
Jeny Rachid Cursino-Santos
Departamento de Genetica, Faculdade de Medicina de Ribeir�o Preto, Universidade de S�o Paulo, SP, Brazil, Av. Bandeirantes, 3900, Ribeir�o Preto, SP, 14049-900, Brazil
Luciana Meneguim
Instituto Agron�mico do Paran�, Londrina, PR, Brazil, Rodovia Celso Garcia Cid, km 375, Tr�s Marcos, Londrina, PR, 86047-902, Brazil
Rui Pereira Leite Jr.
Instituto Agron�mico do Paran�, Londrina, PR, Brazil, Rodovia Celso Garcia Cid, km 375, Tr�s Marcos, Londrina, PR, 86047-902, Brazil
Nilce Maria Martinez-Rossi
Departamento de Genetica, Faculdade de Medicina de Ribeir�o Preto, Universidade de S�o Paulo, SP, Brazil, Av. Bandeirantes, 3900, Ribeir�o Preto, SP, 14049-900, Brazil
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 2 | Page No.: 349-355







ABSTRACT
The deduced amino acid sequence of drrA gene (XAC3759) from Xanthomonas axonopodis pv. citri (Xac) is similar to a variety of earlier described functionally uncharacterized ABC (ATP-Binding Cassette) transporters. In order to investigate the biological process in which this gene is involved, we constructed a drrA deletion mutant strain and tested its susceptibility to drugs with different mechanisms of action and to oxidative stress and also its capability to infect Citrus sinensis. The results showed that Xac responses to seven drugs (tetracycline, penicillin, ampicillin, chloramphenicol, acriflavine, ethidium bromide and doxorubicin chloridrate) and to host infection are independent upon drrA expression. However, Northern Blot analyses revealed that drrA gene is induced as short response to H2O2 exposure, only at the early phase of bacterial growth. Furthermore, the drrA disruption rendered the mutant more sensitive to oxidative stress than the control strain, suggesting that this transporter plays an important role in modulating oxidative stress susceptibility in Xac. Also, the drrA gene is co-transcripted with two hypothetical ORFs (Open Read Frame) as an operon cluster (named here drrABC) when Xac wild type strain was submitted to oxidative stress. The sequence analysis and topology prediction of the deduced proteins from these two neighboring hypothetical ORFs (XAC3758 and XAC3757) show that they have 6 transmembrane segments which are characteristic of permeases, supporting the structure of ABC transporter systems with an ABC subunit and two permeases.
PDF Abstract XML References Citation
How to cite this article
Jeny Rachid Cursino-Santos, Luciana Meneguim, Rui Pereira Leite Jr. and Nilce Maria Martinez-Rossi, 2008. Disruption of drrA Gene Affects the Xanthomonas axonopodis pv. citri Response to Oxidative Stress. Journal of Biological Sciences, 8: 349-355.
DOI: 10.3923/jbs.2008.349.355
URL: https://scialert.net/abstract/?doi=jbs.2008.349.355
DOI: 10.3923/jbs.2008.349.355
URL: https://scialert.net/abstract/?doi=jbs.2008.349.355
INTRODUCTION
Xanthomonas axonopodis pv. citri (Xac) is a soil Gram-negative bacterium that causes citrus bacterial canker, a serious disease of most of citrus species and cultivars in many citrus-producing areas worldwide. This disease leads to the defoliation and premature abscission of affected fruit causing considerable losses for growers and for the economy of citrus-producing countries (Verniere et al., 2003).
Xac genome was sequenced and 4313 ORFs (Open Read Frame) were identified in 5,175,554 bp (Da Silva et al., 2002). The comparative genomic showed that 62.8% of these ORFs have an assigned function inferred by computational analyses, which provides only a general prediction of biochemical function through the identification of the family-specific sequence pattern (family signature). ATP-binding cassette systems (ABC transporters), for instance, are easily identified in the annotation process of sequenced genomes through their three conserved motifs known as the Walker A and B sites and the linker peptide with the signature motif LSSGQ, facilitating the protein clustering even without biological function knowledge (Dassa and Bouige, 2001; Ren and Paulsen, 2005; Biemans-Oldehinkel et al., 2006). However, in order to establish the biological role genes, direct experimentations are necessary.
The gene drrA of Xac (drug resistance gene) (XAC3759, GenBank accession No. AAM38602) was previously identified by the Xac complete genome sequencing (Da Silva et al., 2002) and was characterized as an ATP-binding protein from an ABC transporter, supposedly involved with drug resistance by drug efflux pump. However, an updated sequence similarity analysis using BLASTx program (Basic Local Alignment Search Tool) reveals a higher similarity of XAC3759 nucleotide sequence with functionally uncharacterized proteins from phytopathogenic and environmental Gram-negative bacteria than with drug efflux system (97% identity with X. campestris pv. vesicatoria GenBank accession No. CAJ25609; 92% identity with X. oryzae pv. oryzae GenBank accession No. BAE67328; 86% identity with X. campestris pv. campestris GenBank accession No. AAM42975 and 65% identity with Sphingomonas sp. GenBank accession No. EAT09413).
As it has been shown that ABC transporters play an important role in bacterial lifestyle, virulence and survival besides their involvement on drug resistance mechanism (Burse et al., 2004; Brencic and Winans, 2005; Piddock, 2006), the aim of this study was to investigate the biological process in which drrA gene is involved in Xac. For so, we constructed a drrA deletion mutant strain (ΔdrrAXac) to determine whether the inactivation of drrA gene would affect Xac susceptibility to drugs and to oxidative stress as a simulation of the induced stress conditions at the pathogen-plant interaction when reactive oxygen species (ROS: hydroxyl radical, superoxide anion and hydrogen peroxide) are produced by the host and readily reacted with various pathogen cellular components inhibiting microbial survival (Lamb and Dixon, 1997). Furthermore, we evaluated Xac capability to infect and develop disease in citrus.
MATERIALS AND METHODS
Bacterial strains, plasmids and growth conditions: The Xac306 strain, isolated from citrus canker lesions of sweet orange leaves (Citrus sinensis), was obtained from the culture collection of Instituto Agronômico do Paraná (IAPAR) (Londrina, PR, Brazil) and its genome has been completely sequenced (Da Silva et al., 2002). Xac306 and derived strains were routinely grown in nutrient broth (NYGB) medium (Daniels et al., 1984) or NYGB agar plates for 24-48 h at 28°C. Escherichia coli, DH5α, used during the construction of the disruption cartridge, was grown on Luria-Bertani (LB) agar medium or in LB liquid medium for 12-24 h at 37°C. E. coli DH5α and Xac competent cells were prepared for the heat shock (Sambrook et al., 1989) and electroporation (White and Gonzales, 1991) transformation methods, respectively. Kanamycin and ampicillin, both 50 μg mL-1, were used to maintain selection for resistance markers. The vectors were extracted using the QIAGEN Plasmid Mid Kit (QIAGEN).
Construction of a ΔdrrAXac mutant: Standard techniques for molecular cloning were used as previously described by Sambrook et al. (1989). Genomic DNA from Xac was isolated by protein K lysis method (Ausubel et al., 1987). A ΔdrrAXac mutant was constructed by the disruption of the drrA gene from Xac with a kanamycin resistance cartridge (Kanr). The pdrrA vector was constructed into pGEM®-T Easy vector by cloning a 1275 kb PCR amplification product of Xac genomic DNA using the primers Up1: 5' -GGAGACAGGGAAAGCATTGG-3' and Dn1: 5' -GGGCGATAGATGAAATGG-3', which annealed at 295 nt upstream and 255 nt downstream of drrA start and stop codon, respectively. The pdrrA was digested with BalI and StuI to remove 537 bp from the cloned drrA in which two conserved motifs of the ABC transporter were localized (Walker A and Signature ABC). A 1283 bp EcoRI fragment encoding a kanamycin resistance gene from the pUC4K vector (GE Healthcare) replaced the removed fragment to generate the plasmid pGEM-ΔdrrA. The construction was confirmed by DNA automatic sequencing (ABI Prism®377 DNA Sequencer System). The disruption cartridge was recovered from pGEM-ΔdrrA on a 2021 bp fragment by NotI enzymatic restriction. Subsequently, it was inserted into a single EcoRV site present within the polylinker of pKX33 (Baldini et al., 1999) generating the plasmid pKX33-ΔdrrA, which was used to electrotransform the Xac306 strain as previously described (Ferreira et al., 1995). The Xac306 strain was also electrotransformed with plasmid pKX33 and the resultant strain (Xac-KX33) used as a control. Klenow E. coli DNA polymerase I (USB Corporation) was used to fill protruding ends, when necessary.
The confirmation of drrA deletion by homologous recombination was done by the identification of a 2622 bp PCR amplification product of ΔdrrAXac genomic DNA by using primer Up2: 5' -AACACCTCCAGCGACAGC-3' and primer Dn2: 5' -TAACCCAGGAAGACCAGC-3', which annealed 603 nt upstream and 449 nt downstream of drrA start and stop codon, respectively. This region corresponds to the Xac chromosomal sequence flanking the targeted sequence cloned in the pGEM-T easy vector that was absent in the vector and in the disruption cartridge. The deletion in the ΔdrrAXac mutant was also confirmed by probing a DNA gel blot with both BalI-StuI deleted fragment and disruption cartridge gene Kanr by Southern Blotting method (Sambrook et al., 1989).
Antibiotic and H2O2 susceptibility assay: The Xac strains were spread separately over NYGB agar broth. Sterile 3 MM filter paper disks (0.65 cm2) were impregnated with 10 μL of drug solution and placed on the surface of the inoculated NYGB agar plates. The concentrations of drug solution were: tetracycline, 10 μg μL-1 penicillin, 50 μg μL-1; ampicillin, 50 μg μL-1; chloramphenicol, 100 μg μL-1; acriflavine, 20 μg μL-1; ethidium bromide, 10 μg μL-1 and doxorubicin chloridrate, 4 μg μL-1 (antibiotic from the daunorubicin anthracycline family) (Adriblastina RD®-Pharmacia and Upjohn). Following 48 h incubation at 28°C, the diameter of the bacterial growth inhibition zones surrounding the filter disc was measured. The relative susceptibility of different strains to antibiotics was evaluated based on zone of inhibition around filter discs.
The bacterial growth was also monitored spectrophotometrically after adding H2O2 at two concentrations (1 and 2 mM H2O2) and at two bacterial growth phases: early-phase (OD640 nm = 0.3-0.5 before log phase of bacterial growth) and exponential-phase (OD640 nm >0.5 at exponential log phase culture). This assay was performed with 2 mL of each strain overnight cultures inoculated separately into 30 mL of NYGB broth at 28°C. Xac cultures were grown with shaking for 2-3 h until they reached the desired growth phase before H2O2 addition. Samples (2 mL) were then removed at 0, 2, 6, 8, 10 h intervals after H2O2 addition and optic density was measured.
Determination of drrA gene expression conditions: Northern Blot analyses (Sambrook et al., 1989) were performed with Xac306 cultures after drug and H2O2 treatments at early- and exponential-phase cells. In both cases, the cultures were maintained under shaking and aliquots were removed after 5, 10 and 20 min of exposure to drug treatment and 5, 10, 20, 30 and 60 min of H2O2 treatment. The drug concentrations used were: tetracycline, 5 μg mL-1; penicillin G, 100, 200, 350 μg mL-1; ampicillin, 100 μg mL-1; acriflavine, 0.5 μg mL-1; doxorubicin chloridrate, 0.5, 2.5, 5, 10 μg mL-1. The H2O2 concentrations were 1.5, 2, 3 and 6 mM. The total RNA was isolated with a RNA isolation system (Promega). The BalI-StuI fragment obtained during the cartridge construction was used as probe.
In vivo infection assay: Analyses of host symptoms and bacterial population dynamics in plant tissue were carried out as described previously (Mehta and Leite, 2001) and conducted at IAPAR (Londrina, Paraná State) in 2005. Bacterial suspensions at 105 cfu mL-1 were used to infiltrate young leaves of C. sinensis, a susceptible host. The plants had been pruned to induce new leaf growth and were maintained under greenhouse conditions. Each bacterial strain was inoculated in leaves of different plants by hypodermic syringe carrying out the infiltration. In addition, other plants were also assayed with the three bacterial strains inoculated in different leaves. Approximately 10 leaves per plant were inoculated with bacterial suspensions of each Xac strains. The bacterial growth was determined by sampling 3 foliar disks of 0.63 cm2, which were macerated in 1.5 mL of distilled water. Samplings were made at 0, 1, 2, 4, 8 and 16 days after the inoculation and each suspension obtained was diluted to appropriate concentrations and 100 μL were plated onto the culture medium. The plates were then incubated for 2-3 days.
Sequence analysis of the drrAXac locus and neighborhood: Nucleotide and protein sequence analyses were performed using the InterPro database (Mulder et al., 2003) and Conserved Domain Architecture Retrieval Tool (CDART; of NCBI website) to identify domains and functional sites in the deduced proteins. HMMTOP, a biotool from the Transporter Classification Data Base (TCDB version 2.0.1) (Ren et al., 2004) was used for the transmembrane segment prediction of proteins.
RESULTS AND DISCUSSION
The drrA mutant strain was constructed to perform mutant and wild comparison trials (Fig. 1A). The obtained PCR amplification patterns with primers Dn2 and Up2 showed that the genomic DNA of mutant strain was 740 pb longer than wild type (Fig. 1B) assuring the drrA disruption by deletion replacement with cartridge gene Kanr integration after homologous recombination. The Southern blotting analysis of total DNA of Xac transformed cells confirmed the mutant strain isolation. BalI-StuI deleted fragment and cartridge gene Kanr were used as probe (data not shown).
The effect of drrA gene inactivation on the capacity of Xac to response to different stress conditions and to infect citrus developing disease was evaluated through the comparison among the three bacterial strains (ΔdrrAXac, Xac306 wild type and Xac-KX33).
The relative susceptibilities of ΔdrrAXac, Xac306 and Xac-KX33 strains were determined by the agar diffusion technique to seven growth inhibitors (tetracycline, penicillin G, ampicillin, chloramphenicol, acriflavine, ethidium bromide and doxorubicin chloridrate). No significant difference in drug susceptibility was observed among the three Xac strains (Table 1). In addition, the Northern blotting analyses showed that drrA gene was not required for Xac resistance to these seven drugs since drrA gene expression was not detected after exposure to each drug (data not shown). Hence, the disruption of the drrA gene does not affect the in vitro susceptibility to these drugs under the conditions employed.
| Table 1: | Diameter measurement (cm2) of bacterial growth inhibition zones |
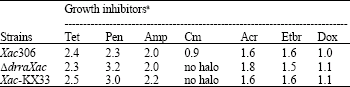 | |
| aThe stock solution concentrations impregnated on sterile filter disks were: tetracycline (Tet), 10 μg μL-1; penicillin (Pen), 50 μg μL-1; ampicillin (Amp), 50 μg μL-1; chloramphenicol (Cm), 100 μg μL-1; acriflavine (Acr), 20 μg μL-1; ethidium bromide (Etbr), 10 μg μL-1 and doxorubicin chloridrate (Dox), 4 μg μL-1. The reported values are representative of two experiments | |
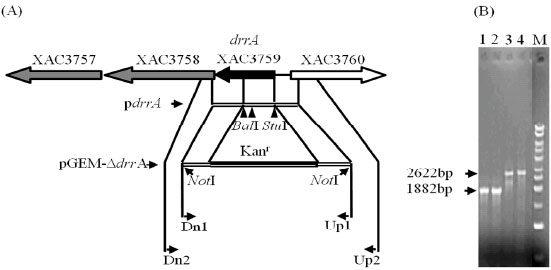 | |
| Fig. 1: | Disruption of drrAXac by allelic exchange with a kanamycin resistant cassette. (A) The diagram illustrates the steps of the disruption cartridge assembling for a ΔdrrA deletion mutant construction. The region of drrA gene cluster was amplified by primers Dn1 and Up1 and cloned into pGEM®-T Easy vector to give rise to plasmid pdrrA. The pdrrA was digested with BalI and StuI and the fragment was substituted by a kanamycin resistance cartridge (Kanr) resulting in pGEM-ΔdrrA carrying the disruption cartridge. Arrowed boxes represent the direction of the putative genes in the Xac genome. (B) PCR amplification of genomic DNA of Xac strain transformed with pKX33-ΔdrrAXac vector. Primers used were Up2 and Dn2. (lanes 1, 2) Xac306 strain (control); (lanes 3, 4) ΔdrrAXac strain and (M) Kilobase DNA marker (GE Healthcare) |
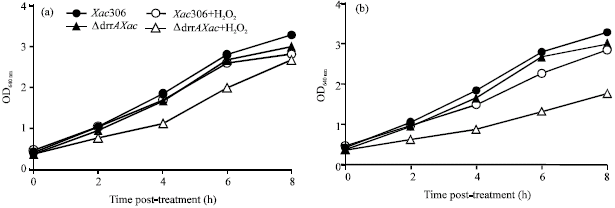 | |
| Fig. 2: | Effect of H2O2 treatment on Xac306 and ΔdrrAXac cultures. H2O2 was added to bacterial culture at early-phase growth (OD640nm≤0.5) to record the absorbance at different times after exposition to oxidative stress. (A) 1 mM H2O2 and (B) 2 mM H2O2. The experiment was assayed in triplicate and a single representative experiment is illustrated here |
As drrA has not seemed to be involved with Xac drug resistance, we assessed the role of drrA in Xac response to H2O2 exposure as a simulation of the induced stress conditions at the pathogen-host interaction. For so, we assayed ΔdrrAXac mutant and control strains under different experimental conditions of H2O2 concentration and bacterial growth phases. The results showed that when H2O2 was added at exponential-phase culture (OD640 nm>0.5) mutant and wild strains were similarly tolerant to both H2O2 concentrations assayed (1 mM and 2 mM H2O2) data not shown. However, when H2O2 was added at early-phase growth (OD640 nm ≤ 0.5) ΔdrrAXac strain was more sensitive to oxidative stress than the wild type showing that drrA gene disruption altered the response to oxidative stress in the mutant strain, an effect dependent on H2O2 concentration (Fig. 2). Northern blotting analyses showed that drrA Xac306 gene is up-regulated after 20 min of exposition to different H2O2 concentrations, as a short and quick response, only at early phase of bacterial growth.
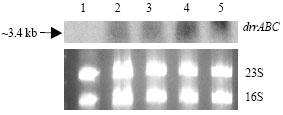 | |
| Fig. 3: | Northern analysis of the drrABC operon using total RNA from Xac306 strain after 20-minute exposure to 1.5 mM (lane 2), 2 mM (lane 3), 3 mM (lane 4) and 6 mM of H2O2 (OD640 nm ≤0.5). Lane 1 corresponds to the untreated culture. A co-transcript of about 3.4 kb, including XAC3759 (drrA gene), XAC3758 (drrB gene) and XAC3757 (drrC gene), was revealed using the BalI-StuI fragment of drrA gene (see Fig. 1A) as the probe. Ethidium bromide-stained rRNA bands are shown for comparison of the quantities of loaded RNA |
(OD640 nm ≤ 0.5) (Fig. 3). At exponential and stationary phases of bacterial growth and in the absence of H2O2 treatment, the drrA gene expression was not detected.
Due to the oxidative tolerance decrease as a consequence of drrA disruption, we tested if the drrA mutation had any effect on growth and survival of Xac in plants. However, unexpectedly, no difference in host symptom development or vesicle size was observed between ΔdrrAXac and control strains when bacterial suspensions were inoculated on leaves of C. sinensis. Typical lesions of citric canker began to be visible in all inocula simultaneously after 5 days. The collapse of the infiltrated area was followed by a premature abscission of the inoculated leaves (Fig. 4A). Also, the monitoring of the Xac population in leaves of C. sinensis during the first sixteen days post-inoculation revealed no variation in growth rate in vivo or in vitro (Fig. 4B). Thus, these results suggest that drrA was not essential for the process of Xac pathogenicity in the plant model tested here.
It has been reported that to several bacteria including Xanthomonas species (Farr and Kogoma, 1991; Mongkolsuk et al., 2005), at exponential and stationary phases of bacterial culture, an enzymatic detoxification mechanism is induced to promote the cellular protection against harmful metabolic products of essential cellular process. Multiple isozymes are induced at later bacterial cultures. Isozymes as catalases (KAT) are induced by carbon excess and by the lack of nutrients during the late phases of bacterial culture and superoxide dismutase (SOD) is responsible for the natural elimination of peroxides and superoxides produced by essential aerobic cell metabolism (Farr and Kogoma, 1991; Fridovich, 1997).
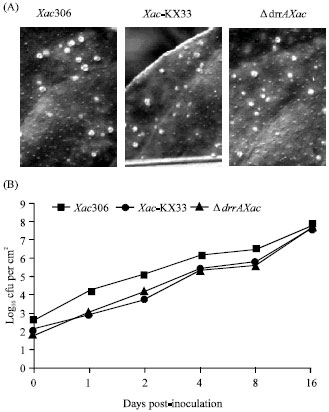 | |
| Fig. 4: | In vivo infection assays. (A) Comparison of canker symptoms (light spots on surface leaves) evoked by the ΔdrrAXac, Xac306 and Xac-KX33 strains 10 days post-inoculation in C. sinensis. (B) Total population of bacteria per cm2 of C. sinensis tissue leaves at different times post-inoculation. The result illustrated here is a single representative experiment from replayed experiments |
Then, indirectly, the action of these enzymes enhances the tolerance to several stress form including subsequent H2O2 stimuli as a cross-protection response. Thus, we presume that the DrrA transporter plays an important role in modulating susceptibility to oxidative stress before the enzymatic detoxification is activated, at early phase of bacterial growth, followed by the inhibition of Xac drrA gene expression when the enzymatic detoxification mechanism is induced.
Furthermore, in plant-pathogen interaction process, the Hypersensitive Response (HR) has been described as the first pathogen-induced defense host response (Bonas and Van den Ackerveken, 1999). HR is triggered in resistant plants by specific plant-pathogen recognition followed by a wide range of physiological changes including ROS production (Lamb and Dixon, 1997). Considering the negative results obtained by in vivo assays to evaluate the essentiality of drrA gene to growth and the survival of Xac in plant host, we supposed that neither plant-pathogen recognition nor ROS took place due to the lack of HR activation in consequence of C. sinensis susceptibility to Xac infection. Consequently, without oxidative stress induction, drrA expression was not required and the mutation effect could not be assessed. However, we can not rule out the possibility of the drrA gene playing a role in bacterium-environmental interaction as part of a defense system against environmental oxidative stress conditions and not being required for bacterium-host interaction.
Interestingly, the gene expression analysis revealed a long RNA transcript with a length of almost 3400 pb, what we deduced to be the co-transcription of three ORFs (XAC3759, drrA gene, 723 pb; XAC3758, hypothetical protein, 1437 pb and XAC3757, hypothetical protein, 1269 pb) as an operon cluster (operon drrABC) (Fig. 1, 3). The sequence analysis and topology prediction of the deduced proteins from these two neighboring hypothetical ORFs (XAC3758 and XAC3757) show that they have 6 transmembrane segments which are characteristic of permeases, supporting the structure of ABC transporter systems with an ABC subunit and two permeases.
CONCLUSION
The growth phenotype observed for ΔdrrAXac mutant suggests that the drrAXac gene is not essential for the survival of Xac in the absence of oxidative stress. Moreover, we presume that DrrA transporter plays an important role in modulating susceptibility to oxidative stress in Xac and that the two co-transcripted hypothetical ORFs should participate of this process.
ACKNOWLEDGMENTS
This study was supported by grants from the Brazilian agencies FAPESP, FAEPA, CNPq and CAPES. We thank R.A. Ferreira; M. Mazucato and A.C.C. Souza for technical assistance and A. Borghi for reviewing the English manuscript.
REFERENCES
- Baldini, R.L., S.T. Tahara and Y.B. Rosato, 1999. A rolling-circle miniplasmid of Xanthomonas campestris pv. glycines: The nucleotide sequence and its use as a cloning vector. Plasmid, 42: 126-133.
CrossRefDirect Link - Biemans-Oldehinkel, E., M.K. Doeven and B. Poolman, 2006. ABC transporter architecture and regulatory roles of accessory domains. FEBS. Lett., 580: 1023-1035.
Direct Link - Bonas, U. and G. Van den Ackerveken, 1999. Gene-for-gene interactions: Bacterial avirulence proteins specify plant disease resistance. Curr. Opin. Microbiol., 2: 94-98.
CrossRef - Brencic, A. and S.C. Winans, 2005. Detection of and response to signals involved in host-microbe interactions by plant-associated bacteria. Microbiol. Mol. Biol. Rev., 69: 155-194.
PubMed - Burse, A., H. Weingart and M.S. Ullrich, 2004. The phytoalexin-inducible multidrug efflux pump AcrAB contributes to virulence in the fire blight pathogen, Erwinia amylovora. Mol. Plant Microb Interact., 17: 43-54.
Direct Link - Da Silva, A.C., J.A. Ferro, F.C. Reinach, C.S. Farah and L.R. Furlan et al., 2002. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature, 417: 459-463.
Direct Link - Daniels, M.J., C.E. Barber, P.C. Turner, M.K. Sawczyc, R.J. Byrde and AH. Fielding, 1984. Cloning of genes involved in pathogenicity of Xanthomonas campestris pv. campestris using the broad host range cosmid pLAFR1. EMBO. J., 3: 3323-3328.
Direct Link - Dassa, E. and P. Bouige, 2001. The ABC of ABCS: A phylogenetic and functional classification of ABC systems in living organisms. Res. Microbiol., 152: 211-229.
Direct Link - Farr, S.B. and T. Kogoma, 1991. Oxidative stress responses in Escherichia coli and Salmonella typhimurium. Microbiol. Rev., 55: 561-585.
Direct Link - Ferreira, H., F.J.A. Barrientos, R.L. Baldini and Y.B. Rosato, 1995. Electrotransformation of three pathovars of Xanthomonas campestris. Applied Microbiol. Biotechnol., 43: 651-655.
Direct Link - Fridovich, I., 1997. Superoxide anion radical (O2), superoxide dismutases and related matters. J. Biol. Chem., 271: 18515-18517.
PubMed - Lamb, L. and R.A. Dixon, 1997. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 251-275.
CrossRefPubMedDirect Link - Mongkolsuk, S., J.M. Dubbs and P. Vattanaviboon, 2005. Chemical modulation of physiological adaptation and cross-protective responses against oxidative stress in soil bacterium and phytopathogen, Xanthomonas. J. Ind. Microbiol. Biotechnol., 32: 687-690.
Direct Link - Mulder, N.J., R. Apweiler, T.K. Attwood, A. Bairoch and D. Barrell et al., 2003. The InterPro Database, brings increased coverage and new features. Nucleic Acids Res., 31: 315-318.
Direct Link - Piddock, L.J., 2006. Multidrug-resistance efflux pumps-not just for resistance. Nat. Rev. Microbiol., 4: 629-636.
Direct Link - Ren, Q., K.H. Kang and I.T. Paulsen, 2004. TransportDB: A relational database of cellular membrane transport systems. Nucleic Acids Res., 32: 284-288.
Direct Link - Ren, Q. and I.T. Paulsen, 2005. Comparative analyses of fundamental differences in membrane transport capabilities in prokaryotes and eukaryotes. PLOS. Comput. Biol., 1: e27-e27.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Verniere, C.J., T.R. Gottwald and O. Pruvost, 2003. Disease development and symptom expression of Xanthomonas axonopodis pc. citri in various citrus plant tissues. Phytopathology, 93: 832-843.
Direct Link - White, T.J. and C.F. Gonzalez, 1991. Application of electroporation for efficient transformation of Xanthomonas campestris pv. oryzae. Phytopathology, 81: 521-524.
Direct Link