
S.S. Ellerton
Department of Biological Sciences and Geology, Queensborough Community College, Bayside, New York, USA
A.J. Litewka
Department of Biology, Brooklyn College, Brooklyn, New York, USA
M. Gonzalez
Department of Biology, Brooklyn College, Brooklyn, New York, USA
C.K. Jue
Department of Biological Sciences and Geology, Queensborough Community College, Bayside, New York, USA
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1211-1215







ABSTRACT
The GDP-mannose transporter gene VRG4 has been identified from a number of yeasts including Saccharomyces cerevisiae. The vrg4-2 allele of the GDP-mannose transporter of S. cerevisiae has been primarily characterized as having general phenotypes affecting glycosylation. In this study, vrg4-2 was characterized, specifically with regard to defects in cell wall biosynthesis. The cell wall in vrg4-2 mutants was found to be profoundly compromised, as measured by the spheroplast lysis assay. These mutant cells exhibited increased sensitivity to Congo red which is believed to interfere with chitin-glucan assembly in the cell wall. An increased level of cell wall fluorescence was also observed when the cells were incubated with Calcofluor white suggesting an increased level of chitin in the wall. Despite these apparent major alterations in the cell wall, the vrg4-2 mutants exhibited near normal growth relative to the wild-type. All together, these results suggest that vrg4-2 mutation affects the cell wall structure, but less so in general cellular metabolism.
PDF Abstract XML References Citation
How to cite this article
S.S. Ellerton, A.J. Litewka, M. Gonzalez and C.K. Jue, 2008. The GDP-Mannose Transporter is Required for Cell Wall Integrity in Saccharomyces cerevisiae. Journal of Biological Sciences, 8: 1211-1215.
DOI: 10.3923/jbs.2008.1211.1215
URL: https://scialert.net/abstract/?doi=jbs.2008.1211.1215
DOI: 10.3923/jbs.2008.1211.1215
URL: https://scialert.net/abstract/?doi=jbs.2008.1211.1215
INTRODUCTION
The cell wall of Saccharomyces cerevisiae is primarily composed of an array of mannoproteins (mannan) and glucans linked to small amounts of chitin (Duran and Nombela, 2004). Although this complex structure is often represented as rigid, it is actually highly dynamic, capable of modifying its own structure in response to changing physiological conditions and environmental stresses. If the cell wall`s normal structure is altered (stressed) such as through antifungal drug echinocandins treatment or cell wall gene mutation or deletion, cells typically respond by incorporating increased levels of chitin which presumably strengthens the cell wall (Imai et al., 2005; Klis et al., 2002; Popolo et al., 1997; Walker et al., 2008).
Mannoproteins are synthesized and processed in the ER and Golgi complex after which they are transported to and incorporated into the cell surface. The core mannose groups are N- and O-linked to the wall proteins in the endoplasmic reticulum and terminal mannosylation is completed in the Golgi apparatus (Gemmill and Trimble, 1999).
The substrate for protein mannosylation in the Golgi is the activated sugar, GDP-mannose. GDP-mannose, which is synthesized in the cytoplasm, must be transported, through a sugar transporter, into the lumen of the organelle where protein mannosylation occurs (Dean et al., 1997). The gene which codes for the GDP-mannose transporter (VRG4) was first isolated from S. cerevisiae by Poster and Dean (1996). Subsequently, her group isolated two further orthologs from two different species of Candida (Nishikawa et al., 2002a, b). More recently, orthologs of the GDP-mannose trans- porter have also been isolated from Pichia pastoris (Arakawa et al., 2006), Cryptococcus neoformans (Cottrell et al., 2007) and Aspergillus nidulans (Jackson-Hayes et al., 2008). In non-yeast species, VRG4 orthologs are found in plants, such as Arabadopsis thaliana (Baldwin et al., 2001) and two species of rice (Oryza sativa) cDNA (Poster and Dean, 1996).
Saccharomyces cerevisiae vrg4-2 mutant cells are defective in the production of mannoproteins. Such cells produce shortened versions of the glycoproteins invertase and chitinase, as evidenced by migration on gel electrophoresis (Dean et al., 1997; Poster and Dean, 1996). Presumably, all mannoproteins produced by these cells will be defective. Consequently, we reasoned that this mutant should also have defective cell wall phenotypes. In this study, we investigated the changes in the integrity and structure of the cell wall of vrg4-2 mutant cells as characterized by a Zymolyase lysis assay and through the binding of two dyes: Congo red and Calcofluor white.
MATERIALS AND METHODS
Yeast strains, media and general methods: The yeast strains used in this study are listed as follows: RSY255: MATα ura3-52 leu2-211; NDY5: MATα ura3-52 leu2-211 vrg 4-2. SEY6210: MATα ura3-52 his3-Δ200 trp1-Δ 901 lys2-801 suc2-Δ 9 leu2-3, 112. All strains were provided as gifts from Dr. Neta Dean of Stony Brook University (SUNY). The strains were maintained on YPAD agar as described by Adams et al. (1997). The strains were grown in YPAD liquid media at 30°C with shaking unless otherwise stated.
Measurement of spheroplast formation and cell lysis by Zymolyase: Spheroplast formation assay was as described by Lim et al. (1995). Briefly, cells were grown in liquid YPAD until mid-log phase (0.3 OD) Cells were washed and suspended in 3 mL CE buffer (0.1 M sodium citrate, 10 mM EDTA, pH 6.25) in 13x100 mm glass tubes. Three microliters of Zymolyase at a concentration of 5 U μL-1 (Zymo Research) was added to each tube and mixed. Every 3 min the OD at 660 nM was measured using a Spectronic 20 spectrophotometer. The decrease of OD value was taken as cell lysis.
Effect of Congo red on cell growth: Sensitivity to Congo red assay has been described by Ram and Klis (2006). Congo red was dissolved in sterile water at 50 mg mL-1. Fifty milliliters of YPAD agar buffered with 100 mM MES-NaOH pH 6.0 was melted and maintained at 50°C in a water bath. An appropriate aliquot of Congo red stock was added to the melted agar to 50 μg mL-1. The Congo red containing agar was poured into an 8.5x12.5 cm sterile plastic plate. The agar was then allowed to solidify at room temperature. Cells were adjusted to 5x106 cells mL-1. The stock was further serially diluted by 10 fold four times. Ten microliters of each dilution was spotted onto the Congo red and control plates which were then incubated at 30°C for 48 h. Sensitivity to Congo red was determined by visually comparing colony density between wild type and mutant cells.
Cell wall chitin content: Calcofluor white binding to yeast cell walls was adapted from Pringle (1991). 2x107 exponential-phase yeast cells were centrifuged at 3,000 rpm and the pellet was washed once with water. The washed pellet was resuspended in 50 μg mL-1 Calcofluor white and incubated for 10 min at room temperature. The cells were again pelleted, washed twice with water and the final pellet was resuspended in water. Cells were observed using brightfield and fluorescent microscopy with an Olympus BX51 microscope. The fluorescence from Calcofluor white was filtered with a DAPI filter.
Growth curves: Cells diluted with YPAD to OD600 ~ = 0.05 were grown in YPAD liquid culture at 30°C. Growth curves were generated by measuring the absorbance at 600 nm every 1.5 h for 9 h and after 24 h.
RESULTS AND DISCUSSION
Vrg4-2 mutant cells are hypersensitive to Zymolyase: Zymolyase has a β-1,3-glucanase activity that hydrolyzes glucose polymers at the β-1,3-glucan linkages, as well as protease activity (Zlotnik et al., 1984). Yeast cells treated with Zymolyase experience a gradual degradation of their cell walls to form cell wall deficient spheroplasts. Unlike intact cells that have lysis resistant cell walls, spheroplasts will undergo lysis in hypoosmotic medium. The sensitivity of the cell wall to Zymolyase will vary depending on the cell wall structure. Cell walls with altered structure (i.e., fewer mannan groups or mannan more accessible to hydrolysis) should be more susceptible to these lytic enzymes and will therefore more readily undergo lysis than normal walls. Cell lysis can be measured as a reduction in optical density at 660 nM and has been used as a sensitive assay for disruption of cell wall integrity by a number of laboratories (Herrero et al., 2002; Lim et al., 1995). As shown in Fig. 1, incubation of vrg4-2 cells with Zymolyase results in a 25% reduction in absorbance after 3 min, a 50% reduction after 5 min and maximal reduction in absorbance (80%) after 20 min. Wild-type cells formed spheroplasts at a much slower rate, with only a 10% reduction in absorbance after 15 min and a 32% reduction after 30 min. The rate of lysis for vrg4-2 cells is quite dramatic, not only in comparison to wild type, but even in comparison to published results for other cell wall deficient mutants such as gda1 (Herrero et al., 2002).
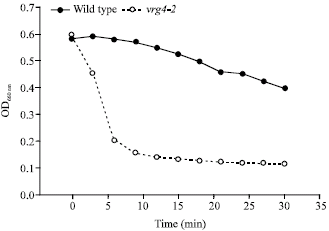 |
| Fig. 1: | Sensitivity of vrg4-2 to Zymolyase. Wild-type and vrg4-2 cells in midlog phase were incubated with Zymolyase. The optical density at 660 nm (OD660) was measured every 3 min |
Clearly vrg4-2 cells rapidly form spheroplasts in the presence of Zymolyase. The results of this experiment do not identify the cell wall components that play a part in this process nor resolve which cell wall components have been altered or if there is increased accessibility of cell wall components to the lytic enzymes. However, these results, together with the known function of the gene, strongly suggest that hypomannosylation in the cell walls of vrg4-2 mutants result in a weakened cell wall that is more susceptible to lysis.
Other manifestations of the differential structure of mutants versus wild type of VRG4 have been published. An important study by Poster and Dean (1996) determined that vrg4-2 mutant cells exhibit significant differences in the morphology of their internal membranes by both confocal and electron microscopy. Abnormal endomembrane structure would likely interfere with its normal role in the processing, transport and integration of glycoproteins into the cell wall. In the electron micrographs published by this group, one can also observe a difference in the appearance of the cell wall of mutant versus wild-type cells. The outer layer of the cell wall appears thicker in wild-type than in mutant cells. In addition, there is staining of some diffuse material outside of the cell walls of wild-type, but not mutant, cells.
VRG4 mutant cells exhibit increased sensitivity to Congo red: Congo red is an anionic dye that interferes with cell wall assembly, likely by binding to cell wall chitin (Imai et al., 2005; Ram and Klis, 2006). This binding prevents chitin molecules from forming cross-linkages with other cell wall components, thereby destabilizing the cell wall (Imai et al., 2005). Other strains of yeast with cell wall abnormalities exhibit increased levels of chitin in their cell walls (Popolo et al., 1997) and cell wall mutants have previously been characterized based on their sensitivity to Congo red (Ram and Klis, 2006).
When incubated with this dye, wild-type cells exhibit a very mild inhibition of growth at a density of 5x101 cells while vrg4-2 cells exhibit inhibition of growth at a cell density of 5x103 cells with no growth of colonies at lower cell concentrations (Fig. 2). This sensitivity to Congo red would be expected if vrg4-2 cells incorporate increased levels of chitin into the cell wall as a compensatory mechanism to mitigate the effects of hypoglycosylation.
Vrg4-2 mutant cells exhibit increased levels of binding to Calcofluor white: Calcofluor white is an anionic dye that binds to chitin in yeast cell walls (Ram and Klis, 2006). Although this dye is not specific for chitin, its binding has been used to visualize chitin content and localization in cell walls (Imai et al., 2005). In wild-type cells, this dye binds significantly to bud necks, sites of bud emergence, bud scars and to a lesser extent, the lateral cell walls, as shown in Fig. 3A. Figure 3C illustrates that many of the vrg4-2 cells exhibit an increase in fluorescence of the lateral cell walls, as compared to wild-type cells (Fig. 3A). As reported by Francois (2006), many cell wall defective mutants, such as fks1, mnn9, gas1, kre6 and krn4, increase the chitin content of the cell wall, presumably to increase cross linking as compensation for quantitative and/or qualitative reductions in cell wall glycoproteins.
 |
| Fig. 2: | Inhibition of vrg4-2 growth in the presence of Congo red. vrg4-2 and wild-type cells (5x106 cells mL-1) were serially diluted (1:10) and plated (10 μL) on YPAD agar plates, with and without Congo red (50 μl mL-1). Plates were incubated for 48 h at 30°C |
Vrg4-2 growth approximates that of wild-type cells: As shown in Fig. 4, vrg4-2 cells exhibit a growth rate similar to that of wild type cells. Although we have shown that the vrg4-2 cells have cell wall defects (Fig. 1, 2) it is interesting that their rate of growth remains unaffected. Normal growth rate in this type of mutant suggests that the most profound effects may be limited to cell wall structure, with relatively little effect on general cellular metabolism.
Indeed, in spite of the profound cell wall effects observed in this study, some significant functions dependent on cell wall structure seem to remain intact. One of the more important functions of the cell wall in yeasts involves mating. These vrg4-2 mutants are mating type α, which produces a sex-specific cell wall glycoprotein called α-agglutinin. This molecule facilitates mating with the opposite mating type (a cells) by promoting adhesion to the corresponding a-agglutinin on the surface of a cells (Terrance et al., 1987). An increase in the concentration of these agglutinins at the cell surface is induced in each cell type under the influence of a sex pheromone produced by the opposite type (Terrance and Lipke, 1987). The resultant sex-specific adhesion increases the efficiency of mating. A fusion protein of α-agglutinin green fluorescence protein was constructed and its incorporation into the cell walls of the mutant and wild type cells was measured (Gonzalez and Lipke, 2003). There was no apparent difference in the amount of fluorescence incorporated into the mutant versus the wild type cells. Therefore, VRG4 is not required for anchoring mannoproteins onto the cell wall.
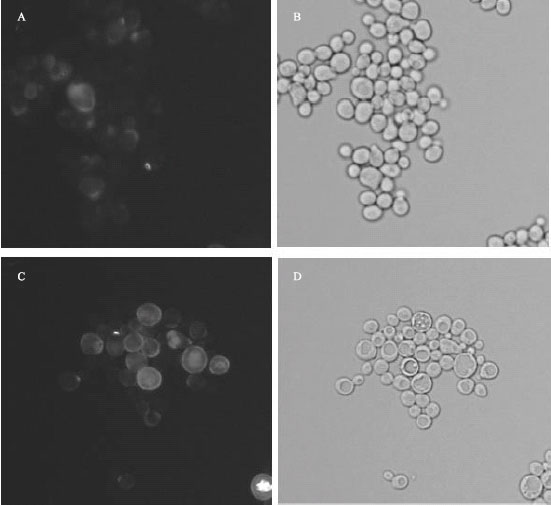 |
| Fig. 3: | Fluorescent staining of chitin by Calcofluor white in wild type and vrg4-2 cells. Mutant and wild-type cells (2x107) were incubated with 50 μg mL-1 calcofluor white and visualized by fluorescent and brightfield microscopy. Fluorescent images of wild-type (A) and vrg4-2 cells (C) are compared with brightfield images of wild type (B) and vrg4-2 cells (D). All cells were viewed with a 60X oil immersion lens |
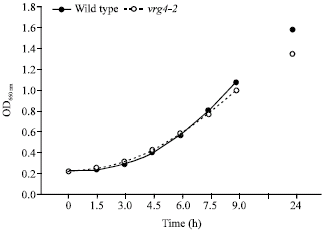 |
| Fig. 4: | Growth curve of vrg4-2 cells. The growth curve of vrg4-2 cells and wild-type cells in YPAD medium at 30°C |
ACKNOWLEDGMENTS
This research program was supported by the PSC-CUNY grants (PSCREG-38-17 and PSCREG-39-1053) awarded to Dr. Chong Jue. This work was partially supported by the NIGMS-SCORE under grant S06 GM076168 to Brooklyn College. We wish to thank Dr. Neta Dean of Stony Brook University for providing the yeast strains and encouragement throughout this work. We also want to thank Dr. Peter Lipke of Brooklyn College for generously providing laboratory space, reagents and helpful discussions in preparation of this study.
REFERENCES
- Arakawa, K., M. Abe, Y. Noda, H. Adachi and K. Yoda, 2006. Molecular cloning and characterization of a Pichia pastoris ortholog of the yeast Golgi GDP-mannose transporter gene. J. Gen. Applied Microbiol., 52: 137-145.
CrossRefDirect Link - Baldwin, T.C., M.G. Handford, M.I. Yuseff, A. Orellana and P. Dupree, 2001. Identification and characterization of GONST1, a Golgi-localized GDP-mannose transporter in Arabidopsis. Plant Cell, 13: 2283-2295.
Direct Link - Cottrell, T.R., C.L. Griffith, H. Liu, A.A. Nenninger and T.L. Doering, 2007. The pathogenic fungus Cryptococcus neoformans expresses two functional GDP-mannose transporters with distinct expression patterns and roles in capsule synthesis. Eukaryot Cell, 6: 776-785.
CrossRefDirect Link - Dean, N., Y.B. Zhang and J.B. Poster, 1997. The VRG4 gene is required for GDP-mannose transport into the lumen of the Golgi in the yeast, saccharomyces cerevisiae. J. Biol. Chem., 272: 31908-31914.
CrossRefDirect Link - Duran, A. and C. Nombela, 2004. Fungal cell wall biogenesis: Building a dynamic interface with the environment. Microbiology, 150: 3099-3103.
CrossRefDirect Link - Francois, J.M., 2006. A simple method for quantitative determination of polysaccharides in fungal cell walls. Nat. Protoc., 1: 2995-3000.
CrossRefDirect Link - Gemmill, T.R. and R.B. Trimble, 1999. Overview of N- and O-linked oligosaccharide structures found in various yeast species. Biochim. Biophys. Acta, 1426: 227-237.
CrossRef - Gonzalez, M. and P.N. Lipke, 2003. An approach to identifying genes required for cross-linking of GPI-mannoproteins to the cell wall in Saccharomyces cerevisiae. Proceedings of the 2nd International Conference on Molecular Mechanisms of Fungal Cell Wall Biogenesis, August 27-September 1, 2003, CSIC/Universidad de Salamanca, Salamanca, Spain, pp: 101-101.
Direct Link - Herrero, A.B., D. Uccelletti, C.B. Hirschberg, A. Dominguez and C. Abeijon, 2002. The Golgi GDPase of the fungal pathogen candida albicans affects morphogenesis, glycosylation and cell wall properties. Eukaryot Cell, 1: 420-431.
CrossRefDirect Link - Imai, K., Y. Noda, H. Adachi and K. Yoda, 2005. A novel endoplasmic reticulum membrane protein Rcr1 regulates chitin deposition in the cell wall of saccharomyces cerevisiae. J. Biol. Chem., 280: 8275-8284.
CrossRefDirect Link - Jackson-Hayes, L., T.W. Hill, D.M. Loprete, L.M. Fay and B.S. Gordon, et al, 2008. Two GDP-mannose transporters contribute to hyphal form and cell wall integrity in aspergillus nidulans. Microbiology, 154: 2037-2047.
CrossRefDirect Link - Klis, F.M., P. Mol, K. Hellingwerf and S. Brul, 2002. Dynamics of cell wall structure in saccharomyces cerevisiae. FEMS Microbiol. Rev., 26: 239-256.
CrossRefDirect Link - Lim, S.T., C.K. Jue, C.W. Moore and P.N. Lipke, 1995. Oxidative cell wall damage mediated by bleomycin-Fe (II) in saccharomyces cerevisiae. J. Bacteriol., 177: 3534-3539.
Direct Link - Nishikawa, A., B. Mendez, Y. Jigami and N. Dean, 2002. Identification of a Candida glabrata homologue of the S. cerevisiae VRG4 gene, encoding the Golgi GDP-mannose transporter. Yeast, 19: 691-698.
CrossRefDirect Link - Nishikawa, A., J.B. Poster, Y. Jigami and N. Dean, 2002. Molecular and phenotypic analysis of CaVRG4, encoding an essential Golgi apparatus GDP-mannose transporter. J. Bacteriol., 184: 29-42.
CrossRefPubMedDirect Link - Popolo, L., D. Gilardelli, P. Bonfante and M. Vai, 1997. Increase in chitin as an essential response to defects in assembly of cell wall polymers in the ggp1delta mutant of saccharomyces cerevisiae. J. Bacteriol., 179: 463-469.
Direct Link - Poster, J.B. and N. Dean, 1996. The yeast VRG4 gene is required for normal golgi functions and defines a new family of related genes. J. Biol. Chem., 271: 3837-3845.
Direct Link - Pringle, J.R., 1991. Staining of bud scars and other cell wall chitin with calcofluor. Methods Enzymol, 194: 732-735.
CrossRefDirect Link - Ram, A.F. and F.M. Klis, 2006. Identification of fungal cell wall mutants using susceptibility assays based on calcofluor white and congo red. Natl. Protoc., 1: 2253-2256.
CrossRefDirect Link - Terrance, K., P. Heller, Y.S. Wu and P.N. Lipke, 1987. Identification of glycoprotein components of alpha-agglutinin, a cell adhesion protein from saccharomyces cerevisiae. J. Bacteriol., 169: 475-482.
Direct Link - Terrance, K. and P.N. Lipke, 1987. Pheromone induction of agglutination in saccharomyces cerevisiae a cells. J. Bacteriol., 169: 4811-4815.
Direct Link - Walker, L.A., C.A. Munro, I. de Bruijn, M.D. Lenardon, A. McKinnon and N.A. Gow, 2008. Stimulation of chitin synthesis rescues Candida albicans from echinocandins. PLoS Pathog.
CrossRefDirect Link - Zlotnik, H., M.P. Fernandez, B. Bowers and E. Cabib, 1984. Saccharomyces cerevisiae mannoproteins form an external cell wall layer that determines wall porosity. J. Bacteriol., 159: 1018-1026.
Direct Link