
Z.A. Intan Zarina
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Selangor, Malaysia
Z.A. Shahrul Hisham
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Selangor, Malaysia
M.A.W. Rohaya
Department of Orthodontics, Faculty of Dentistry, Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia
S. Sahidan
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Selangor, Malaysia
Z.A. Zaidah
Department of Microbiology, Faculty of Applied Sciences, Universiti Teknologi Mara, Selangor, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 506-516







ABSTRACT
The aim of this study is to differentiate peripheral blood mononucleated cells that have been isolated from ICR mice`s (Mus musculus)peripheral blood into mature osteoblast and osteoclast cells with addition of growth factors. The mononucleated cells were isolated by density centrifugation using Ficoll-PaqueTM Plus. The culture mediums were then supplemented with growth factors; 50 ng mL-1 RANKL and 25 ng mL-1 M-CSF to differentiate into osteoclast cells. On the other hand, osteoblast assay, 50μg mL-1 ascorbic acid and 10 mM β-glycerophosphate were added to support differentiation. For control, the same cells were used without supplementation of respective growth factors. Biochemical analysis for osteoblast, i.e., Alkaline Phosphatase (ALP) activity was determined on day 3-14. The results showed that the ALP activity in the differentiation medium is significantly increased (p<0.05) on day 10 and day 14 as compared to control, i.e., cells without growth factors. Tartrate Resistant Acid Phosphatase (TRAP) activity that represented as osteoclast biomarker also showed significant increased (p<0.05) from day 3 until day 10 in the present of RANKL and M-CSF. The viability of differentiated cells also showed that the cells were able to survive until 10 to 14 days in the presence of respective growth factors without significant increased in respective differentiation medium. RT-PCR analysis on isolated RNA from mononucleated cells after 14 and 10 days in their differentiation medium showed that the osteoblast and osteoclast were expressing ALP (~373 bp) and TRAP (~281 bp) gene, respectively. Mononucleated cells originated from peripheral blood have the potential to differentiate into osteoblast and osteoclast cells in the presence of specific growth factors. The respective cells are primitive enough to differentiate into two distinct types of mature cells hence can be categorized as multipotent stem cells.
PDF Abstract XML References Citation
How to cite this article
Z.A. Intan Zarina, Z.A. Shahrul Hisham, M.A.W. Rohaya, S. Sahidan and Z.A. Zaidah, 2008. Osteoclast and Osteoblast Development of Mus musculus Haemopoietic Mononucleated Cells. Journal of Biological Sciences, 8: 506-516.
DOI: 10.3923/jbs.2008.506.516
URL: https://scialert.net/abstract/?doi=jbs.2008.506.516
DOI: 10.3923/jbs.2008.506.516
URL: https://scialert.net/abstract/?doi=jbs.2008.506.516
INTRODUCTION
Stem cell is categorized into two types: adult and embryonic stem cells. The most primitive cell such as embryonic stem cell is totipotent cells followed by pluripotent and multipotent cells types. Although multipotent cells are the most differentiated stage of stem cells development, the total number of these cells in a tissue is still extremely low. The haemopoietic stem cells in adult bone marrow for example, comprise of only 0.01-0.05% of total bone population (Shahrul Hisham et al., 2005).
Embryonic stem cells originate from the inner cell mass of the blastocyst stage during embryonic development, whereas adult haemopoietic stem cells are derived from bone marrow (Shahrul Hisham et al., 2005). Embryonic Stem (ES) cells are primitive cells represent embryonic precursor cells that give rise to any cell type in the embryo. Embryonic stem cells were first identified in 1981 as cell populations capable of differentiating into cell types of all embryonic germ layers (Martin, 1981). The cells were derived from the Inner Cell Mass (ICM) of the blastocyst. The embryonic stem cells would differentiate into specific cells such as skeletal muscle (Wakitani et al., 1995), endothelium (Yamashita et al., 2000), chondrocytes (Kramer et al., 2000), cardiac muscle (Orlic et al., 2001), blood (haemopoietic) cells (Spangrude et al., 1991), osteoblast (Buttery et al., 2001) and adipocytes (Dani et al., 1997) with specific medium and growth factors.
Adult stem cells are not lineage restricted. These cells are able to cross lineage boundaries and differentiate into different types of cells or tissue. These type of cells able to replace senescent cells or restore the lost function of cells due to disease or trauma (Sadiq and Gerber, 2004), such as hemopoietic stem cells that able to give rise to multiple haemopoietic phenotype cells (Baum et al., 1992), CNS-derived stem cells in producing neuronal cells (Kondo and Raff, 2000; Gritti et al., 1999) and intestinal stem cells that develop into multiple cell types in the gut (Bjerknes and Cheng, 1999).
Therefore, both adult and embryonic stem cells can be defined as undifferentiated cells that capable to proliferate and differentiated into more than one specialized cell type (Weissman, 2000). The potential degree of cellular differentiation varies among stem cell population has led to their categorization. This cell population can be categorized into three categories as totipotent, pluripotent and multipotent (Shahrul Hisham et al., 2005). Totipotent cells which originated from a fertilized egg comes from the Latin word totus, meaning entire, because it has the potential to generate into all different types of cells in the body and capable to develop into one complete organism. Pluripotent stem cells are those that can give rise to any type of cell in the body except those needed to develop a fetus. Pluripotent cells give rise to cells derived from three Embryonic Germs (EG) layers, i.e., endoderm, mesoderm and ectoderm (Shanthly et al., 2006). Whilst, multipotent cells are stem cells that can give rise to a small number of different cell types associated with different organs.
The multipotent cells type is the most differentiated stage of stem cells. The respective cells are more committed and give rise to lineage-restricted tissue specific cell types (Shahrul Hisham et al., 2005). The respective cells is different compared to progenitor cells as multipotent cells have the potential to develop into two types of cells or more whereas, progenitor cells are committed into only one matured cells. Mature blood cells have a limited life-span and are continuously replaced by the proliferation and differentiation of a very small population of pluripotent haemopoietic stem cells found primarily in the healthy adults Bone Marrow (BM). Haemopoietic stem cells have the ability to replenish themselves and to differentiate into progenitor cells and mature blood cells of all haemopoietic lineages.
Current reports show that adult stem cells are not lineage restricted and able to differentiate into various cells or tissue. This phenomenon also known as metaplasia, i.e., define as conversion of one cells or tissue type to another. Studies showed that haemopoietic stem cells derived from bone marrow are capable of transdifferentiating into liver hepatocytes (Theise et al., 2000a, b; Lagasse et al., 2000; Krause et al., 2001; Fujii et al., 2002), neurons (Brazelton et al., 2000) and cardiac myocytes (Orlic et al., 2001). On the other hand, neural progenitor cell can differentiate into muscle cells (Galli et al., 2000) and transdifferentiation of liver stem cells into pancreatic (Yang et al., 2002) and cardiac phenotypes (Malouf et al., 2001).
The term transdifferentiation or de-differentiation was originally invented by developmental biologists to describe the ability of apparently fully differentiated embryonic cells of one tissue type to become cells of a different tissue type in response to in vitro cell culture (Asashima and Okada, 2001). Transdifferentiation currently used to describe a similar behavior attributed to stem cells in postnatal somatic tissues, i.e., adult stem cells. A few studies suggest that adult stem cells originated in one tissue when transferred to a different tissue environment, they are able to produce cells with gene expression properties characteristic of cells in other tissues (Watt and Hogan, 2000; Morrison, 2001; Clarke and Frisén, 2001).
In present studies, we managed to isolate primitive mononucleated cells from peripheral blood cells using Ficoll-Paque cell density isolation (Forraz et al., 2004; Pearce et al., 2005). The quantity of the respective cell was increased in a standard cells proliferation condition for 30 days before been subjected into separated differentiation assay of osteoclast and osteoblast for another 10 to 14 days. Present result showed that the mononucleated cells isolated from mouse peripheral blood have potential to develop into more than one type of mature cells. The respective cell was shown could be partially differentiated into osteoclast and osteoblast cells. The osteoclast and osteoblast cells were determined using biochemical and molecular analysis with their respective biomarkers.
MATERIALS AND METHODS
The studies were conducted from September 2005 until November 2007 at Laboratory 4172 and 2176, School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, Malaysia.
Isolation of mononucleated cells from mice peripheral blood: Mus musculus (Mice) ages between 4 to 6 weeks were used to obtain their peripheral blood. Approximately, 1 mL of peripheral blood was taken from a mouse to be isolated by density centrifugation. The blood was diluted two times with Phosphate Buffered Saline (PBS) and 2% Newborn calf serum (NBCS). The mixture then was over layered carefully onto 1.5 mL Ficoll-PaqueTM Plus and centrifuged at 400 g for 30 min at room temperature. The upper plasma layer was removed and discards without disturbing the plasma-Ficoll interface. The mononuclear cell layer at the plasma-Ficoll interface then was removed and retains without disturbing the erythrocyte/granulocyte pellet. The isolated mononuclear cells were washed four times with growth medium (alpha minimal essential medium (AMEM), 10% heat-inactivated NBCS and 2% penicillin/streptomycin) at 400 g for 15 min. After final washed, the supernatant was discarded while the pellet was dissolved with 1 mL complete growth medium and incubated in 24 well plates at 37 °C for 30 days prior to differentiation assay.
Proliferation of mononucleated cells: Mononucleated cells were cultured in complete growth medium. Cells were maintained in Alpha Minimal Essential Medium (AMEM) with 10% heat-inactivated Newborn Calf Serum (NBCS) and 2% penicillin/streptomycin. The cells were counted every day using Trypan blue exclusion method for 30 days to observe the proliferation of the cells.
Differentiation of mononucleated cells into osteoclast cells: Cells were seeded at 1.0x105 cells mL-1 in 24 well plates for osteoclast differentiation assay. The respected culture medium was then supplemented with 50 ng mL-1 RANKL and 25 ng mL-1 M-CSF to induce cells` differentiation. The cultures were maintained at 37 °C in a fully humidified atmosphere of 5% CO2 in air. For control, the same cells were cultured with complete medium without supplementation of growth factors.
Differentiation of mononucleated cells into osteoblast cells: Cells were seeded at 1.0x105 cells mL-1 in 24 well plates for osteoblast differentiation assay. The culture in complete medium was then supplemented with 50 μg mL-1 ascorbic acid and 10 mM β-glycerophosphate to induce cells` differentiation into osteoblast. The cultures were maintained at 37 °C in a humidified 5% CO2 atmosphere. For control, the same cells were cultured with complete medium without supplementation of ascorbic acid and β-glycerophosphate.
Measurement of total protein concentration: Total protein concentration was measured at the same day ALP and TRAP were assayed. The cultures were washed once with complete growth medium followed by several times with PBS. The total cellular protein concentration was determined using Bradford protein assay reagent and the absorbance was measured at 595 nm.
Tartrate resistant acid phosphatase (TRAP) assay: The cells were washed with PBS and 50 μL sodium acetate buffer (50 mM, pH 5.5) containing Triton X-100 (0.1% v/v) prior to TRAP assay. The cultures were then centrifuged at 800 g for 5 min. Supernatant was discarded and 500 μL PBS was added to each tube and stored at -20 °C until subjected to determination of TRAP activity. The cell extracts were collected and centrifuged after thawing. TRAP activity was assayed in the supernatant of the extracts.
TRAP enzyme activity in the cell extracts was assayed using p-nitrophenylphosphate (pNPP) as substrate in an incubation medium (500 μL) containing the following: 10 mM pNPP, 0.1 M Na-acetate (pH 5.8), 0.15 M KCl, 0.1% (v/v) Triton X-100, 10 mM Na-tartrate, 1 mM ascorbic acid and 0.1 mM FeCl3. The p-nitrophenol will be liberated into p-nitrophenylate after 1 h of incubation at 37 °C and by the addition of 500 μL of 0.3 M NaOH. The absorbance was immediately taken at 405 nm using a spectrophotometer. A TRAP activity was represented as specific activity. One unit of TRAP activity is represented by hydrolyzes of 1 μM pNPP per minute at 37 °C. The specific activity was determined by unit activity per total protein content (mg).
Alkaline phosphatase (ALP) assay: ALP activity in the cells was assayed after washing the cells with complete growth medium followed by PBS. Then the cells were incubated in 0.1 M NaHCO3-Na2CO3 buffer (pH 10.0) containing 0.1% (v/v) Triton X-100, 2 mM MgSO4 and 6 mM p-nitrophenolphosphate (pNPP) for 30 min at 37 °C. The reaction was stopped by adding 1 mL 1.5 M NaOH and the absorbance was measured at 405 nm. An ALP activity was represented as specific activity. One unit of ALP activity is represented by hydrolyzes of 1 μM pNPP per minute at 37 °C. The specific activity was determined by unit activity per total protein content (mg).
RT-PCR: Total RNA was isolated from treated cells using the SV Total RNA Isolation System (Promega) and immediately reverse-transcribed using AMV reverse transcriptase and Tfl DNA polymerase according to the Access RT-PCR System protocol (Promega). The specific primer sequences were determined based on ALP and TRAP sequences in NCBI database with accession number NM 007431 and NM 007388, respectively (Table 1). The annealing temperature and cycle numbers for the amplification of each gene are shown in Table 1. Then the PCR products were separated by electrophoresis on 1.0% agarose gels followed by staining with Ethidium Bromide (EtBr) before been observed and photographed under UV illumination.
Statistical analyses: Data are expressed as mean ± SE of at least three independent experiments (n = 3). Statistical
| Table 1: | Primers and conditions used for RT-PCR |
 | |
| Fw: Forward primer, Rv: Reverse primer | |
significance was computed by paired t-test using Microsoft Excel. A p<0.05 was conventionally considered as statistically significant.
RESULTS AND DISCUSSION
Isolation of mononucleated cells from mice peripheral blood: Four layers were formed after centrifugation. The upper layer is plasma platelets layer followed by monocytes, Ficoll-Paque and granulocytes/erythrocytes layers. The upper plasma layer was removed and discarded without disturbing the other layers. The monocytes layer at the plasma-Ficoll interface was removed and retained. Differences in cell density are exploited to separate granulocytes and erythrocytes from mononucleated/monocytes cells. The denser granulocytes and erythrocytes sedimented through Ficoll during centrifugation, while the monocytes remain at the plasma-Ficoll interface. Monocytes are not dense enough to penetrate into the Ficoll-Paque layer. Erythrocyte sedimentation is enhanced by aggregation due to Ficoll. Aggregation increases the rate of sedimentation of the red cells, which will forms a pellet at the bottom of the tube and well separated from monocytes. Washing and centrifugation of the harvested cells subsequently removes platelets, plasma and Ficoll-Paque contamination. The resulting cell suspension then contains highly purified and viable mononucleated cells. The respective cells will be retained and suitable for further studies.
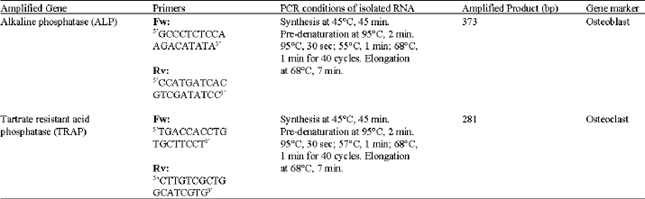
Proliferation of mononucleated cells: Two batches of mice mononucleated cells have been isolated. Each batch was proliferated in standard tissue complete culture medium. Starting from day 0 until day 30, the viability of mononucleated cells were assessed and showed 80% of cells` viability. The first and second batches of cells were showed able to proliferate in standard tissue culture medium. Figure 1 showed the viability of mononucleated cells isolated from two batches, i.e., Cell 1 (A) and Cell 2 (B). The mononucleated cells showed to increase significantly in viable cell numbers. Statistical analysis using paired t-test comparing day 0 and day 30 sets of data showed significantly (p<0.05) increased in viable cell count. The first and second batches of monocytes were proliferating in average at approximately 1491 fold after 30 days as compared to day 0 in the culture. Therefore, the cell division occurred in average of 10.5 times during 30 days in culture. This means that the primary cells need two days to divide, i.e., two days/division.
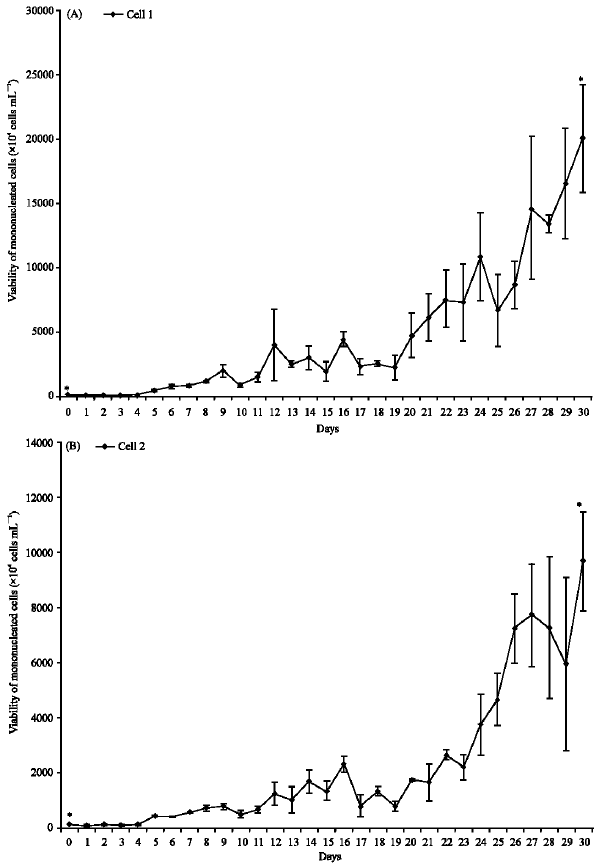
Mononucleated primary cells viability in osteoclast differentiation medium: Mononucleated cells were seeded at 1.0x105 cells mL-1 in complete medium with addition of RANKL and M-CSF for osteoclast differentiation assay after 15 days in complete proliferation medium. For control, the same cells were cultured with complete medium without supplementation of growth factors. Figure 2 showed the viability of mononucleated cells in proliferation medium vs osteoclast differentiation assay starting from day 0 to day 10. Cells from proliferation assay showed increment of 24 fold in viable cells compared to day 0, whilst, cells in osteoclast differentiation assay showed similar in numbers of viable cells throughout 10 days in the respective medium (Fig. 2). The increment in viable cells from proliferation assay is an indication that the cells are able to proliferate. Cells from osteoclast differentiation assay did not show any indication of proliferation ability since no increment showed in numbers of viable cells throughout 10 days. Statistical analysis of paired t-test between day 0 and 10 showed not significant (p>0.05). The results showed that the cells are producing survival signals during differentiation assay. Study by Faccio et al. (2003) showed that a few small multinuclear osteoclasts appeared by day 3 and became fully differentiated by
 |
| Fig. 1: | Growth curve of isolated mononucleated cells from mice`s peripheral blood. The cells` viability was determined using trypan blue staining three times on each cell. The (A) Cell 1 and (B) Cell 2, mononucleated cells were isolated from different mouse. The results showed an increment in proliferating at 1491 fold after 30 days in the culture (n = 3). * Comparison of data between day 0 and day 30 showed significantly difference using paired t-test (p<0.05) |
 |
| Fig. 2: | Viability of mononucleated cells in proliferation medium vs osteoclast differentiation assay starting from day 0 to day 10. Viable cells assessed by trypan blue exclusion is shown as mean values of viable cell numbers from three experiments with ± SEM |
day 7-10. Furthermore, studies involving primary bone marrow monocytes (Roodman, 1999) and RAW264.7 cell line (mouse macrophage cell line) (Voronov et al., 2005) showed both types of cells were fully differentiated after 7 to 10 days and 5 days in their differentiation medium, respectively. An indication that the primary mononucleated cells that produce survival signal during 10 days cultured in differentiation medium was differentiated into osteoclast cells.
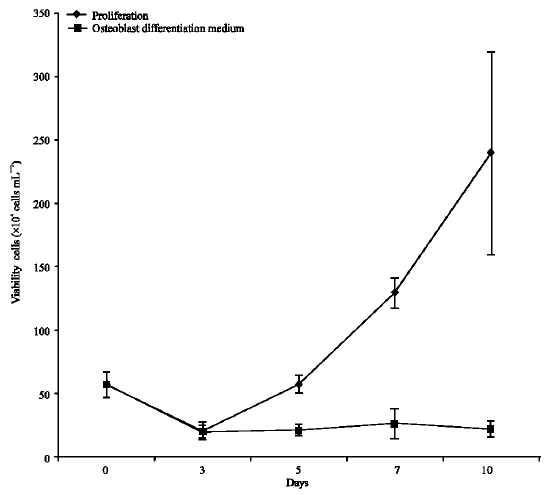
Mononucleated primary cells viability in osteoblast differentiation medium: Mononucleated cells were seeded at 1.0x105 cells mL-1 in complete medium with supplementation of ascorbic acid and β-glycerophosphate for osteoblast differentiation assay. For control, the same cells were cultured with complete medium without supplementation of growth factors. Figure 3 showed the viability of mononucleated cells in proliferation medium vs osteoblast differentiation assay starting from day 0 to 14. Cells from proliferation assay showed increment in viable cells, an indication that the cells showed the ability to proliferate. The cells cultured in proliferation medium, i.e., complete medium showed to increase 21 fold, which equivalent to 4 times cells` division as compared to cells` cultured in differentiation medium, i.e., culture medium containing acid ascorbic and β-glycerophosphate. However, cells in osteoblast differentiation assay did not showed any increment but showed similar in numbers of viable cells throughout 14 days in the respective medium. This is an indication that the mononucleated cells originated from peripheral blood are producing both proliferation and survival signals. ALP is a marker for bone formation. Studies by Wang et al. (1999) and Zhao et al. (2005) showed that the MC3T3-E1 cell line cultured in differentiation medium (presence of ascorbic acid and β-glycerophosphate) exhibits high ALP activity after 10 and 14 days in the medium, respectively. The life cycle of an osteoblast from stromal cell to cell death is approximately 30 days, during which time the cell undergoes a well-defined series of phenotypic transitions (Owen et al., 1990).
 |
| Fig. 3: | Viability of mononucleated cells in proliferation medium vs osteoblast differentiation assay starting from day 0 to day 14. Viable cells assessed by trypan blue exclusion is shown as mean values of viable cell numbers from three experiments with ± SEM |
Differentiation of osteoclast cells using tartrate resistant acid phosphatase(TRAP) activities: TRAP activities of 1x105 mononucleated cells mL-1 were measured throughout 10 days differentiation. The differentiation assay consist complete medium and supplementation of RANKL and M-CSF, whilst, the control assay consist only complete medium. Figure 4 showed the percentages of TRAP activities in differentiation assay (full gray) and control assay (white). The cells cultured in osteoclast differentiation medium were found to be increased significantly in percentage of TRAP specific activities throughout 10 days differentiation. At day 10, the respective cells produced TRAP specific activities 5.1 times higher compared to cells cultured in complete medium (control). Control cells are considered as 100% TRAP activity throughout the assay. TRAP is a metalloprotein enzyme that belongs to the acid phosphatase and known to be expressed by osteoclasts (Honig et al., 2006). Other studies also shown that higher level of TRAP activities exist when bone resorption metabolism is active (Minkin, 1982; Halleen et al., 2000; Janckila et al., 2003). In this study, TRAP activity was performed on day 3 until day 10 to detect the presence of TRAP enzyme in osteoclast differentiation medium. The mononucleated cells are found able to differentiate to osteoclast cells since the cells that cultured in respective medium has produced significant increase of TRAP activities from day 0 that act as basal activity, i.e., 100%, until day 10. Statistical analysis using paired t-test for osteoclast differentiation assay showed significantly (p<0.05) increased starting on day 3 until 10 as compared to day 0 (Fig. 4).
Differentiation of osteoblast cells using alkaline phosphatase (ALP) activities: Biochemical analysis was performed to detect osteoblast cells in the differentiation medium consist of ascorbic acid and β-glycerophosphate. Osteoblasts actively produced ALP enzyme (Kartsogiannis and Ng, 2004), therefore ALP activity is regard as biomarker for osteoblast existence. Alkaline phosphatase (ALP) was first suggested to be important in biomineralization by Robison (1923). The primary
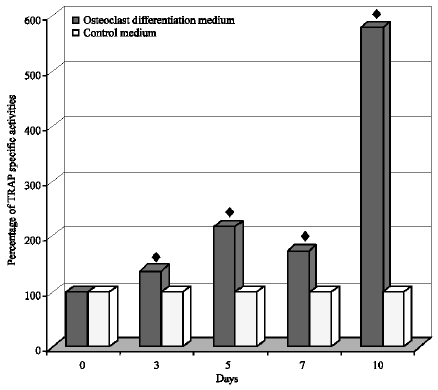 |
| Fig. 4: | TRAP activities of 1x105 mononucleated cells mL-1. Percentages of TRAP activities in differentiation assay (full gray) and control assay (white). The enzyme specific activity on day 0 and cells cultured in complete medium, i.e., control medium considered as basal activity thus 100%. ♦ Statistical analysis of paired t-test when compared to their respective control showed significant different (p<0.05) |
biological function of ALP in bone formation is associated with the calcification of the skeleton. This function is exerted either by its ability to catalyze the hydrolysis of organic phosphate esters, thereby providing inorganic phosphate, or by removing the inorganic pyrophosphate that inhibits calcification at the site of active mineralization (Kim et al., 2004). In this study, ALP activity assay was performed on day 3 until day 14 to detect the presence of ALP enzyme in osteoblast differentiation medium. ALP activities of 1x105 mononucleated cells mL-1 were measured throughout 14 days differentiation. Figure 5 showed the percentages of ALP activities in differentiation assay (full gray) and control assay (white). The ALP activities of cells cultured in osteoblast differentiation medium were found to be rapidly increased from day 3 to day 14. At day 14, ALP enzymatic activity showed 19 fold higher when compared to cells cultured in complete medium. The mononucleated cells have the ability to differentiate to osteoblast cells since the cells that cultured in osteoblast differentiation medium has produced ALP activities throughout 14 days of differentiation. Statistical analysis using paired t-test for osteoblast differentiation assay showed significantly (p<0.05) increased in producing ALP activities on day 10 and 14 as compared to day 0.
Expression of TRAP and ALP in differentiated mononucleated cells: The differentiated osteoclast cells express specific markers, such as Tartrate Resistant Acid Phosphatase (TRAP) (Perez-Amodio et al., 2005), whilst, osteoblast cells express specific markers, such as Alkaline Phosphatase (ALP) (Christenson, 1997; Chaudhry et al., 2004). We investigated the expression of selected markers in both the osteoclast and osteoblast cells. The results of the experiments are shown in Fig. 6. The TRAP was expressed only in osteoclast cells but not in the osteoblast cells. The expression of ALP was evident in the differentiated osteoblast cells. The RT-PCR analysis of the transcripts of the osteoclast and osteoblast cells showed expression of specific markers. The total RNA from these cells yielded RT-PCR amplified products of TRAP and ALP.
RT-PCR analysis was performed to determine the presence of TRAP and ALP gene in mononucleated cells cultured in osteoclast and osteoblast differentiation medium, respectively. As shown in Fig. 6, both mononucleated cells from osteoclast and osteoblast differentiation medium were positive for the presence of TRAP and ALP gene, respectively. The product showed amplification of expected size for ALP gene, i.e., ~373 bp (Lane 1) and TRAP gene, i.e., ~281 bp (Lane 3).
 |
| Fig. 5: | ALP activities of 1x105 mononucleated cells mL-1. Percentages of ALP activities in differentiation assay (full gray) and control assay (white). The enzyme specific activity on day 0 and cells cultured in complete medium, i.e., control medium considered as basal activity thus 100%. ♦ Statistical analysis of paired t-test when compared to their respective control showed significant different (p<0.05) |
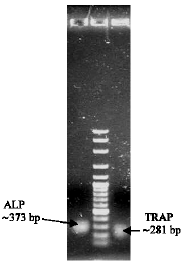 |
| Fig. 6: | Electrophoresis of 1% agarose gel showed RT-PCR products run at 85V for 60 min. Lane 1; represent RT-PCR product of RNA isolated from mononucleated cells cultured in the osteoblast differentiation assay medium for 14 days. The product showed amplification of expected size for ALP gene, i.e., ~373 bp. Lane 2 is 100 bp marker ladder whilst lane 3 is product of RT-PCR from mononucleated cells RNA cultured in osteoclast differentiation assay for 10 days. The product showed amplification of TRAP gene (~281 bp) |
CONCLUSION
Mononucleated cells were showed to produce survival signal when assayed in differentiation medium. The respective cells were also able to differentiate into osteoblast and osteoclast cells in their respective differentiation medium using biochemical and molecular markers. These were showed by increased activity of specific biochemical markers, i.e., ALP and TRAP after 10 and 14 days in their differentiation medium, respectively. Osteoblast and osteoclast cells were originated from different lineage, i.e., osteoblast from mesenchymal stromal cell lineage (Yano et al., 2005) and osteoclast was originated from monocyte-macrophage lineage precursor (Evans and Fox, 2007). Therefore, the ability of mononucleated cells to differentiate into both different cells lineages showed that this type of cells can be categorized as multipotent stem cells.
ACKNOWLEDGMENTS
The authors would like to thank the Ministry of Science, Technology and Innovation of Malaysia for the financial grants (IRPA 09-02-02-10042-EAR and Science fund 02-01-02-SF0156) during this study.
REFERENCES
- Asashima, M. and T.S. Okada, 2001. Spemann’s influence on Japanese developmental biology. Int. J. Dev. Biol., 45: 57-65.
Direct Link - Baum, C.M., I.L. Weissman, A.S. Tsukamoto, A.M. Buckle and B. Peault, 1992. Isolation of a candidate human hematopoietic stem-cell population. Proc. Natl. Acad. Sci. USA., 89: 2804-2808.
Direct Link - Bjerknes, M. and H. Cheng, 1999. Clonal analysis of mouse intestinal epithelial progenitors. Gastroenterology, 116: 7-14.
PubMed - Brazelton, T.R., F.M.V. Rossi, G.I. Keshet and H.M. Blau, 2000. From marrow to brain: Expression of neuronal phenotypes in adult mice. Science, 290: 1775-1779.
Direct Link - Buttery, L.D., S. Bourne, J.D. Xynos, H. Wood and F.J. Hughes et al., 2001. Differentiation of osteoblasts and in vitro bone formation from murine embryonic stem cells. Tissue Eng., 7: 89-99.
Direct Link - Chaudhry, G.R., D. Yao, A. Smith and A. Hussain, 2004. Osteogenic cells derived from embryonic stem cells produced bone nodules in three-dimension scaffolds. J. Biomed. Biotechnol., 2004: 203-210.
Direct Link - Christenson, R.H., 1997. Biochemical markers of bone metabolism: An overview. Clin. Biochem., 30: 573-593.
CrossRefDirect Link - Clarke, D. and J. Frisén, 2001. Differentiation potential of adult stem cells. Curr. Opin. Genet. Dev., 11: 575-580.
Direct Link - Dani, C., A.G. Smith, S. Dessolin, P. Leroy and L. Staccini et al., 1997. Differentiation of embryonic stem cells into adipocytes in vitro. J. Cell Sci., 110: 1279-1285.
Direct Link - Evans, K.E. and S.W. Fox, 2007. Interleukin-10 inhibits osteoclastogenesis by reducing NFATc1 expression and preventing its translocation to the nucleus. BMC Cell Biol., 8: 4-4.
CrossRefDirect Link - Faccio, R., S. Takeshita, A. Zallone, F.P. Ross and S.L. Teitelbaum, 2003. c-FMS and the v 3 integrin collaborate during osteoclast differentiation. J. Clin. Invest., 111: 749-758.
Direct Link - Forraz, N., R. Pettengell and C.P. McGuckin, 2004. Characterization of a lineage-negative stem-progenitor cell population optimized for Ex vivo expansion and enriched for LTC-IC. Stem Cells, 22: 100-108.
Direct Link - Fujii, H., T. Hirose, S. Oe, K. Yasuchika and H. Azuma et al., 2002. Contribution of bone marrow cells to liver regeneration after partial hepatectomy in mice. J. Hepatol., 36: 653-659.
Direct Link - Galli, R., U. Borello, A. Gritti, M.G. Minasi and C. Bjornson et al., 2000. Skeletal myogenic potential of human and mouse neural stem cells. Nat. Neurosci., 3: 986-991.
Direct Link - Gritti, A., P. Frolichsthal-Schoeller, R. Galli, E.A. Parati and L. Cova et al., 1999. Epidermal and fibroblast growth factors behave as mitogenic regulators for a single multipotent stem cell-like population from the subventricular region of the adult mouse forebrain. J. Neurosci., 19: 3287-3297.
PubMed - Halleen, J.M., S.L. Alatalo, H. Suominen, S. Cheng, A.J. Janckila and H.K. Väänänen, 2000. Tartrate-resistant acid phosphatase 5b: A novel serum marker of bone resorption. J. Bone Miner. Res., 15: 1337-1345.
Direct Link - Honig, A., L. Rieger, M. Kapp, M. Krockenberger, M. Eck, J. Dietl and U. Kämmerer, 2006. Increased tartrate-resistant acid phosphatase (TRAP) expression in malignant breast, ovarian and melanoma tissue: An investigational study. BMC Cancer, 6: 199-199.
CrossRefDirect Link - Janckila, A.J., Y.R. Nakasato, D.H. Neustadt and L.T. Yam, 2003. Disease-specific expression of tartrate-resistant acid phosphatase isoforms. J. Bone Miner. Res., 18: 1916-1919.
Direct Link - Kartsogiannis, V. and K.W. Ng, 2004. Cell lines and primary cell cultures in the study of bone cell biology. Mol. Cell Endocrinol., 228: 79-102.
Direct Link - Kim, Y.J., M.H. Lee, J.M. Wozney, J.Y. Cho and H.M. Ryoo, 2004. Bone morphogenetic protein-2-induced alkaline phosphatase expression is stimulated by Dlx5 and repressed by Msx2. J. Biol. Chem., 279: 50773-50780.
Direct Link - Kondo, T. and M. Raff, 2000. Oligodendrocyte precursor cells reprogrammed to become multipotential CNS stem cells. Science, 289: 1754-1757.
Direct Link - Kramer, J., C. Hegert, K. Guan, A.M. Wobus, P.K. Muller and J. Rohwedel, 2000. Embryonic stem cell-derived chondrogenic differentiation in vitro: Activation by BMP-2 and BMP-4. Mech. Dev., 92: 193-205.
Direct Link - Krause, D.S., N.D. Theise, M.I. Collector, O. Henegariu and S. Hwang et al., 2001. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell, 105: 369-377.
Direct Link - Lagasse, E., H. Connors, M. Al-Dhalimy, M. Reitsma and M. Dohse et al., 2000. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat. Med., 6: 1229-1234.
Direct Link - Malouf, N.N., W.B. Coleman, J.W. Grisham, R.A. Lininger and V.J. Madden et al., 2001. Adult-derived stem cells from the liver become myocytes in the heart in vivo. Am. J. Pathol., 158: 1929-1935.
Direct Link - Martin, G.R., 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA., 78: 7634-7638.
Direct Link - Minkin, C., 1982. Bone acid phosphatase: Tartrate-resistant acid phosphatase as a marker of osteoclast function. Calcif. Tissue Int., 34: 285-290.
CrossRefDirect Link - Morrison, S.J., 2001. Stem cell potential: Can anything make anything? Curr. Biol., 11: R7-R9.
Direct Link - Orlic, D., J. Kajstura, S. Chimenti, I. Jakoniuk and S.M. Anderson et al., 2001. Bone marrow cells regenerate infracted myocardium. Nature, 410: 701-705.
Direct Link - Owen, T.A., M. Aronow, V. Shalhoub, L.M. Barone and L. Wilming et al., 1990. Progressive development of the rat osteoblast phenotype in vitro: Reciprocal relationships in expression of genes associated with osteoblast proliferation and differentiation during formation of the bone extracellular matrix. J. Cell Physiol., 143: 420-430.
CrossRef - Pearce, D.J., D. Taussig, C. Simpson, K. Allen, A.Z. Rohatiner, T.A. Lister and D. Bonnet, 2005. Characterization of cells with a high aldehyde dehydogenase activity from cord blood and acute myeloid leukemia Samples. Stem Cells, 23: 752-760.
Direct Link - Perez-Amodio, S., I.M.C. Vogels, T. Schoenmaker, D.C. Jansen and S.L. Alatalo et al., 2005. Endogenous expression and endocytosis of tartrate-resistant acid phosphatase (TRACP) by osteoblast-like cells. Bone, 36: 1065-1077.
Direct Link - Robison, R., 1923. The possible significance of hexosephosphoric esters in ossification. Biochem. J., 17: 286-293.
Direct Link - Sadiq, T.S. and D.A. Gerber, 2004. Stem cells in modern medicine: Reality or myth? J. Surg. Res., 122: 280-291.
Direct Link - Shahrul-Hisham, Z.A., M.A.W. Rohaya, I. Ismanizan, M. Nor-Muhammad and Z.A. Zaidah, 2005. Stem cells, cytokines and their receptors. Asia Pacific J. Mol. Biol. Biotechnol., 13: 1-13.
Direct Link - Shanthly, N., M.R. Aruva, K. Zhang, B. Mathew and M.L. Thakur, 2006. Stem cells: A regenerative pharmaceutical. Q. J. Nucl. Med. Mol. Imag., 50: 205-216.
Direct Link - Spangrude, G.J., L. Smith, N. Uchida, K. Ikuta, S. Heimfeld, J. Friedman and I.L. Weissman, 1991. Mouse hematopoietic stem cells. Blood, 78: 1395-1402.
Direct Link - Theise, N.D., S. Badve, R. Saxena, O. Henegariu and S. Sell et al., 2000. Derivation of hepatocytes from bone marrow cells in mice after radiation-induced myeloablation. Hepatology, 31: 235-240.
Direct Link - Theise, N.D., M. Nimmakayalu, R. Gardner, P.B. Illei and G. Morgan et al., 2000. Liver from bone marrow in humans. Hepatology, 32: 11-16.
CrossRefDirect Link - Voronov, I., J.N.M. Heersche, R.F. Casper, H.C. Tenenbaum and M.F. Manolson, 2005. Inhibition of osteoclast differentiation by polycyclic aryl hydrocarbons is dependent on cell density and RANKL concentration. Biochem. Pharmacol., 70: 300-307.
Direct Link - Wakitani, S., T. Saito and A.I. Caplan, 1995. Myogenic cells derived from rat bone marrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve, 18: 1417-1426.
PubMed - Wang, D., K. Christensen, K. Chawla, G. Xiao, P.H. Krebsbach and R.T. Franceschi, 1999. Isolation and characterization of MC3T3-E1 preosteoblast subclones with distinct in vitro and in vivo differentiation/mineralization potential. J. Bone Miner. Res., 14: 893-903.
CrossRefDirect Link - Watt, F.M. and B.L. Hogan, 2000. Out of Eden: Stem cells and their niches. Sci., 287: 1427-1430.
Direct Link - Weissman, I.L., 2000. Translating stem and progenitor cell biology to the clinic: Barriers and opportunities. Science, 287: 1442-1446.
Direct Link - Yamashita, J., H. Itoh, M. Hirashima, M. Ogawa and S. Nishikawa et al., 2000. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature, 408: 92-96.
CrossRefPubMedDirect Link - Yang, L., S. Li, H. Hatch, K. Ahrens, J.G. Cornelius, B.E. Petersen and A.B. Peck, 2002. In vitro trans-differentiation of adult hepatic stem cells into pancreatic endocrine hormone-producing cells. Proc. Natl. Acad. Sci. USA., 99: 8078-8083.
Direct Link - Yano, S., R. Mentaverri, D. Kanuparthi, S. Bandyopadhyay, A. Rivera, E.M. Brown and N. Chattopadhyay, 2005. Functional expression of-chemokine receptors in osteoblasts: Role of regulated upon activation, normal T cell expressed and secreted (RANTES) in osteoblasts and regulation of its secretion by osteoblasts and osteoclasts. Endocrinology, 146: 2324-2335.
Direct Link - Zhao, Y., H. Guan, S.F. Liu, R.C. Wu and Z. Wang, 2005. Overexpression of QM induces cell differentiation and mineralization in MC3T3-E1. Biol. Pharm. Bull., 28: 1371-1376.
Direct Link