Rompin area is situated at the boundary of Johor and Pahang States in Peninsular Malaysia (Fig. 1). It is one of the few remaining lowland forests in Peninsular Malaysia. It encompasses the watershed of the Endau and Rompin rivers. The Endau River, flows across the park in an easterly direction and drains into the South China Sea. Water flow is more or less smooth and the water is less transparent than in streams. A total 108 species in 26 families have been reported to be present in the Endau basin (Ng and Tan, 1999). It was also reported that the upstream portion of Endau River is the breeding ground of the arowana (Zakaria-Ismail, 1987).
The Asian arowana (Scleropages formosus) is also known as dragonfish, Asian bonytongue, Kelisa or Baju rantai. This ancient Ossteoglossid fish is one of the most expensive and sought after fish in aquatic world. The Asian arowana, has been reported in Cambodia, Thailand, Malaysia, Borneo and Sumatra (Pouyaud et al., 2003). There are three main colour varieties (green, gold and red). The green and gold varieties are found in Peninsular Malaysia (Tang et al., 2004). The green variety is still relatively common in some areas such as Tasek Bera Lake in Pahang, Endau River in Johor (Ng and Tan, 1999; Sim, 2002) and Trengganu drainage area (Cramphorn, 1983). It is believed that the wild populations of the golden variety are almost extirpated due to over
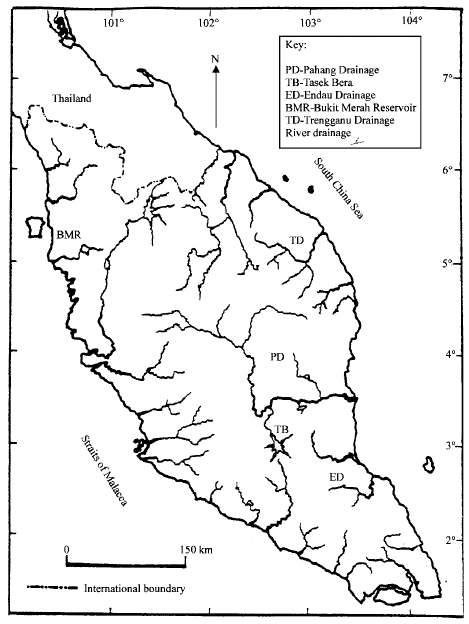 |
| Fig. 1: | Major river drainages and major habitats of arowana (Scleropages formosus) in Peninsular Malaysia |
harvesting and habitat modification. The arowana has been listed as one of most highly endangered fish by CITES the Convention on International Trade in Endangered Species of Wild Fauna and Flora (Greenwood et al., 1996). Arowana is threatened with extinction and placed on the Red List (Hilton-Taylor, 2000) of the International Union for Conservation of Nature and Natural Resources (IUCN). Only the F2 generation from the commercially captive-bred individuals can legally be exported. The survivability of the arowana is further threatened due to some of its natural behavior of low fecundity rate, oral-brooding habit and open-water spawning (Dawes, 1999).
To conserve such a declining species, it is necessary to have sound knowledge on its biology, ecology, biogeography and inter-population genetic diversity. From the genetic perspective, the aim of natural fishery management should be to conserve intra-specific genetic diversity.
DNA fingerprints of arowana have been studied by different methods to determine which methods were most suitable in providing information on genetic variability. As the DNA fingerprint is a pattern made up of DNA fragments that are resolved by electrophoresis, each individual has its own unique fingerprint due to its genetic make-up. The capability of Restriction Fragment Length Polymorphism (RFLP) and random amplified polymorphic DNA (RAPD) techniques to asses genetic variability of arowana is low (Fernando, 1997). Analysis using Amplified Fragment Length Polymorphism (AFLP) have shown significant genetic differentiation between the green, red and gold arowana (Yue et al., 2002, 2004).
The use of microsatellites is well documented in the study of fish genetics (Sekino et al., 2002; Was and Wenne, 2002; Elliott and Reily, 2003). It is a powerful technique to detect genetic variability and can be used to assess variability between and within captive strains of arowana (Yue et al., 2000, 2004). However, genetic variation in wild population of arowana has yet to be studied. In this study, we used microsatellite markers to assess genetic variability of wild populations of green arowana in the Tasek Bera Lake and Endau River in comparison with a captive population of golden arowana as an out-group. Work such as this will considerably help to protect the wild populations of arowana as the microsatellite DNA fingerprint provides undisputed and reproducible evidence of relatedness and population identification. This information can be applied for long-term management of wild population as well as captive population through analysis of their pedigree and heterozygosity and provide guidelines for captive breeding.
MATERIALS AND METHODS
Fish samples collection: A total of 135 individuals were obtained from both of the habitats Tasek Bera Lake and Endau River. The locations selection was based on where the aboriginal people catch arowana fishes. Ninety-seven green arowana were collected from Tasek Bera Lake, Pahang and thirty-eight green arowana were sampled from Endau River. Fishermen used dugout canoes to scoop individuals after dark. The fish were located from the reflection of theirs eyes in torchlight. Initially all the collections were kept in polystyrene basins for several days. When the catch reached 10 individuals, the fish were shifted to the Ecology and Biology Laboratory, Faculty of Science, University of Malaya and where it`s reared individually in aquarium. Twenty-three Malaysian golden arowana samples were collected from the Malacca Fisheries Department as an out-group population for comparison.
Tissue sample collection: Scales, over turned gill covers, fin clips and muscle tissues sample were used for DNA extraction. Scales were either removed from live specimens or were from accidental loss while over turned gill cover and fin clips were only from live specimen. Muscles were collected only from dead. These were preserved in either a laboratory standard bottle or a 1.5 mL micro-centrifuge tubes containing 90% ethanol until genomic DNA extraction was performed. All samples were stored at -20 °C.
DNA extraction and PCR: The method of DNA extraction from the preserved tissues was modified from Asahida et al. (1996). Approximately 20-30 mg of specimen tissue was used per extraction in 260 μL TNES-Urea buffer (6 M Urea; 10 mM Tris-HCl, pH 7.5; 125 mM NaCl; 10 mM EDTA; 1% SDS) and 0.2 mg (muscle) to 0.8 mg (scale, fin and overturned gill cover) of Proteinase K was added to the samples. The mixture was incubated for 12 h (muscle tissue) or 15 h (scale, fin and overturned gill cover) at 55 °C followed by conventional phenol-chloroform extraction.
Genetic variation within and between these populations was assessed using nine microsatellite loci. Among these markers four were described by Yue et al. (2000), while five loci were isolated by Tang et al. (2004). The Polymerase Chain Reaction (PCR) procedure was performed on a Hybaid Omnigene thermal cycler in total a volume of 25 μL. Reactions contained 1x PCR buffer (Promega), 1.5 mM MgCl2, 200 μM of each dNTP, 0.2 μM of each primer, 1U Taq polymerase (Promega) and 20 ng of genomic DNA. Amplifications for the four loci described by Yue et al. (2000) were carried out using 4 min of initial denaturation followed by 30 cycles of 30 sec of denaturation at 94.5 °C, 30 sec annealing at the temperature detailed in Table 1 and 30 sec extensions at 72 °C, with a final extension of 5 min at 72 °C. The other five loci were amplified with 3 min of initial denaturation followed by 40 cycles of 10 sec of denaturation at 95 °C, annealing at specific temperature (Table 1) for 10 and 30 sec extensions at 72 °C, with a final extension of 5 min at 72 °C. Polymerase Chain Reaction (PCR) products were run on a 10% nondenaturing polyacrylamide gel (16H20 cm) at 250 V for 4-5 h. A 20 bp DNA marker (Cambrex BioScience) was used to estimate the PCR fragment size. Gels were visualized using DNA silver staining system (Promega) and analyzed using GelCompar the II Software (Applied Maths).
Statistical analysis: Microsatellite allele frequencies of each population at each locus were estimated using GENEPOP version 3.1c (Raymond and Rousset, 1995). The observed (Ho) and expected heterozygosities (He), of the polymorphic loci of each strain were estimated using ARLEQUIN version 2.000 (Schneider et al., 2001). Hardy-Weinberg equilibrium (HWE) at each locus was assessed using ARLEQUIN. A Markov-chain method with 1000 steps, 1000 dememorizations was used to calculate an unbiased estimate of the p-value. The inbreeding coefficient, FIS (Weir and Cockerham, 1984) was estimated using the GENEPOP programme to measure the HWE departures for each population. The software MICROCHECKER (Van Oosterhout et al., 2004) using the
| Table 1: | Primers sequences of 9 microsatellite loci |
 | |
Brookfield`s (1996) method was used to identify the presence of null alleles. MICROCHECKER was also used to test for another source of errors in mis-scoring due to stuttering and allelic dropout. Population differentiation was measured by calculating pairwise weighted FST (Weir and Cockerham, 1984) values over seven loci (D92, D27, D85, K10, K13, K16 and K27) using ARLEQUIN. RST values were also calculated as sum of squared size differences based on number of repeats (Slatkin, 1995). The probability associated with FST and RST values was estimated through random permutation procedure (1000 permutation).
RESULTS
Microsatellite variability: All nine microsatellite loci were polymorphic and exhibited variability in the three populations. Allele frequency distributions for these populations are listed in Table 2. A total of 40 alleles were detected in 158 individuals. The average number of alleles per locus ranged from 2.33 in the Endau River population to 2.78 in the Malaysian golden population. Alleles D92116, D2776 and K13206 were found in the Tasek Bera Lake population but undetected in the Endau River population. Allele K13202 was only detected in the Endau River and the Malaysian golden population. Fifteen alleles were only present in the Malaysian golden population while 15 alleles were only present in the Tasek Bera Lake and the Endau River populations. Most of the alleles found in this study showed high frequencies and rare alleles were not detected.
Hardy-Weinberg equilibrium: The Malaysian golden population showed the highest average observed heterozygosity (0.522), followed by the Tasek Bera Lake (0.301) and the Endau River (0.304) populations. The average expected heterozygosity at the nine loci was highest (0.407) in the Malaysian golden population, followed by the Endau River population (0.405) and the Tasek Bera Lake (0.368) population. Positive values of inbreeding coefficient (FIS) were observed for most of the loci across all the 3 populations. The highest average FIS value (0.311) was observed in the Endau River population, whereas the lowest (-0.256) was seen in the Malaysian golden population. The data implicate inbreeding in the Tasek Bera Lake and the Endau River populations (Table 3).
Among the nine loci screened in the Tasek Bera Lake population, seven deviated significantly (p<0.05) from Hardy-Weinberg equilibrium. Five out of nine loci from the Endau River-populations departed significantly from HWE (p<0.05). Five loci in the Malaysian golden population were deviated (p<0.05) from HWE (Table 3). MICROCHECKER indicated that departure from HWE in these three populations was not due to stuttering or large allele drop out. Departure from HWE at loci D11 and K37 in the Tasek Bera Lake and Endau River populations was contributed by null alleles. The frequencies of null alleles were 0.33 for D11, 0.17 for K37 in the Tasik Bera Lake population and 0.32 for D11, 0.09 for K37 in the Endau River population.
Genetic differentiation: Pairwise FST values indicate that there is significant genetic differentiation (p<0.05) between these three populations (Table 4). The Tasek Bera Lake and the Endau River populations were the most similar. The population differentiations based on FST and RST values were similar but RST values were higher than FST values.
| Table 2: | Allele distributions for nine microsatellite loci in three populations of arowana |
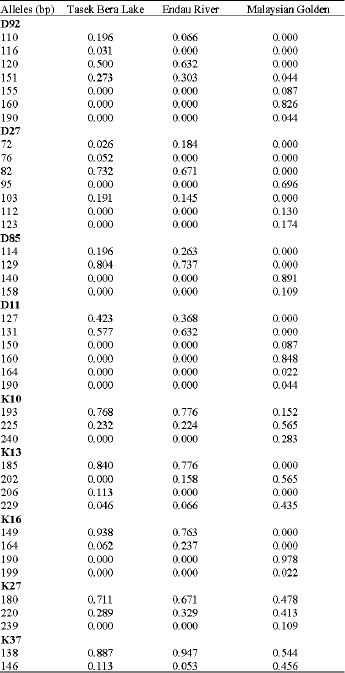 | |
| Table 3: | Number of alleles (N), observed heterozygosity (Ho), expected heterozygosity (He) and number of studied specimens (n) at nine microsatellite loci for three populations of arowana |
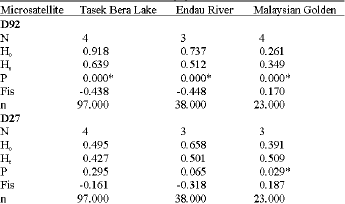 | |
 | |
*: Indicates a significant deviation from Hardy-Weinberg equilibrium, p<0.05 | |
| Table 4: | Pairwise comparisons of microsatellite FST (below diagonal) and RST (above diagonal) between 3 populations of arowana |
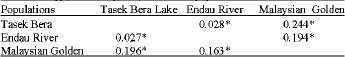 | |
| *: Indicates a significant genetic distance, p<0.05) | |
DISCUSSION
Deviation from Hardy-Weinberg equilibrium: In this study, most of the loci screened deviated from HWE. The observed heterozygosities observed in our study were lower than estimates by Yue et al. (2000, 2004) but comparable to estimates by Sivananthan (2004) and Tang (2004). This may be attributed to the territorial behaviour of arowana in nature (Scott and Fuller, 1976) where the entire population may be divided into several subpopulations and mating between related individuals is more common. Thus, inbreeding increases the proportion of homozygous individuals in a population. Positive values of inbreeding coefficient (FIS) further supported possibility of inbreeding in the Tasek Bera Lake and the Endau River Populations. The presence of positive values of inbreeding coefficient (FIS) could also be caused by the presence of null alleles which has also been reported in a previous study of arowana (Tang, 2004). Bentzen et al. (1996) suggested null alleles might be responsible for significant deficiency of heterozygotes. An alternative explanation is that although the individuals were sampled as far as possible, they may not represent the true population due to the natural behavior of the arowana. The arowana is territorial in nature and fry could be collected in the mouth of the males or are in the vicinity of adults. Thus, individuals collected at the same location might often be siblings produced by a relatively few adults.
Microsatellite variability: The distribution of alleles per locus in this study is less than Yue et al. (2000, 2002, 2004) but there is considerable overlap in the distribution. The present result showed that there was a lack of private alleles to differentiate the two green arowana populations. Three private alleles were detected, 2 in the Tasik Bera Lake population and 1 in the Endau River population. Individuals which carried this allele might not be sampled there or there might not be such an allele. Low sample sizes can impact the precision of estimates of population structure and genetic distance. However, the sample size of the Tasek Bera Lake population fell within the range recommended by O'Connel and Wright (1997) and was higher than the sample size used by Yue et al. (2000, 2002, 2004) but this population still displayed a very low level of genetic variability. Thus, suboptimal sample size should not be the main reason for the over all low genetic variability in this study. However, sample size of the Endau River population was lower than that recommended. Furthermore, this comparison is considered biased because in the present study all of the samples were collected from single region while the green arowana stock studied by Yue et al. (2000) was pooled from farms in Singapore and Indonesia. The study carried out by Yue et al. (2004) was based on stock established from wild-caught arowana in the early 1980s and this might not represent the recent natural populations. Low number of allele was also recorded in a green arowana stock with unknown pedigree obtained from the Malacca Fisheries Department (Sivananthan, 2004; Tang, 2004).
The numbers of alleles were found to be lower than those reported by other authors in other fish species (Naish and Skibinski, 1998; McConnell et al., 2001). Loss of allelic variation also has been reported for Danish and Polish populations of brown trout (Hansen et al., 2000; Was and Wenne, 2002). Tessier et al. (1997) observed significant changes in allele frequencies and losses of low-frequency alleles, but no reduction in heterozygosity of wild Atlantic salmon populations. Reduction in allelic variation in the present study of arowana in wild populations may be explained by the founder effect and genetic drift (Alam and Islam, 2005). It may thus be assumed that the populations might have a small effective number of parents (Ne) as illegal fishing is an ongoing activity at Tasek Bera Lake and Endau River even though the arowana is protected under the State Fisheries Enactment, 1991. The estimates of effective population size were very low for a natural populations (Rahman, 2007) compared with captive populations (Yue et al., 2004). Thus, the losses of alleles and heterozygosity may increase with bottlenecking and inbreeding in the populations and it can be postulated that the wild population may be under stress due to over fishing and deterioration in the breeding environment. Populations that have recently undergone bottlenecks are likely to lose rare alleles severely but may contain substantial amounts of heterozygosity (Nei et al., 1975; Allendorf, 1986). Our results showed that no rare allele was detected in these three populations.
A low fecundity rate further threatens the population size of the arowana. Reduction in genetic diversity had been linked to decreases in growth and fecundity, changes in sex ratio and the ability to adapt to environmental changes (Chapman et al., 1999). Maintenance of genetic variation is essential for long-term survival of populations. However, stock transfer will be a difficult task. To date, no survey has been carried out to determine the genetic diversity of the wild populations in other river systems such as the Kahang River (Endau Drainage; Fig. 1) and Kenyir Lake (Terengganu Drainage; Fig. 1). Furthermore, local adaptation will be lost if the population is mixed with others (Haig, 1998). Another source of diversity will be the populations in captivity. In captivity it is possible to develop this kind of diversified stocks by selection and breeding and this had been reported for captive populations of arowana by Yue et al. (2000, 2002, 2004). Through genetically compatible introductions, some individuals from the hatcheries can be released to their natural habitat to improve the gene pool (Philipp et al., 1993). However since, the green arowana fetches the lowest price compared to the gold and red varieties, its breeding programme in the private farms is not extensive. Since the captive breeding involves selection, it must be noted that the adverse genetic effects of stock transfer such as outbreeding depression (Templeton et al., 1986; Philipp, 1991) should be taken into consideration.
Genetic differentiation: Our estimates of FST and RST were statistically greater than zero for each comparison and this suggested the existence of three genetically distinct populations. However, the value of FST is low between Tasek Bera Lake and Endau River. The pairwise genetic differentiation provide evidence of underlying geographic and temporal components of population divisions between these three populations. The Tasek Bera Lake and the Endau River populations were collected from two geographically distant drainages but FST suggest that gene flow is high. The effect of geographical distance on FST and gene flow (Nm) values has been reported in stream-living brown trout collected from different sections of the main stream (Carlsson et al., 1999) and Indian major carp Catla catla collected from three major rivers (Alam and Islam, 2005). However, in this study, these two wild populations showed a close relationship reflecting historical patterns of connections between river systems in Southeast Asia (Voris, 2000). Heist et al. (1996) attributed that relative genetic homogeneity between populations of different regions does not always signify that separate fishery stocks do not exist; reasons for this are historical associations and minimal gene flow. The populations which may not have been isolated long enough to established differences in allele frequencies, or exchange of individuals between populations, even small population size is sufficient to maintain the same alleles in different populations and contribute to allele genetic similarity between in the same alleles in different populations.
CONCLUSION
Information on the genetic structure of natural fish species is essential for optimizing fisheries management and stock improvement programmes. The study revealed a relatively low level of genetic variation at microsatellite loci within and between arowana populations, with genetic variation in the natural population lower than the captive populations. Knowledge of genetic structure of the wild as well as captive arowana populations is indispensable for management of the populations in order to maintain their genetic quality.
ACKNOWLEDGMENTS
This study was supported by short-term research grants 0139/2002A, 0142/2003A and 0131/2004A from the University of Malaya and the research was partly funded by a research grant from the Ministry of Science, Technology and Innovation, Malaysia (05-01-03-SF0172). We wish to express our sincere thank to the Malaysian Government for granting the sponsorship through the Malaysian Technical Cooperation Programme (MTCP) for the study.
REFERENCES
- Alam, M.S. and M.S. Islam, 2005. Population genetic structure of Catla catla (Hamilton) revealed by microsatellite DNA markers. Aquaculture, 246: 151-160.
CrossRefDirect Link - Allendorf, F.W., 1986. Genetic drift and the loss of alleles versus heterozygosity. Zoo Biol., 5: 181-190.
CrossRefDirect Link - Asahida, T., T. Kobayashi, K. Saitoh and I. Nakayama, 1996. Tissue preservation and total dna extraction form fish stored at ambient temperature using buffers containing high concentration of urea. Fish. Sci., 62: 727-730.
CrossRef - Bentzen, P., C.T. Taggart, D.E. Ruzzante and D. Cook, 1996. Microsatellite polymorphism and the population structure of Atlantic cod (Gadus morhua) in the Northwest Atlantic. Can. J. Fish. Aquat. Sci., 53: 2706-2721.
CrossRef - Brookfield, J.F.Y., 1996. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol., 5: 453-455.
CrossRef - Carlsson, J., K.H. Olsen, J. Nilsson, O. Oerli and O.B. Stabell, 1999. Microsatellites reveal fine-scale genetic structure in stream-living brown trout. J. Fish Biol., 55: 1290-1303.
CrossRef - Elliott, N.G. and A. Reily, 2003. Likelihood of bottleneck event in the history of the Australian population of Atlantic salmon (Slamo salar L.). Aquaculture, 215: 31-44.
Direct Link - Fernando, A.A., 1997. DNA fingerprinting: Application to conservation of the CITES-listed dragon fish, Scleropages formosus (Osteoglossidae). Aquarium Sci. Conserv., 1: 91-104.
CrossRef - Hansen, M.M., E.E. Nielsen, D.E. Ruzzante, C. Bouza and K.L.D. Mensberg, 2000. Genetic monitoring of supportive breeding in brown trout (Salmo trutta L.) using microsatellite DNA markers. Can. J. Fish. Aquat. Sci., 57: 2130-2139.
CrossRef - Heist, E.J., J.A. Musick and J.E. Graves, 1996. Mitochondrial DNA diversity and divergence among sharpnose sharks, Rhizoprionodon terraenovae, from the Gulf of Mexico and Mid-Atlantic bight. Fishery Bull., 94: 664-668.
Direct Link - McConnell, S.K.J., J. Leamon, D.O.F. Skibinski and G.C. Mair, 2001. Microsatellite markers from the Indian major carp species, Catla catla. Mol. Ecol. Notes, 1: 115-116.
Direct Link - Naish, K.A. and D.O.F. Skibinski, 1998. Tetranucleotide microsatellite loci for Indian major carp. J. Fish Biol., 53: 886-889.
CrossRef - Nei, M., T. Maruyama and R. Chakraborty, 1975. The bottleneck effect and genetic variability in populations. Evolution, 29: 1-10.
Direct Link - O'Connell, M. and J.M. Wright, 1997. Microsatellite DNA in fishes. Rev. Fish. Biol., 7: 331-363.
CrossRef - Philipp, D.P., J.M. Epifanio and M.J. Jennings, 1993. Conservation genetics and current stocking practices-are they compatible. Fish. Sci., 18: 14-16.
Direct Link - Pouyaud, L., Sudarto and G.G. Teugels, 2003. The different colour varieties of the Asia arowana Scleropages formosus (Osteoglossidae) are distinct species: Morphologic and genetic evidences. Cybium, 27: 287-305.
Direct Link - Scott, D.B.C. and J.D. Fuller, 1976. The reproductive biology of Scleropages formosus (Muller and Schlegel) (Osteoglossomorpha, Osteoglossidae) in Malaya and the morphology of its pituitary gland. J. Fish Biol., 8: 45-53.
CrossRefDirect Link - Sekino, M., M. Hara and N. Taniguchi, 2002. Loss of microsatellite and mitochondrial DNA variation in hatchery strains of Japanese flounder Paralichthys olivaceus. Aquaculture, 213: 101-122.
Direct Link - Slatkin, M., 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics, 139: 457-462.
PubMed - Tessier, N., L. Bernatchez and J.M. Wright, 1997. Population structure and impact of supportive breeding inferred from mitochondrial and microsatellite DNA analyses in land-locked Atlantic salmon Salmo salar L. Mol. Ecol., 6: 735-750.
CrossRefDirect Link - Van Oosterhout, C., W.F. Hutchinson, D.P.M. Wills and P. Shipley, 2004. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes, 4: 535-538.
CrossRefDirect Link - Voris, H.K., 2000. Maps of Pleistocene sea levels in Southeast Asia: Shorelines, river systems and time durations. J. Biogeogr., 27: 1153-1167.
CrossRefDirect Link - Was, A. and R. Wenne, 2002. Genetic differentiation in hatchery and wild sea trout (Slamo trutta) in the Southern Baltic at microsatellite loci. Aquaculture, 204: 493-506.
Direct Link - Weir, B.S. and C.C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358-1370.
CrossRefDirect Link - Yue, G.H., F. Chen and L. Orban, 2000. Rapid isolation and characterization of microsatellites from the genome of Asian arowana (Scleropages formosus, Osteoglossidae, Pisces). Mol. Ecol., 9: 993-1011.
Direct Link - Yue, G.H., Y. Li, F. Chan, S. Chao, L.C. Lim and L. Orban, 2002. Comparison of three DNA marker systems for assessing genetic diversity in Asian arowana (Scleropages formosus). Electrophoresis, 23: 1025-1032.
Direct Link - Yue, G.H., Y. Li, L.C. Lim and L. Orban, 2004. Monitoring the genetic diversity of three Asian arowana (Scleropages formosus)captive stocks using AFLP and microsatellites. Aquaculture, 237: 89-102.
Direct Link