
M. A. Al-Kahtani
Department of Biology, College of Science, King Faisal University, Saudi Arabia
A. A. Fathi
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1226-1230







ABSTRACT
The effects of microcystin produced by the toxic cyanobacterial strain Microcystis aeruginosa on bioaccumulation and antioxidant enzymes (superoxide dismutase and catalase) of tilapia fish (collected from Al-Khadoud spring, Al-Hasa, Saudi Arabia) were investigated. The results showed that microcystin contained in cyanobacterial blooms induce CAT and SOD activity in a time-dependent manner. The data also shows that microcystin concentration in muscle was much lower than in liver, as the liver is the target organ of these toxins. In addition, microcystin concentration in faeces increased gradually until the end of the treatment period.
PDF Abstract XML References Citation
How to cite this article
M. A. Al-Kahtani and A. A. Fathi, 2008. Physiological Studies on Tilapia Fish (Oreochromis niloticus) as Influenced by the Cyanobacterial Toxins Microcystin . Journal of Biological Sciences, 8: 1226-1230.
DOI: 10.3923/jbs.2008.1226.1230
URL: https://scialert.net/abstract/?doi=jbs.2008.1226.1230
DOI: 10.3923/jbs.2008.1226.1230
URL: https://scialert.net/abstract/?doi=jbs.2008.1226.1230
INTRODUCTION
The cyanobacteria, are a major group of bacteria that occur throughout the world. Some species of cyanobacteria produce toxins which are classified, according to their mode of action, such as hepatotoxins (e.g., microcystins), neurotoxins (e.g., anatoxins) and skin irritants. The hepatotoxins are produced by various species within the genera Microcystis, Anabaena, Oscillatoria, Nodularia, Nostoc and Cylindrospermopsis. Most hepatotoxins (all cyclic heptapeptides) are microcystins. The chemical structure of microcystins includes two variable amino acids and an unusual aromatic amino acid, ADDA (3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid), containing a substituted phenyldecadienoic acid. Microcystin-LR is a cyclic heptapeptide with a relative molecular mass of about 1000 (Carmichael, 1988, 1999).
Microcystins bioaccumulate in common aquatic vertebrates and invertebrates, including fish (Sipia et al., 2001; Magalhaes et al., 2003; Mohamed et al., 2003), mussels (Falconer et al., 1992; Prepas et al., 1997; Watanabe et al., 1997) and zooplankton (Thostrup and Christoffersen, 1999; Mohamed, 2001). Williams et al. (1997) have shown covalent binding and accumulation of microcystin-LR in salmon liver and crab larvae. Whether the levels of microcystin accumulation are sufficient to pose a risk to humans is uncertain and this probably will depend on levels of consumption and the severity of toxic blooms in the area where fish or shellfish are caught or collected. Common advice given by water authorities is that the viscera of the fish should not be eaten, and caution should be taken in all cases where major toxic blooms occur.
The acute effects of microcystin in fish on inhibition of PP1 and 2A in liver have been evaluated in several studies. Sahin et al. (1995) fed trouts with M. aeruginosa extracts containing microcystin-LR and showed 50% inhibition of protein phosphatases activity with 0.05 nM of toxin. Tencalla and Dietrich (1997) observed complete inhibition of proteins phosphatase with 2 μg of microcystin-LR g-1 in the liver of trouts. Soares et al. (2004) studied the accumulation and depuration of microcystins (cyanobacteria hepatotoxins) in Tilapia rendalli (Cichlidae) under laboratory conditions. Mohamed and Hussein (2006) studied the accumulation of microcystins in different organs, particularly the liver, of tilapia fish (Oreochromis niloticus) in an Egyptian fish farm containing toxic blooms of Microcystis aeruginosa. Mohamed and Al Shehri (2007) investigated the cyanobacteria and their toxins in treated-water storage reservoirs in Abha city, Saudi Arabia. Recently, Mohamed (2008) studied the toxic cyanobacteria and cyanotoxins in public hot springs in Saudi Arabia.
Eutrophic water ecosystems favour the intense development of cyanobacteria blooms. These microorganisms are potentially toxins producers able to damage aquatic biota and mammals. Oreochromis sp. is an important group of fish species in water springs of Al-Hasa, Saudi Arabia. Moreover, they are commonly found in brackish water in estuaries around the world and respond promptly to environmental alterations (Almeida et al., 2002). This study aims to investigate the effects of microcystin produced by cyanobacterial strain Microcystis aeruginosa on some metabolic activities of tilapia fish collected from Al-Khadoud spring, Al-Hasa, Saudi Arabia.
MATERIALS AND METHODS
Organism and culture conditions: The toxic strain Microcystis aeruginosa was isolated from Al-Asfar Lake. Al-Asfar Lake is one of the important shallow wetland lakes. It is located on the eastern region of Saudi Arabia, Al-Hasa Province. This strain was cultured in ASM-1 medium (Gorham et al., 1964), pH = 8.0, temperature of 23±2 C, 22 μE m-2 sec-1 of light intensity and photoperiod of 12 h. At the end of exponential growth phase, cells were concentrated by centrifugation and keep on the freeze.
Fish sampling: Tilapia (Oreochromis niloticus) fish were collected by bottom trap net by fishermen from Al-Khadoud spring channel during May 2007. Al-Khadoud spring is one of the most important water resources in Al-Hassa region and plays an important role in agricultural activities in the area. It is located nearly 5.0 km Northwest of King Faisal University main campus. The fish were transported to the laboratory in a container with spring drainage water.
Experiment design: Studies were conducted using tilapia fish with mean weight 50.62±10.0 g. The fish were grown (40 fish) in aquariums 90 L-1 at room temperature (25±2 °C). Fish were fed with commercial fish food and were acclimatized for 10 days before the beginning of the experiments. Tilapia fish were exposed to microcystin by feeding with cyanobacterial cells under laboratory conditions. In a test aquarium fish food and dried algal cells dusted on the water surface, daily for 10 days (one time). In accordance with the dose of toxin chosen (approximately 50.0 μg Microcystin/fish/day) and the concentration of microcystin contained in the cyanobacterial cells (3.5 mg g-1). Control fish were grown under the same conditions in a separate tank. Groups of five fish were sacrificed every two days. The experiment period was 10 days. Three replicates for each treatment were used on these experiments.
Extraction and analysis of MCYSTs from fish tissues and faeces: Livers, gallbladder and intestines of each (every two days) sacrificed group of fish were separately homogenized in 90% methanol, stirred overnight at room temperature and then centrifuged at 5000 rpm. The pellets were re-extracted with the same procedure. The supernatants of each organ were combined and diluted with a known volume of distilled water (100 mL). The organic solvent was dried by sterilized air. The aqueous fractions remaining after removing the organic solvent and the daily sampled tank water were applied to C18 silica cartridges. The toxin was then eluted with 80% methanol and microcystine content that remained in the aqueous fraction after methanol evaporation was determined by both enzyme-linked immunosorbent assay (ELISA) and protein phosphatase inhibition assay (PPIA) according to Carmichael and An (1999). The value of each parameter in this study is the mean of five readings. Faeces were collected with a glass pipette from the bottom of each aquarium in. It was dried at 50°C, weighed and extracted twice. Microcystine concentration in the faeces material was also determined using ELISA plate kits according to Carmichael and An (1999).
Determination of antioxidant enzymes: Catalase (CAT) activity was assayed by the method of Beers and Sizer (1952). Superoxide dismutase (SOD) activity was measured using the xanthine oxidase-cytochrome c method as described by McCord and Fridovich (1969). The activities of both enzymes were determined just two times, after 5 and 10 days from the incubation period.
RESULTS AND DISCUSSION
The production of microcystins (Fig. 1) in aquatic environments has been associated with a decline in microorganism growth (Ostensvik et al., 1998; Bloor and England, 1989), a reduction in animal and algal community diversity (Aboal et al., 2000, 2002) and even their bioaccumulation in the food chain (Lauren-Määttä et al., 1995).
The data of (Fig. 2) shows that no notable effects were observed in CAT activity of liver or kidney of tilapia fishes after 5 days of exposure, but the activity increased in liver and kidney after 10 days of treatment (Fig. 2). On the other hand, the activity of CAT in gills was lower than that in the other two
tested organs. Regards to SOD activity the data of (Fig. 3) shown that there is no difference on the activity of SOD in liver, kidney and gills of fish that had been exposed to dried cyanobacteria cells for 5 days. However the longer exposure (10 days) resulted in a significant increase in the SOD activity only in liver and gills, whereas induction in kidney was not appeared (Fig. 3). But the SOD activity in gills for day 10 is lower than the control. In general, the results also show that microcystin contained in cyanobacterial blooms induce CAT and SOD activity in a time-dependent manner.
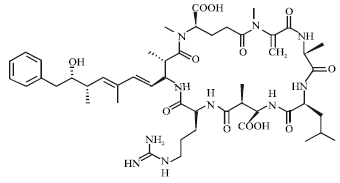 | |
| Fig. 1: | Chemical structure of microcystin |
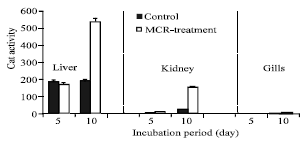 | |
| Fig. 2: | CAT (nmol H2O2 consumed/min/mg protein) activity in liver, kidney and gills of tilapia fish exposed to dried cyanobacterial cells after 5 and 10 days from the investigation period. Vertical bars indicate SE, n = 3 |
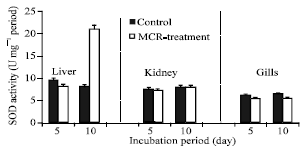 | |
| Fig. 3: | SOD (U mg-1 protein) activity in liver, kidney and gills of tilapia fish exposed to dried cyanobacterial cells after 5 and 10 days from the investigation period. Vertical bars indicate SE, n = 3 |
As far as SOD and CAT activities are concerned, a simultaneous induction response is usually observed after exposure to pollutants (Dimitrova et al., 1994). Li et al. (2003) reported enhanced SOD and CAT activities in the hepatocytes of common carp (Cyprinus carpio L.), induced by MC-LR. Recently, Jos et al. (2005) reported that when tilapia fish were exposed to cyanobacterial cells under laboratory conditions the antioxidant enzymes (SOD, CAT, GPx and GR) activities were induced. The results of this investigation suggest that the antioxidant enzymes are induced as a defense mechanism (Jos et al., 2005). On the other hand, the liver was the most affected organ in all cases because microcystins are potent hepatotoxins in mammals and fish (Fisher and Dietrich, 2000; Towner et al., 2002; Jos et al., 2005).
| Table 1: | Microcystins concentrations in liver muscle and faeces of Tilapia (Oreochromis niloticus) |
 | |
The kidney was the next most affected organ. This agrees with the results obtained by other researchers who reported kidney lesions induced by MC-LR in fish (Rabergh et al., 1991; Kotak et al., 1996; Fisher and Dietrich, 2000; Jos et al., 2005). Finally, gills were the least affected organs. Some laboratory studies have shown that the gill and skin epithelia of freshwater fish form a barrier to microcystin transport (Tencalla et al., 1994; Bury et al., 1995) and, consequently, ingestion may be the most probable route for microcystin uptake in fish (Bury et al., 1998).
Microcystin accumulation in liver muscle and faeces of tilapia fish during the incubation period can be observed in (Table 1). The data performed that the highest concentration in liver was observed on the fourth day, when it was detected 0.72 μg g-1. However, from the 10th day of incubation the microcystin concentration in liver decreased. The data of (Table 1) also shows that microcystin concentration in muscle was much lower than in liver, certainly because liver is the target organ of these toxins. The highest concentration in muscle during the accumulation period (0.028 μg g-1) was observed on the sixth day of the incubation period.
The major exposure of fish and zooplankton to microcystin is through ingestion of toxic cyanobacterial cells or their toxins released during cell lysis (Carbis et al., 1996; Mohamed, 2001; Mohamed and Hussein, 2006). Microcystis is rapidly taken up by hepatocytes through carrier-mediated transport. Once in the hepatocytes, it binds covalently and irreversibly to the catalytic subunit of protein phosphatases 1 and 2A (Williams et al., 1997; Bischoff, 2001; Mohamed and Hussein, 2006). The results obtained with this study demonstrated that microcystin accumulation in liver and muscle of tilapia fish occurred with different profiles. Further decreases of microcystin concentration in the tissues might also be associated to excretion of microcystin through the faeces. On the other hand, Microcystin concentration in faeces increased gradually until the end of the treatment period. The highest concentration observed on the last day was 0.110 μg g-1 per fish (Table 1). The excretion of microcystin through faeces has already been reported in the literature and can be due to bile excretion, considered as the main excretion via for these toxins (Falconer and Humpage, 1996; Rabergh et al., 1991; Sahin et al., 1996; Jos et al., 2005). Decreases of microcystin concentration in the tissues might also be associated to excretion of microcystin through the faeces. Many earlier studies have been detected microcystin in muscle of fish (Mohamed et al., 2003; Mohamed and Hussein, 2006). All those studies verified levels in muscle that would not be recommended for human consumption
ACKNOWLEDGMENT
We thank the Deanship of Scientific Research, King Faisal University for financial support (Grant No. 8063).
REFERENCES
- Aboal, M., M.A. Puig, P. Mateo and E. Perona, 2002. Implications of cyanophyte toxicity on biological monitoring of calcareous streams in North-East Spain. J. Applied Phycol., 14: 49-56.
Direct Link - Almeida, J.A., Y.S. Diniz, S.F.G. Marques, L.A. Faine, B.O. Ribas, R.C. Burneiko and E.L.B. Novelli, 2002. The use of the oxidative stress responses as biomarkers in Nile tilapia (Oreochromis niloticus) exposed to In vivo cadmium contamination. Environ. Int., 27: 673-679.
Direct Link - Beers, Jr. R.F. and I.W. Sizer, 1952. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem., 195: 133-140.
CrossRefPubMedDirect Link - Bloor, S. and R.R. England, 1989. Antibiotic production by the cyanobacterium Nostoc muscorum. J. Applied Phycol., 1: 367-372.
CrossRef - Fisher, W.J. and D.R. Dietrich, 2000. Pathological and biochemical characterization of microcystin-induced hepatopancreas and kidney damage in carp (Cyprinus carpio). Toxicol. Applied Pharmacol., 164: 73-81.
CrossRefDirect Link - Jos, A., S. Pichardo, A.I. Prieto, G. Repetto, C.M. Vazquez, I. Moreno and A.M. Camean, 2005. Toxic cyanobacterial cells containing microcystins induce oxidative stress in exposed tilapia fish (Oreochromis sp.) under laboratory conditions. Aquat. Toxicol., 72: 261-271.
CrossRefDirect Link - Li, X., Y. Liu, L. Song and J. Liu, 2003. Responses of antioxidant systems in the hepatocytes of common carp (Cyprinus carpio L.) to the toxicity of microcystin-LR. Toxicon, 42: 85-89.
CrossRefDirect Link - Magalhaes, V.F., M.M. Marinho, P. Domingos, A.C. Oliveira and S.M. Costa et al., 2003. Microcystins (cyanobacteria hepatotoxins) bioaccumulation in fish and crustaceans from Sepetiba Bay (Brasil, RJ). Toxicon, 42: 289-295.
Direct Link - McCord, J.M. and I. Fridovich, 1969. Superoxide dismutase: An enzymic function for erythrocuprein (Hemocuprein). J. Biol. Chem., 244: 6049-6055.
PubMedDirect Link - Mohamed, Z.A., 2001. Accumulation of cyanobacterial toxins by Daphnia in some Egyptian irrigation canals. Ecotoxicol. Environ. Saf., 50: 4-8.
CrossRefDirect Link - Mohamed, Z.A., 2008. Toxic cyanobacteria and cyanotoxins in public hot springs in Saudi Arabia. Toxicon, 51: 17-27.
CrossRefDirect Link - Mohamed, Z.A. and A. Hussein, 2006. Depuration of microcystins in tilapia fish exposed to natural populations of toxic cyanobacteria: A laboratory study. Ecotoxicol. Environ. Saf., 63: 424-429.
Direct Link - Mohamed, Z.A., W.W. Carmichael and A.A. Hussein, 2003. Estimation of microcystins in the freshwater fish Oreochromis niloticus in an Egyptian fish farm containing a Microcystis bloom. Environ. Toxicol., 18: 137-141.
Direct Link - Mohamed, Z.A. and A.M. Al-Shehri, 2007. Cyanobacteria and their toxins in treated-water storage reservoirs in Abha city, Saudi Arabia. Toxicon, 50: 75-84.
CrossRefDirect Link - Ostensvik, O., O.M. Skulberg, B. Underdal and V. Hormazabal, 1998. Antibacterial properties of extracts from selected planktonic freshwater cyanobacteria-a comparative study of bacterial bioassays. J. Applied Microbiol., 84: 1117-1124.
CrossRefDirect Link - Rabergh, C.M.I., G. Bylund and J.E. Eriksson, 1991. Histopathological effects of microcystin-LR, a cyclic peptide toxin from the cyanobacterium (blue-green alga) Microcystis aeruginosa, on common carp (Cyprinus carpio L.). Aquatic Toxicol., 20: 131-146.
CrossRefDirect Link - Sipia, V.O., H.T. Kankaanpaa, J. Flinkman, K. Lahti and J.A. Meriluoto, 2001. Time-dependent accumulation of cyanobacterial hepatotoxins in flounders (Platichthys flesus) and mussels (Mytilus edulis) from the northern Baltic Sea. Environ. Toxicol., 16: 330-336.
Direct Link - Soares, R.M., V.F. Magalhaes and S.M.F.O. Azevedo, 2004. Accumulation and depuration of microcystins (cyanobacteria hepatotoxins) in Tilapia rendalli (Cichlidae) under laboratory conditions. Aquatic. Toxicol., 70: 1-10.
Direct Link - Towner, R.A., S.A. Sturgeon and K.E. Hore, 2002. Assessment of in vivo oxidative lipid metabolism following acute microcystin-LR-induced hepatotoxicity in rats. Free Radic. Res., 36: 63-71.
Direct Link