
E. Farshadfar
College of Agriculture, Razi University, Kermanshah, Iran
M. Aghaie Sarbarzeh
Seed and Plant Institute, Karage, Iran
M. Sharifi
College of Agriculture, Razi University, Kermanshah, Iran
A. Yaghotipoor
College of Agriculture, Razi University, Kermanshah, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 2 | Page No.: 461-465







ABSTRACT
In order to estimate additive and dominance components of genetic variance and detection of non-allelic interaction for the salt tolerance criteria in barley, 7 generations (P1, P2, F1, F2, F3, BC1, BC2) derived from the cross Wiesel burger/AborxLokus/Bda and the check were used in a split plot design with two replications in hydroponic culture. Significant differences were found between both salinity levels and genotypes for all the characters investigated. Mean generation analysis indicated the involvement of additive, dominance and epistatic type of gene action in the inheritance of leaf weight (additive, dominance and epistasis), biomass (additive and epistasis), K+, Na+ and K+/Na+ (dominance and epistasis). Heritability estimate was low for K+ and Na+, moderate for shoot length, leaf weight, biomass and K+/Na+ and high for root length. Over dominance type of gene action was found for shoot length, biomass, K+, Na+ and K+/Na+, while partial dominance for root length and leaf weight.
PDF Abstract XML References Citation
How to cite this article
E. Farshadfar, M. Aghaie Sarbarzeh, M. Sharifi and A. Yaghotipoor, 2008. Assessment of Salt Tolerance Inheritance in Barley via Generation Mean Analysis. Journal of Biological Sciences, 8: 461-465.
DOI: 10.3923/jbs.2008.461.465
URL: https://scialert.net/abstract/?doi=jbs.2008.461.465
DOI: 10.3923/jbs.2008.461.465
URL: https://scialert.net/abstract/?doi=jbs.2008.461.465
INTRODUCTION
Salinity is considered as a major limiting factor for crop plants especially in arid and semi-arid regions (Forster et al., 1990; Munns et al., 2002). Developments of salt tolerant varieties which can be adapted to a wide range of environmental stresses is the ultimate goal of plant breeders in barley improvement programs (Sabaghpour et al., 2003). Improving salt tolerance is therefore a major objective in plant breeding programs for arid and semi-arid regions (Farshadfar et al., 2007).
To formulate an efficient breeding program for developing salt-tolerant varieties, it is essential to understand the mode of inheritance, the magnitude of gene effects and their mode of action (Farshadfar et al., 2001; Sharma, 1998). Many workers developed genetic models for the estimation of different genetic effects (Gamil and Saheal, 1986; Kearsey and Pooni, 1996). However, the majority of these genetic models are basically additive-dominance models or simply additive models.
The epistatic or non-allelic interactions are largely ignored so as to have a simplified interpretation of genetic variation. But, it has now been established that such inter-allelic interactions are of frequent occurrence in the control of trait-expression for continuous variation. Thus, inferences drawn from additive models are likely to be based to an unknown extent. That is why, Jinks et al. (1969) suggested: it is no longer possible to Justify the use of a biometrical genetical analysis which does not have a built in test for epistasis. To be on safe side, it is therefore, rather essential to test the presence or absence of non-allelic interaction i.e., to carry out test of additivity or epistasis (Sharma, 1998; Kearsey and Pooni, 1996; Singh and Chaudhary, 1995; Farshadfar, 1998).
More than one procedure is available to test the deviation from additive models, i.e. to detect epistatic effects. These are: Wr-Vr tests of additivity (Hayman, 1954), the triple test cross test (Kearsey and Jinks, 1968), test of epistasis (Jinks et al., 1969), computation of interaction per se (Jinks and Jones, 1958) and scaling test (Mather, 1949; Hayman, 1954; Jinks and Jones, 1958).
We know that phenotypic mean is consummated by additive (a), dominance (d) and interaction effects (i) of genes in point.
The interaction effect is again of two kinds: (i) complementary (aa) and (ii) duplicate (ad and dd) at digenetic level.
The analysis of generation mean provides the opportunity first to detect the presence or absence of epistasis (by scaling test) and when present, it measures them appropriately. It also determines the components of heterosis in terms of gene-effects and some other statistics. Such as potence ratio, levels of dominance, number of effective factors, etc. (Kearsey and Pooni, 1996; Singh and Narayanan, 1993; Farshadfar, 1998; Farshadfar et al., 2001, 2007).
The objectives of the present investigation were to genetic analysis of salt tolerance criteria in barley, to detection of non-allelic interaction for the characters investigated and to the estimation of additive and dominance components of genetic variance for the traits not influenced by epistasis.
MATERIALS AND METHODS
The experimental materials consisted of 7 generations (P1, P2, F1, F2, F3, BC1 and BC2) derived from crosses between Wiesel burger/Abor cultivar (P2 = salt tolerant) and Lokus/Bda (P1 = salt sensitive) in the experimental farm of Dryland Agricultural Research Institute, Sararood, Kermanshah, Iran in 2003 and 2004. The parents (P1 and P2), the first (F1) and second (F2) generation hybrids and the first (P1xF1 = BC1) and second (P2xF1 = BC2) backcrosses together with check (Sararood = adaptable to the region) were grown in the green house of college of Agriculture, Razi University, Kermanshah, Iran. A split plot design based on completely randomized design (CRD) with two replications was carried out using hydroponic culture. Salinity was the main plot with three levels (0, 75 and 150 μmol of NaCl L-1 and genotypes were considered as sub-plot with 8 levels (parents, subsequent generations and control).
From each genotype 70 seeds were selected, surface sterilized with 5% calcium hypochlorite for 3 min, rinsed in distilled water and placed on sterile blotting paper in petriplates. They were germinated at 27°C and then transferred into growth chamber. At two leaves stage, 30 seedlings with roots of approximately 5 cm length from each genotype were selected and transferred into the tanks.
Hogland solution was the basic nutrient stock solution used in the Hydroponic culture system (Gorham, 1990). Saline solution was made for each treatment using NaCl and CaCl2 salts with the pH of 5.45 to 5.55. The plantlets were hanged in the solution in such a way that roots were immersed in the aerated solution. The tanks were kept in growth chamber at 16-24°C, 300 μmol m-3 sec-1 light intensity, 18/6 h (light/darkness) and 45-50% relative humidity (Forster et al., 1990).
Plants were grown in tanks for 2-3 weeks (2-3 leaf stage of seedling), the saline solution was gradually added to the container in such a way that final concentration was obtained after 4 days.
Five plants from fully expanded leaf (the 5th leaf) of each genotype were selected and the traits root length (RL), shoot length (SL), 5th leaf weight (LW) and biomass (Bio) were measured. The fully expanded leaf blade samples from each genotype were collected, rinsed in distilled water for 5 sec and dried at 70°C. The samples were ground with a mortar and pestle, dissolved in 10% acetic acid and 0.1% nitric acid and kept at room temperature for 24 h for complete digestion. Each sample was diluted 1:6 with acid solution and analyzed by flame photometer and the concentration of Na+, K+ and the ratio of K+/ Na+ were determined.
Statistical analysis: Analysis of variance, mean comparison and correlation analysis was done using SPSS and MSTAT-C software.
Generation mean analysis was performed using the Mather and Jinks method (1982). In this method the mean of each character is indicated as follows:
Y = m+ α [d] + β [h] + α2 [i] +2α β [j] + β2 [l]
where, Y = the mean of one generation, m = the mean of all generation, [d] = the sum of additive effects, [h] = the sum of dominance effects, [i] = the sum of additivexadditive interaction (complementary), [l] = the sum of dominancexdominance interaction (duplicate), [j] = sum of additivexdominance and α, β, 2α β and β2 are the coefficients of genetic parameters. The genetic parameters (m, [d], [h], [i], [j], [l]) were tested for significance using a t-test. The adequacy of the additive dominance model was determined by χ2-test. Broadsense and narrowsense heritability were estimated by Warnner (1952) and Allard (1960) formulas:
Hb2 = [VF2-(VP1 + VP2 + VF1)/ 3]/ VF2
Hn2 = [2VF2-(VBC1 + VBC2)]/ VF2
The components of F2 variance were obtained by the following formula (Farshadfar, 1998):
D = 4VF2 -2 (VBC1 + VBC2)
H = 2VF2-2D-2E
E = 1/3 (VP1 + VP2 + VF1)
F = VBC1 + VBC2
where, V means variance.
RESULTS AND DISCUSSION
The results of analysis of variance (Table 1) revealed significant differences for salinity levels and generations for all the characters investigated indicating the presence of genetic variability and possibility of selection for salt tolerance (Baghizadeh et al., 2003).
| Table 1: | Analysis of variance for various characters investigated |
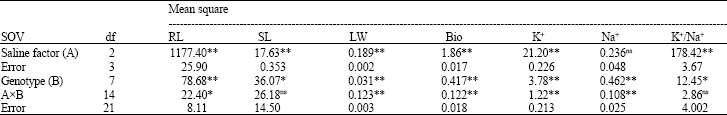 | |
| *,**, ns: Significant at 0.05, 0.01 level of probability and non-significant, respectively | |
| Table 2: | Mean comparison between sub-main plots (genotypes) for various characters investigated |
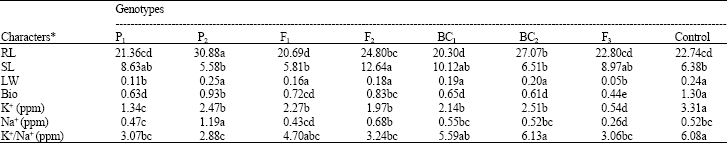 | |
| *: Common letter(s) mean no significant difference | |
| Table 3: | Correlation coefficients between the characters studied |
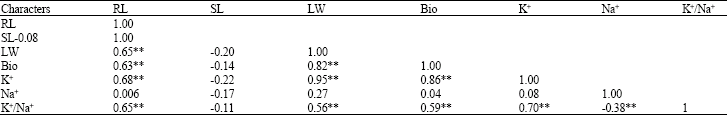 | |
| **: Significant at 1% level of probability | |
| Table 4: | Estimates of genetic components of the mean for the characters studied |
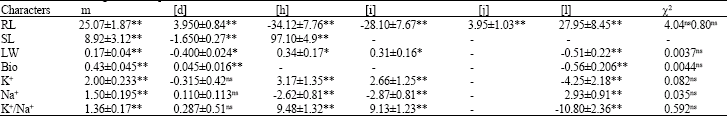 | |
| *,**, ns: Significant at 5%, 1% level of probability and non-significant, respectively | |
The genotypexsalt tolerance interaction was also significant, except for SL and K+/ Na+ displaying their similar response and different responses of other traits. Mean comparison (Table 2) between the genotypes showed that P1 and P2 have significant differences for most of the traits. The F1 hybrid, BC1 and BC2 exhibited the highest heterosis over the best parent, although it was not significant for F1 and BC2 and significant for BC1, for the criteria of salt tolerance (K+/Na+) (Schachtman and Munns, 1992; Deal et al., 1999). Most of the K+ uptake belonged to the control (Sararood). Most of the Na+ uptake was attributed to P2 which is salt tolerant.
High significant correlation coefficient (Table 3) was found between RL, LW and Bio with K+ and K+/Na+ indicating that these traits enhance salt tolerance in barley through correlated response. Omielan et al.(1991) showed that K+/ Na+ ratio was highly correlated with biomass, RL and LW (Farshadfar et al., 2007).
Generation mean analysis: The joint scaling test (Mather and Jinks, 1982; Farshadfar, 1998) was employed to estimate the mean (m), additive effect (d), dominance effect (h), additivexadditive (i), additivexdominance (j) and dominancexdominance (l) values. The best dominance-additive model was selected using non-significant Chi-square test (χ2) and the lower standard level (Table 4).
The dominance-additive model was adequate and fitted for all the characters investigated. All the genetic components of the mean were significant for root length indicating that besides additive and dominance gene effects, epistasis is also involved in the inheritance of RL.
| Table 5: | Genetic parameters and components of variation for various characters investigated |
 | |
The contribution of dominance effect (h) and epistasis (I) is greater than additive effect. As the sign of h and l is opposite, hence duplicate epistasis is involved (Farshadfar et al., 2001; Zahravi, 1999). Significancy of (j) revealed that selection through selfing is not effective for improvement of root length (Farshadfar et al., 2001; Sharifi, 2005).
For shoot length (d) and (h) were significant but inter-allelic interaction was not significant, therefore the genetic of SL is under the control of dominance and additive type of gene action, but the contribution of dominance effect is greater (Honarnejad and Tarang, 2000). The components [d], [h], [i] and [l] were significant for leaf weight exhibited the involvement of additive, dominance and epistatic effect in the control of leaf weight. The opposite sign of [h] and [l] displayed duplicate epistasis. As [j] was not significant and the contribution of [d] was greater than [h], hence selection under selfing can be effective for the breeding of leaf weight (Farshadfar et al., 2001; Sharifi, 2005).
For the biomass only [d] and [l] were significant, therefore additive type of gene action and epistasis are important in the inheritance of this trait and selection through selfing will be effective (Khalifa, 1982).
The components [h], [i] and [l] were significant for the characters K+, Na+ and K+/ Na+ indicating the role of dominance and epistasis in the control of these salt tolerance criteria. As the sing of [h] and [l] is opposite, hence duplicate type of epistasis is involved. This complementary interaction increases the variation between the generations and in the segregating population (Farshadfar et al., 2007).
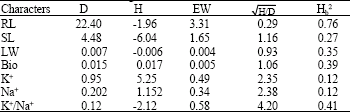
Heritability and genetic components of variation are presented in Table 5. The variation observed between the genotypes for the characters investigated exhibited that selection may be effective for the improvement of salt tolerance, however, the selection efficiency is related to the magnitude of heritability (Farshadfar et al., 2001).
Low broadsense heritability (less than 0.2) was shown for K+ and Na+, but SL, LW, Bio and K+/ Na+ displayed moderate (0.2-0.5) and root length revealed high (greater than 0.5) heritability (Stanfield, 2002). The low Hb for K+ and Na+ suggested that environmental effects constitute a major portion of the total phenotypic variation for these characters (Khaksar, 2006).
D, H and EW (Table 5) are additive, dominance and environmental components, respectively. The ratio![]() is a suitable estimator of dominance (Kearsey and Pooni, 1996; Farshadfar, 1998; Khaksar, 2006). The ratio of
is a suitable estimator of dominance (Kearsey and Pooni, 1996; Farshadfar, 1998; Khaksar, 2006). The ratio of![]() showed over dominance for SL, Bio K+, Na+ and K+/ Na+, while, partial dominance for RL and LW.
showed over dominance for SL, Bio K+, Na+ and K+/ Na+, while, partial dominance for RL and LW.
REFERENCES
- Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Deal, K.R., S. Goyal and J. Dvorak, 1999. Arm location of Lophopyrum elongatum genes affecting K+/Na+ selectivity under salt stress. Euphytica, 108: 193-198.
CrossRefDirect Link - Farshadfar, E., M. Ghandha, M. Zahravi and J. Sutka, 2001. Generation mean analysis of drought tolerance in wheat (Triticum aestivum L.). Acta Agron. Hung, 49: 59-66.
Direct Link - Forster, B.P., M.S. Phillips, T.E. Miller, E. Baird and W. Powell, 1990. Chromosome location of genes controlling tolerance to salt (NaCl) and vigor in Hordeum vulgare and Hordeum chilense. Heredity, 65: 99-107.
CrossRef - Gorham, J., 1990. Salt tolerance in the Triticeae: K/Na discrimination in synthetic hexaploid wheats. J. Exp. Bot., 41: 623-627.
CrossRefDirect Link - Jinks, J.L. and R.M. Jones, 1958. Estimation components of heterosis. Genetics, 43: 223-234.
Direct Link - Kearsey, M.J. and J.L. Jinks, 1968. A general method of detecting additive, dominance and epistatic variation for metrical traits I. Theory. I. Heredity, 23: 403-409.
CrossRefDirect Link - Kearsey, M.J. and J.L. Jinks, 1968. A general method of detecting, additive, dominance and epistatic variation for metric traits. I. Theor. Heredity, 23: 403-409.
PubMed - Kearsey, M.J. and H.S. Pooni, 1996. The Genetical Analysis of Quantitative Traits. 1st Edn., Chapman and Hall, London.
Direct Link - Munns, R., S. Husain, A.R. Rivelli, T. Condon and A.G. Lindsay et al., 2002. Avenues for increasing salt tolerance of crops and the role of physiologically based selection traits. Plant Soil, 247: 93-105.
CrossRefDirect Link - Omielan, J.A., E. Epstein and J. Dvorak, 1991. Salt tolerance and ionic relations of wheat as affected by individual chromosomes of salt-tolerant Lophopyrum elongatum. Genome, 34: 961-974.
CrossRefDirect Link - Schachtman, D.P. and R. Munns, 1992. Sodium accumulation in leaves of Triticum species that differ in salt tolerance. Aust. J. Plant Physiol., 19: 331-340.
Direct Link - Singh, R.K. and B.D. Choudhary, 1995. Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi, Pages: 318.
Direct Link