
R. Jahanian
Department of Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad 91775-1163, Iran
H. Nassiri Moghaddam
Department of Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad 91775-1163, Iran
A. Rezaei
Department of Immunology, Faculty of Medicine, Isfahan University of Medical Sciences, Isfahan, Iran
A.R. Haghparast
Department of Veterinary Immunology, College of Veterinary Sciences, Ferdowsi University of Mashhad, Mashhad, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 2 | Page No.: 321-327







ABSTRACT
The present study was performed to evaluate the effects of dietary zinc-methionine (ZnMet) supplementation on broiler performance and carcass characteristics. Three added zinc levels (40, 80 and 120 mg kg-1) from each of zinc sulfate or ZnMet were used in a 2x3 factorial arrangement with four replicates of ten birds. Two hundred-forty day-old broiler chicks were fed with the experimental diets from 1 to 42 days of age and at the final day of experiment two randomly selected birds from each replicate were slaughtered and carcass parameters were measured. Inclusion of ZnMet into the diets caused to significant (p<0.01) increase in feed intake. Almost in all cases, increasing supplemental Zn level as either Zn sulfate or ZnMet sources lowered feed consumption. Body weight gain was affected (p<0.01) by zinc source in all experimental periods, with the highest weight gains assigned to chicks fed on ZnMet-supplemented diets. Except for week 1, feed conversion efficiency was not affected by Zn source or dietary Zn concentration. Increasing supplemental Zn level from 40 to 80 mg kg-1 from both Zn sources caused increase in liver weight percentage, but this parameter was reversely affected by further increase to 120 mg added Zn kg-1 of diet. Pancreas, heart and thigh weight percentages were not affected by dietary treatments; however, carcass and breast meat percentages were increased by dietary ZnMet supplementation. The present results suggest that dietary supplementation with more available Zn sources can improve production economics.
PDF Abstract XML References Citation
How to cite this article
R. Jahanian, H. Nassiri Moghaddam, A. Rezaei and A.R. Haghparast, 2008. The Influence of Dietary Zinc-Methionine Substitution for Zinc Sulfate on Broiler Chick Performance. Journal of Biological Sciences, 8: 321-327.
DOI: 10.3923/jbs.2008.321.327
URL: https://scialert.net/abstract/?doi=jbs.2008.321.327
DOI: 10.3923/jbs.2008.321.327
URL: https://scialert.net/abstract/?doi=jbs.2008.321.327
INTRODUCTION
Zinc is commonly supplemented to diets for livestock and poultry, because many natural feed ingredients are marginally Zn-deficient (Cao et al., 2002). Although we do not yet fully understand the element’s metabolism, however, Zn homeostasis and kinetics is extremely complex compared with other essential trace elements. The model with multiple components and time lags describing Zn kinetics in humans proposed by Foster et al. (1984) is evidence of the highly intricate nature of this trace element, although a similar model was not found for avian models. The reason for this complexity may be, in part, because Zn is only essential trace mineral that has a filled 3 days shell and thus has no unpaired electrons (Chesters, 1997) and is necessary for so many diverse enzymatic reactions and other physiological processes (Sandoval et al., 1998).
Numerous experiments have been conducted during the last 50 years to estimate the bioavailability of Zn in supplemental sources and dietary ingredients; however, more than those with other trace elements, these studies have proven to be fraught with various difficulties (Baker and Ammerman, 1997). Some researchers (Spears, 1989; Wedekind et al., 1992) have reported greater bioavailability for organic Zn sources than that observed for inorganic forms, including Zn oxide and Zn sulfate; consequently, organic forms of this nutritionally essential trace element have been used with increasing frequency by the feed industry. It is believed that Zinc-methionine (ZnMet), an organic complex of Zn, provides a source of Zn with greater bioavailability than Zn from inorganic sources such as Zn oxide (ZnO) and Zn carbonate (Wedekind et al., 1990). However, research by Hill et al. (1986) and Pimentel et al. (1991) with pigs and chicks, respectively, indicated no differences in Zn bioefficacy among inorganic and organic Zn sources.
The purpose of the present study was to compare ZnMet and equal additional levels of Zn sulfate for their influences on broiler performance and carcass characteristics.
MATERIALS AND METHODS
Animals, dietary treatments and management: The present study was performed in the experimental farm of Ferdowsi University of Mashhad, Mashhad, Iran. A total of two hundred-forty 1-day-old, cloacal sexed, male RossxRoss broiler chicks were assigned to 24 floor pens in a completely randomized design. There were four pen replicates of ten birds in each of six treatment combinations. The basal corn-soybean meal diets (Table 1) containing 25.9 and 24.2 mg kg-1 zinc (by analysis; as fed basis) for starter and grower stages, respectively, were formulated to meet or exceed nutrimental requirements of broiler chicks (NRC, 1994) except of Zn. Zinc was removed from mineral premix so that the basal diets were contained minimum amount of this element. Dietary treatments consisted of the basal diet supplemented with 40, 80 or 120 mg kg-1 added Zn as either ZnSO4.H2O (as commonly used Zn source) or ZnMet (as organic source; Zinpro Corporation, Edina, MN).
| Table 1: | Chemical composition of basal diets (%) |
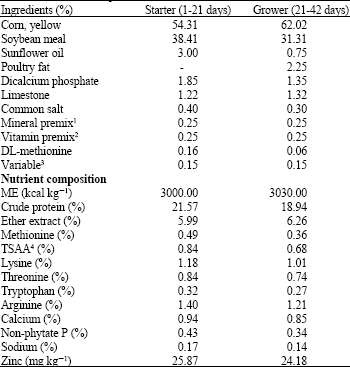 | |
1: Zinc-free mineral premix. Provided per kilogram of diet: Mn (from MnSO4.H2O), 60 mg; Fe (from FeSO4.7H2O), 50 mg; Cu (from CuSO4.5H2O), 6 mg; I (from Ca (IO3)2.H2O), 1 mg; Se, 0.20 mg. | |
Experimental Zn concentrations were achieved by replacing an appropriate amount of inert filler (washed builder sand) with each of the above Zn sources. All of the diets were calculated to contain equal concentration of methionine and other nutrients except of Zn. Chicks were maintained on a 24 h constant-lighting schedule in the floor pens containing painted partitions which placed in a thermostatically-controlled room. Chicks were allowed ad libitum access to feed and tap water from plastic instructions to minimize environmental Zn contamination. At 1 day of age, the temperature was set at 33°C and then the temperature was reduced by 3°C/week until the birds were 4 weeks old. Feed intake and body weight were measured weekly and mortality was recorded daily to adjust feed: gain data. At 42 days of age, two birds from each pen replicate were selected randomly, then weighed and slaughtered by cervical cutting. Liver, pancreas and heart were precisely removed immediately after slaughtering and weighed separately (prior to weighing, heart cleaned of adhered fat). Carcass eviscerated manually. After the eviscerated weight had been obtained, breast and thigh were cut and weighed to evaluate the effects of dietary organic Zn supplementation on meat yield and carcass efficiency.
Chemical analysis: Prior to formulating the diets, the ingredients and Zn sources used in the study herein were analyzed for crude protein (Kjeltec Auto 1030 Analyzer, Tecator), ether extract, crude fiber and ash content according to standard methods of AOAC (1995), then the metabolizable energy content of ingredients was estimated using NRC (1994) recommended formula. Experimental diets were formulated by using these more accurate values. Zinc concentration in feed ingredients and Zn sources were determined by atomic absorption spectrophotometry (Perkin Elmer, Precisely AAnalyst 200, Absorption spectrophotometer). Samples of feedstuffs and Zn sources were dried at 105°C for 16 h, ashed at 550°C for 16 h and solubilized in HCl. All samples were filtered through 42 Whatman filter paper and brought to an appropriate volume with deionized water, then give to spectrophotometer instrument (Anonymous, 1982). Zinc sources were also analyzed for other trace elements using spectrophotometric method.
Statistical analysis: Data were analyzed by two-way ANOVA with the general linear model procedures of SAS (1999), using a model that included dietary Zn source and Zn concentration as the main effects and their interaction. Pen was the experimental unit. Duncan’s multiple range tests (Duncan, 1955) was used to compare treatment means at p<0.05 significant level.
RESULTS AND DISCUSSION
Zinc sources used in the present study (zinc sulfate and zinc-methionine) contained 32.46 and 9.31% zinc, respectively. Sulfate source was Zn sulfate monohydrate and consisted of 1486 mg Fe2+, 27 mg Cu2+ and 1572 mg Mn2+ kg-1 of preparation.
As shown, daily feed intakes in the all trial weeks and in the all experimental periods were highly significantly (p<0.01) affected by Zn source (Table 2). Dietary supplementation with ZnMet led to significant increase in feed intake in the all experimental stages. Increasing supplemental Zn level from either sources caused to significant decline in daily feed consumption in starter (p<0.01), grower and entire experimental period (p<0.001). Incremental levels of ZnMet followed by the higher depression in feed intake than those observed by sulfate, resulted in signifying Zn source by Zn level interaction in grower (p<0.05) and entire trial period (p<0.01). This trend did not reach to significant level in week 6 (p = 0.0623). Currently, there are two feed-grade Zn sources commonly used by the animal feed industry: ZnO (72% Zn) and ZnSO4.H2O (36% Zn). Of the supplemental zinc fed, 80 to 90% is ZnO, which is less bioavailable for poultry than reagent-grade or feed-grade ZnSO4.H2O (Wedekind and Baker, 1990; Sandoval et al., 1997; Edwards and Baker, 1999). In production animals, recently, organic zinc sources such as ZnMet or Zn-propionate were more bioavailable than inorganic sources such as ZnO or ZnSO4.H2O (Spears, 1989; Wedekind et al., 1992; Hahn and Baker, 1993); consequently, organic forms of the element have been used with increasing frequency by the feed industry (Batal et al., 2001). According to our findings, Sahin et al. (2005) showed that zinc-picolinate (ZnPic) as an organic Zn source was more effective than ZnSO4.H2O. The higher absorption rate of ZnPic (Hahn and Baker, 1993) and ZnMet sources allows lower inclusion rates of zinc supplementation and makes mineral balance in animals easier to maintain. This is reasonable explanation why feed intake was more reduced by the highest level of ZnMet as compared with Zn sulfate in the study presented here. Probably, the higher decline in feed intake in ZnMet-supplemented group than sulfate-supplemented ones can be contributed to greater absorption rate of former. In the study by Sandoval et al. (1998), feeding diets containing 1000 mg Zn kg-1 continuously for 3 weeks caused decreased feed intake and reduced growth. Highly available Zn sources should theoretically be more toxic when ingested at high levels, as absorption represents a major portion of bioavailability in the case of Zn (Ammerman et al., 1995). This theoretical effect appeared to be the case for ZnMet, which decreased feed intake to a greater extent than sulfate. In contrast, Oh et al. (1979) reported little effect of dietary Zn on feed consumption of chicks until 4000 mg kg-1 Zn as acetate was fed. The biochemical mechanism by which high dietary Zn decreases feed intake and subsequent growth remains unclear, but the concentration of the element in brain does not appear to be an effective factor (Sandoval et al., 1998).
As noted in Table 3, dietary supplementation with ZnMet caused highly significant (p<0.01) increase in body weight gain in the all experimental stages. Higher levels of supplemental Zn from either sources were associated with numerical (p>0.05) slower weight gain in weeks 1-4; however, weight gains were increased by raising supplemental Zn level from 40 to 80 mg kg-1 in weeks 5 and 6 despite of lower feed intake in the given weeks.
| Table 2: | Effects of zinc source and dietary zinc concentration on daily feed intake of male broiler chicks during entire 42 days experimental period (g day-1 per bird) |
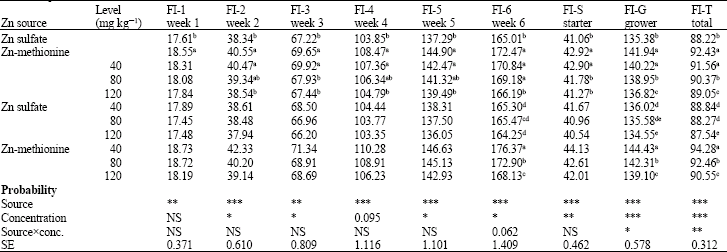 | |
| FI-1, -2, -3, -4, -5, -6, -S, -G, -T refer to feed intake during weeks 1, 2, 3, 4, 5, 6, starter, grower and entire experimental periods, respectively. a-e: Means with no common superscript within a column of each classification (Zn source, Zn level, or SourcexConcentration) differ significantly (p<0.05). NS: Not Significant, *: p<0.05, **: p<0.01, ***: p<0.001 | |
| Table 3: | Effects of zinc source and dietary zinc concentration on daily body weight gain of male broiler chicks during entire 42 days experimental period (g day-1 per bird) |
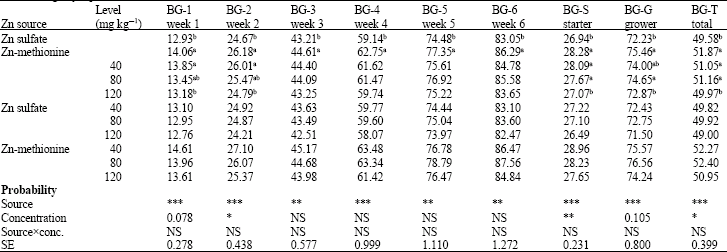 | |
| BG-1, -2, -3, -4, -5, -6, -S, -G, -T refer to body weight gain during weeks 1, 2, 3, 4, 5, 6, starter, grower and entire experimental periods, respectively. ab: Means with no common superscript within a column of each classification (Zn source, Zn level, or SourcexConcentration) differ significantly (p<0.05). NS: Not Significant, *: p<0.05, **: p<0.01, ***: p<0.001 | |
| Table 4: | Effects of zinc source and dietary zinc concentration on feed conversion ratio (g of feed g-1 of gain) of male broiler chicks during entire 42 days experimental period |
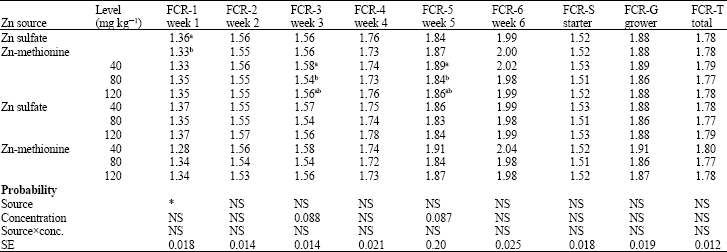 | |
| FCR-1, -2, -3, -4, -5, -6, -S, -G, -T refer to feed conversion ratio during weeks 1, 2, 3, 4, 5, 6, starter, grower and entire experimental periods, respectively. ab: Means with no common superscript within a column of each classification (Zn source, Zn level, or SourcexConcentration) differ significantly (p<0.05). NS: Not Significant, *: p<0.05 | |
Further increase in Zn level to 120 mg kg-1 had a reverse effect. Overall, dietary inclusion of ZnMet increased growth performance by approximately 5%, while if the highest level of supplemental Zn (120 mg kg-1) was omitted from calculations (data not shown), this advantageous effect was more pronounced. The greater body weight gain in chicks fed on diets containing ZnMet as the Zn source may be due to more available Zn supplied by this product. Wedekind et al. (1992) reported results of two experiments in which ZnMet was supplemented to maize-soybean meal diets for chicks from 8 to 22 days posthatching. These authors calculated a Relative Bioavailability Value (RBV) for ZnMet of 2.06 compared to 1.00 for Zn sulfate. Of course, only the control and two lowest levels of supplementation were included in the regression of total tibia Zn on supplemental Zn intake due to curvature in the response. When data for all treatments were included in a model to estimate a value that would be more representative of supplemental amounts of Zn used by the poultry feed industry, the RBV estimate dropped to 1.40.
Except of week 1 in which dietary ZnMet supplementation improves (p<0.05) feed conversion ratio, FCR data was not affected by dietary added Zn source or concentration, or interaction between them (Table 4). In agreement with the present results, Wedekind et al. (1994) reported that inclusion of organic zinc sources into the swine diets had no beneficial effect on feed efficiency. Similarly, Sandoval et al. (1999) observed that feed conversion from day 15 through day 21 was not affected (p>0.10) by the dosing method or Zn source.
| Table 5: | Effects of zinc source and dietary zinc concentration on relative weights of some body organs and carcass efficiency of chicks at 42 days of age (% of live body weight) |
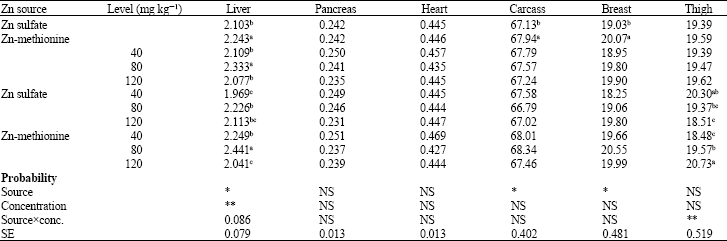 | |
| abc: Means with no common superscript within a column of each classification (Zn source, Zn level, or SourcexConcentration) differ significantly (p<0.05). NS: Not Significant, *: p<0.05, **: p<0.01 | |
Lack of improvement in feed conversion efficiency by dietary inclusion of ZnMet may be due to higher feed intake from ZnMet-supplemented diets.
The effects of dietary supplementation with different Zn levels on organ weight and carcass characteristics are seen in Table 5. Liver weight percentage was affected by dietary supplemental zinc source (p<0.05) and concentration (p<0.01). Inclusion of ZnMet into the diets caused significant increase (p<0.05) in liver weight percent. Increasing supplemental Zn level from 40 to 80 mg kg-1 as either sulfate or ZnMet sources was associated with heavier liver weight; however, further increase to 120 mg Zn kg-1 reversely affected this parameter. Reduction of liver weight percentage by dietary zinc level of 120 mg kg-1 was more dramatic with ZnMet source, caused interaction between zinc source and zinc level came near to significant level (p = 0.0866). The responsibility of liver weight to dietary zinc alterations may be due to the critical role of this organ in Zn metabolism. Transfer of Zn to liver from plasma is 5-6 times faster than transfer to other major tissues (House and Wastney, 1997).
Pancreas, heart and thigh weight percentages were not affected by supplemental zinc source or level; however, carcass and breast were heavier (p<0.05) in ZnMet-supplemented groups. The greater liver, carcass and breast weight percentages in chicks fed on ZnMet-supplemented diets may be related to higher absorption rate and consequently more available zinc supplied by this supplement. Tissue Zn turnover varies among tissues and is a factor in metabolism of Zn and in its excretion from the body. Rapidly turning over tissues such as liver can respond more rapidly to the changes in Zn intake and alterations in dietary utilizable (bioavailable) zinc content (Buckley, 2000). On the other hand, tracer studies indicate that increasing dietary Zn level and in turn, excessive Zn intake leads to a reduction in tissue tracer specific activity or retention. These findings indicate that tissue Zn turnover is decreased in Zn deficiency status and increased when Zn supply is excessive (Windisch and Kirchgessner, 1994). These observations can be an explanation why increasing Zn level to 120 mg kg-1 had reverse effect on liver and pancreas weight percentages. The probable reason for heavier breast and carcass weights in ZnMet-supplemented birds (more bioavailable zinc source compared with sulfate) is participation of Zn in DNA and protein biosynthesis. Zinc is present in all cells and participates in a wide variety of metabolic processes by virtue of its diverse catalytic roles in over 200 enzymes. Zn enzymes are involved in the biosynthesis and/or degradation of carbohydrates, lipids, proteins and nucleic acids and encompass all known classes of enzymes (Falchuk and Vallee, 1985; Kaim and Schwederski, 1994). Its deficiency arrests proliferation in all cells, while in multicellular organisms it also results in abnormal differentiation and development leading to extensive teratological abnormalities (Falchuk and Vallee, 1985).
From the present results it can be concluded that zinc-methionine provides more bioavailable zinc than feed-grade zinc sulfate. Although some reports indicate that higher levels of supplemental zinc than those studied here had no considerable adverse effect on feed intake; however, increasing supplemental Zn level to 120 mg kg-1 in the present study caused significant decline in feed consumption. In summary, use of ZnMet in poultry diets seems to lower zinc requirements and to improve appetite and growth performance of broiler chicks. In addition, the greater bioefficacy for zinc-methionine relative to zinc sulfate in enhancing chick performance suggests that the metabolism of former complex differs from metabolism of Zn supplied by inorganic zinc sources.
ACKNOWLEDGMENTS
The authors thank Dr. Ajilchi (The chairman of Sanadam Pars Co., Iran) for providing zinc-methionine supplement. We also thank Excellent Center of Animal Science and Deputy for Research in Ferdowsi University of Mashhad, Mashhad, Iran.
REFERENCES
- AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Batal, A.B., T.M. Parr and D.H. Baker, 2001. Zinc bioavailability in tetrabasic zinc chloride and the dietary zinc requirement of young chicks fed soy concentrate diet. Poult. Sci., 80: 87-90.
Direct Link - Cao, J., P.R. Henry, S.R. Davis, R.J. Cousins, R.D. Miles, R.C. Littell and C.B. Ammerman, 2002. Relative bioavailability of organic zinc sources based on tissue zinc and metallothionein in chicks fed conventional dietary zinc concentrations. Anim. Feed Sci. Technol., 101: 161-170.
CrossRefDirect Link - Edwards, H.M. and D.H. Baker, 1999. Bioavailability of zinc in several sources of zinc oxide, zinc sulfate and zinc metal. J. Anim. Sci., 77: 2730-2735.
CrossRefDirect Link - Foster, D.M., M.E. Wastney and R.I. Henkin, 1984. Zinc metabolism in humans: A kinetic model. Math. Biosci., 72: 359-372.
CrossRefDirect Link - Hahn, J.D. and D.H. Baker, 1993. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci., 71: 3020-3024.
CrossRefDirect Link - Hill, D.A., E.R. Peo, Jr., A.J. Lewis and J.D. Crenshaw, 1986. Zinc-amino acid complexes for swine. J. Anim. Sci., 63: 121-130.
PubMed - House, W.L. and M.E. Wastney, 1997. Compartmental analysis of zinc kinetics in mature male rats. Am. J. Physiol., 273: R1117-R1125.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Oh, S.H., N. Nakaue, J.T. Deagen, P.D. Whanger and G.H. Arscott, 1979. Accumulation and depletion of zinc in chick tissue metallothioneins. J. Nutr., 109: 1720-1729.
Direct Link - Pimentel, J.L., M.E. Cook and J.L. Greger, 1991. Research note: Bioavailability of zinc-methionine for chicks. Poult. Sci., 71: 1637-1639.
CrossRefDirect Link - Sahin, K., M.O. Smith, M. Onderci, N. Sahin, M.F. Garsu and O. Kucuk, 2005. Supplementation of zinc from organic or inorganic source improves performance and antioxidant status of heat-distressed quail. Poult. Sci., 84: 882-887.
CrossRefDirect Link - Sandoval, M., P.R. Henry, C.B. Ammerman, R.D. Miles and R.C. Littell, 1997. Relative bioavailability of supplemental inorganic zinc sources for chicks. J. Anim. Sci., 75: 3195-3205.
Direct Link - Sandoval, M., P.R. Henry, X.G. Luo, R.C. Littell, R.D. Miles and C.B. Ammerman, 1998. Performance and tissue zinc and metallothionein accumulation in chicks fed a high dietary level of zinc. Poult. Sci., 77: 1354-1363.
Direct Link - Sandoval, M., P.R. Henry, R.C. Littell, R.D. Miles, G.D. Butcher and C.B. Ammerman, 1999. Effect of dietary zinc source and method of oral administration on performance and tissue trace mineral concentration of broiler chicks. J. Anim. Sci., 77: 1788-1799.
Direct Link - Spears, J.W., 1989. Zinc methionine for ruminants: Relative bioavailability of zinc in lambs and effects of growth and performance of growing heifers. J. Anim. Sci., 67: 835-843.
Direct Link - Wedekind, K.J. and D.H. Baker, 1990. Zinc bioavailability in feed-grade sources of zinc. J. Anim. Sci., 68: 684-689.
Direct Link - Wedekind, K.J., A.E. Hortin and D.H. Baker, 1992. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate and zinc oxide. J. Anim. Sci., 70: 178-187.
PubMedDirect Link - Wedekind, K.J., A.J. Lewis, M.A. Giesemann and P.S. Miller, 1994. Bioavailability of zinc from inorganic and organic sources for pigs fed corn-soybean meal diets. J. Anim. Sci., 72: 2681-2689.
PubMed