
A.A . Moazedi
Department of Biology, School of Science, Shahid Chamran University, Ahwaz, Iran
Z. Valizadeh
Department of Biology, School of Science, Shahid Chamran University, Ahwaz, Iran
G. H.A. Parham
Department of Statistic, School of Mathematic, Shahid Chamran University, Ahwaz, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1205-1210







ABSTRACT
The present investigation on the modulatory effects of zinc on NMDA receptors in this study the effects of intra dorsal (CA1) hippocampus injection of NMDA receptor agonist and antagonist to passive avoidance task in the presence and absence of ZnCl2 have been investigated in adult male rats, by using step down apparatus. So, 10 groups (n = 8) rats were considered, first group as a control, the second group received 0.1 μg rat-1 NMDA in 0.5 μL saline for 4 days immediately after shock. Third group receives 1 μg rat-1 MK-801 in 0.5 μL saline 10 min before training for 4 days. Sham groups receive saline at the same condition. Five remain groups received 100 mg/kg/day ZnCl2 in drinking water for 2 weeks. Sixth group only receives 100 mg/kg/day ZnCl2, but other groups (7, 8, 9 and 10), in addition of consumption ZnCl2, receive drug or saline in the same condition to 2, 3, 4, 5 groups. While administration of NMDA improve the impairment effects on ZnCl2 consumption (p<0.05), present results showed that consumption of 100 mg/kg/day ZnCl2 on adult male rat`s impair both learning and memory (p<0.05) but administration of MK-801 increasing the impairment effects of ZnCl2 (p<0.05). It seems that consumption of zinc chloride influence on passive avoidance learning and memory by its effects on NMDA receptor subunits in the hippocampus.
PDF Abstract XML References Citation
How to cite this article
A.A . Moazedi, Z. Valizadeh and G. H.A. Parham, 2008. The Role of CA1 Hippocampus NMDA Receptors to Passive Avoidance Task in Presence and Absence of ZnCl2 in Adult Male Rats. Journal of Biological Sciences, 8: 1205-1210.
DOI: 10.3923/jbs.2008.1205.1210
URL: https://scialert.net/abstract/?doi=jbs.2008.1205.1210
DOI: 10.3923/jbs.2008.1205.1210
URL: https://scialert.net/abstract/?doi=jbs.2008.1205.1210
INTRODUCTION
N-methyl-D-aspartate (NMDA) receptors, a subtype of ionotropic glutamate receptors have been implicated in numeral neural processes. NMDA receptor, For example is involved in long-term potentiation (LTP) which is considered as a physiological base for learning and / or memory (Homayovn et al., 2004; Kawabe et al., 2007). Accumulating evidence indicates that NMDA receptors are importantly involved in formation of memory for aversive conditioning, spatial memory training and nonspatial, nonaversive tasks (Noêmia et al., 2005). NMDA receptors are widely distributed in the brain but they present in high densities in the amygdala basolateral nuclei. The highest concentrations of NMDA binding sites are CA1 of the hippocampus that can, with the substantial concentration, localize in the dentate gyrus (Jafari-Sabet, 2006). NMDA receptors in hippocampus CA1 area are very important in the regulation of synaptic plasticity and the processes of learning and memory including short-and long-term memory (Jafari-Sabet, 2006; Noêmia et al., 2005). Adult brain has shown to contain large amount of chelatable zinc which is predominantly localized to glutamatergic terminals in the hippocampal formation (Chen et al., 1997). Zinc- an essential dietary trace element is the only secondary abundance body`s iron. Zinc is an endogenous metal which plays an important role in the modulation of some central nervous system function. It is mainly found in synaptic vesicles of hippocampus and is considered to be co released with glutamate at these synapses (Chen et al., 1997; De Oliveria et al., 2001). Thus zinc can interact with either inhibitory (GABAergic) or excitatory (glutamatergic) neurotransmitter system (De Oliveria et al., 2001; Molnar and Nader, 2001). Applying at high concentration, it has neurotoxic effects on neuron cultures or when it presents at concentrations below the physiological values, it causes cognitive impairment (De Oliveria et al., 2001). Localization of zinc in the presynaptic area has observed in the hippocampal formation (Homayovn et al., 2004). Presenting of zinc-containing neurons that sequester zinc in the presynaptic vesicles and release it in a calcium and impulse- dependent manner has been demonstrated in the brain (Noêmia et al., 2005). The zinc-containing neurons are thought to form episodic memory function neural circuits (Chen et al., 1997; Keller et al., 2001). Zinc may be important for cognitive operation (Molnar and Nader, 2001; Takeda et al., 2000). Several studies have suggested a functional role for zinc in the mental function. For instance, zinc supplementation improves cognitive behavior in school, age and children (Maureen et al., 2004). A role for zinc as an endogenous modulator of glutamatergic transmission has been suggested; it is, however, zinc effect on the regulation of NMDA receptor activity which is not well understood. So, in this study the effects of administration of agonist (NMDA) and antagonist (MK-801) of NMDA receptor into dorsal hippocampus (intra-CA1) in presence and absence of zinc chloride on adult male rat`s passive avoidance learning and memory have been investigated.
MATERIALS AND METHODS
Animals: Male wistar rats weighting 180±20 had used at the time of the experiment; the animals were individually housed in stainless cages at a temperature of 23±2°C and 12/12 h day-night cycle (7:00 am light on, 7:00 pm light off). All animals were provided from animal facility of Ahwaz Medical Science University. Animal`s food, as follow, was provided from standard Laboratory feed : 5% soybean protein isolate 0.3% DL-methionine, 32.7% corn starch, 25% sucrose, 2% cellulose powder, 5% mineral mixture, 1% vitamin mixture.
Animals had access to food and water ad libitum and were allowed to adapt to the laboratory conditions for at least one week before the experiment. Rats were handling about 3 mines each day prior to behavioral testing. All experiments were performed between 8:00 am and 12:00 pm The time required for this study was 8 months from May 2006 to January 2007. All experiments were performed in the Laboratory of Learning and Memory in department of biology Shahid Chamran University in Iran.
Experimental procedures: In the first experiment rats were divided into 5 groups. The control group with free access to food and water, a second group which received 0.1 μg rat-1 NMDA in a volume of 0.5 μL saline intra-CA1 immediately after shock in step down task for 4 days in training trial (Jafari-Sabet, 2006). Third group which received 1 μg rat-1 MK-801 in volume of 0.5 μL saline intra-CA1 (Jafari-Sabet, 2006) 10 min before training trial for 4 days (Li-Sha et al., 2005). The sham groups were received saline under the same conditions. In the second experiment rats were divided into 5 groups. In this part of study all ones received a daily oral dose of 100 mg kg-1 ZnCl2 for two weeks in drinking water. First group only received 100 mg/kg/day ZnCl2, second group which received 0.1 μg rat-1 NMDA in a volume of 0.5 μL saline intra-CA1 immediately after shock for 4 days in training trial in presence of zinc chloride. Third group which received 1 μg rat-1 MK-801 in volume of 0.5 μL saline intra-CA1 (Jafari-Sabet, 2006) 10 min before training trial for 4 days in presence of zinc chloride. Sham groups which received saline under the same condition. All animals had tested in step down passive avoidance task.
Stereotaxic surgery and microinjection: Animals were anesthetized with intraperitoneal injection (100 mg kg-1) of ketamine hydrochloride plus (10 mg kg-1) xylazine (Flecknell, 1996) and positioned in a Stoelting sterotaxic instrument. A 22 gauge stainless steel guide cannula was placed 2 mm above the intended site of injection. Stereotaxic coordinates for the CA1 regions of dorsal hippocampus were -3.8 mm posterior to bergma, -2.2 mm lateral to the midline and -3 mm ventral of the skull`s dorsal surface. Cannula was, however, secured to anchor jeweler`s screws with dental acrylic. To prevent clogging, stainless steel stylets (27 gauge) had placed in the guide cannula until the animals were given the CA1 injection. All animals were allowed one week to recover from surgery and anesthesia. animals were gently restrained by hand and the styles were removed from the guide cannula and replaced by 27 gauge injection needles (2 mm below the tip of the guide annual). Each injection unit was connected by polyethylene tubing to a 10 μL Hamilton syringe. The right CA1 infused with 0.5 μL solution (0.5 μL rat-1) over a 60 sec period. The injection needles were kept in their places for an additional 60 sec period to allow diffusion and the stylets reinsertion into the guide cannula.
Drug administration: Drugs, in this study, were zinc chloride (Merck, Germany), N-methyl-D-aspartate, (M107-5MG) (+)-MK-801 hydrogen maleate (Sigma). NMDA and MK-801 dissolved in sterile 0.9% saline and injected into the intra-CA1 of hippocampus in a volume of 0.5 μL rat-1.
Passive avoidance apparatuses and procedure: The experimental device is a 30x30x30 cm avoidance-response-chamber, made of Plexiglas on its four sides. The chamber has a bottom of parallel 0.5 cm stainless steel bars spaced 1 cm apart. A rubber platform (5 cm high, 8 cm in diameter at its top surface) was fixed in place in the center at the bottom of the chamber as described by Chen (2005). Rats were continually trained in one-trial step down inhibitory avoidance task for 4 times (one time/day, conducted between 8:00 am and 12:00 pm) and tested for their memory retention at the same 24 h time after training. Rats were placed on the platform and their latency to step down placing and their four paws on the grids had measured.
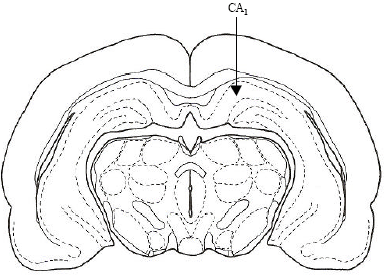 | |
| Fig. 1: | The approximate placement of injection cannula within the CA1 is indicated in figure |
Their times of placing on the platform, At the meantime, were recorded. In training sessions immediately upon stepping down, the rats received a 0.5 mA, 2S, scramble foot shock. No foot shock was given in test sessions. Test session step down latencies and errors (during 3 min) were taken as a measure of memory retention (Chen, 2005).
Histology: After the completion of experimental sessions, each animal was killed with an overdose of chloroform. Animals received a 0.5 μL side-1 injection of ink (1% aqueous methylene blue solution). Their brains were then removed and immersion fixed in a 10% formalin solution for 5 days before sectioning the fixed brains. Brains were sectioned directly across the injection sites and the cannula placement verified using the atlas of Paxino and Watson (2007).The result of histological examination CA1 region of dorsal hippocampus injection cannula placement is shown in (Fig. 1). Data from rats with injection site located outside the CA1 area of the dorsal hippocampus were not used in the analysis.
Statistic analysis: Statistical analysis of data using one way analysis of variance (ANOVA) and post hoc analysis (Tukey test) using the least significant difference (LSD). The decision for statistic test of significance was p<0.05. Calculation were performed using the SPSS statistical package.
RESULTS
The effects of NMDA receptor agonist and antagonist on passive avoidance learning and memory: Present data indicated that, there were significant differences between rats received 0.1 μg rat-1 NMDA immediately after shock in 2,4 days of training and 24 h after training and sham group (p<0.05) (Fig. 2). there were no significant differences between control group and sham group of NMDA in 1, 2, 3, 4 day of training but there were significant differences between these groups 24 h after training (p<0.05) (Fig. 3). Also there were significant differences between rats received MK-801 10 min before training in each five day and sham group (p<0.05) (Fig. 2). Furthermore there were no significant differences between control group and sham group of MK-801 in step-down latencies in any days of training (Fig. 3). The results indicated that NMDA increases passive avoidance learning and memory in adult male rats, but MK-801 as a noncompetitive antagonist decrease learning and memory.
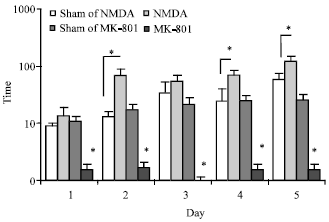 | |
| Fig. 2: | The effects of NMDA receptor agonist and antagonist in absence of ZnCl2 on passive avoidance learning and memory in adult male rats. *p<0.05, n = 8/group |
The effects of ZnCl2 consumption on passive avoidance learning and memory: There were significant differences between rats received 100 mg/kg/day ZnCl2 and control group in 1 and 2 days of training and 24 h after training (p<0.05) (Fig. 4). These data indicate that consumption of ZnCl2 impairs learning and memory in step down task.
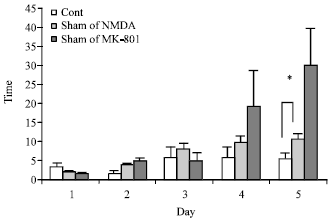 | |
| Fig. 3: | Comparison between control and sham groups in absence of ZnCl2 on passive avoidance learning and memory in adult male rats. * p<0.05, n = 8/group |
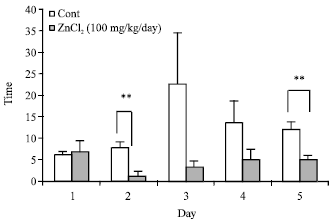 | |
| Fig. 4: | The effects of ZnCl2 (100 mg/kg/day) on passive avoidance learning and memory in adult male rats. ** p<0.01, n = 8/group |
The effects of NMDA receptor agonist and antagonist on passive avoidance learning and memory in presence of zinc chloride: Statistical analysis of data showed that there were significant differences between rats received 0.1 μg rat-1 NMDA+ZnCl2 (100 mg/kg/day) and sham group in fourth day of training and 24 h after training (p<0.05) (Fig. 5) and rats received 1 μg rat-1 MK-801+ ZnCl2 (100 mg/kg/day) and sham group in fourth day of training and 24 h after training (p<0.05) (Fig. 5). There were no significant differences between rats received 100 mg/kg/day ZnCl2 and sham group of NMDA + ZnCl2 (Fig. 6), but There were significant differences between rats received 100 mg/kg/day ZnCl2 and sham group of MK-801+ZnCl2 in second days of training (p<0.05) and 24 h after training (p<0.0) (Fig. 6). Indeed the impairment effects of zinc chloride significantly reduced by NMDA (0.1 μg rat-1 intra-CA1) but increasing by MK-801 (1 μg rat-1 intra-CA1).
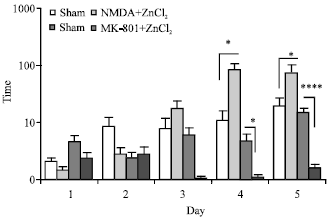 | |
| Fig. 5: | The effects of NMDA receptor agonist and antagonist in presence of ZnCl2 on passive avoidance learning and memory in adult male rats. * p<0.05, **** p<0.000, n=8/group |
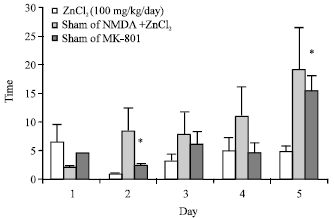 | |
| Fig. 6: | Comparison between control group and sham groups in presence of ZnCl2 on passive avoidance learning and memory in adult male rats.* p<0.05, n = 8/group |
DISCUSSION
The purpose of this study is to investigate the effects of intra-dorsal hippocampal (intra-CA1) administration of N-methyl-D-aspartate receptor agonist and non competitive NMDA receptor antagonist (MK-801) on passive avoidance learning and memory in adult male rats in absence and presence of ZnCl2. The present study shows that NMDA receptor in the CA1 area of dorsal hippocampus interacts with zinc in the modulation of learning memory. The present data indicate that post-training Intra- CA1 administration of NMDA (a NMDA receptor agonist) improves the retention latency. Jafari Sabet indicated that intra dorsal hippocampus injection of NMDA improve learning and memory in shattel box task. This result is in agreement with present result. It has been shown that NMDA receptor activation is involved directly with the primary neural plasticity mechanism underlying the memory formation (Jafari-Sabet, 2006). In spite of such positive findings, some researchers failed to detect a positive effect of NMDA agonists on learning and in some studies, even impairment was observed. present experiments indicate that pre- training intra-CA1 administration of MK-801 a competitive NMDA antagonist reduces the retention latency. Castellano et al. (2001) had shown, pre training administration of MK-801 impaired learning and memory in rats. This result is in agreement with present report. Some study indicated post training injection of MK-801 reduces learning and memory in rat. It is opposite with present result. This contrasting result might be due to difference of time injection (Castellano et al., 2001). Furthermore, some researchers reported an improvement of learning after NMDA receptor antagonists in passive avoidance tasks. These contrasting results might be due to the choice of infusion site (Castellano et al., 2001; Riedel et al., 2003). Moreover, present findings indicated that NMDA receptors are involved not only with acquisition, but also with the consolidation processes.
The present experiments show that zinc chloride consumption 100 mg/kg/day for two weeks, could have impaired learning and memory. This result is similar with the results of many other investigators. For instance Turner et al. (2000) had shown administration of high doses of zinc to rats resulted in spatial reference memory deficit (Turner and Soliman, 2000). Also, Filinn et al. (2005) had shown that enhanced zinc consumption causes memory deficit and increasing brain levels of zinc, because of the influx of toxic amount of zinc from presynaptic vesicles into post-synaptic degeneration neuron seems to be mainly responsible for the neurodegeneration process (Filinn et al., 2005). But some researches indicated that removing zinc from synaptic vesicles didn`t impair spatial learning, memory, or sensorimotor functions in the mouse (Cole et al., 2001). On the other hand in glutamatergic neurons, zinc can also be accumulated into synaptic vesicles and released in order to modulate directly the NMDA and GABA receptors, both of these receptors are essential for memory function (Mocchegiani et al., 2005).
In this experiment we showed that intra- dorsal hippocampal administration of NMDA can improve the impairment effective of zinc consumption but injection of MK-801a non competitive antagonist of NMDA receptor into CA1 increasing the impairment affects of zinc. Zinc may alter brain`s function through changes in neurotransmitter content and receptor activity. For example, it has been shown that zinc negatively modulates N-methyl- D-aspartate (NMDA) receptor activity (Keller et al., 2001). Zinc on the other hand, is believed to be an endogenous modulator, particularly of glutamatergic excitatory synaptic transmission. NMDA receptor- mediated excitation and response induced by activation of metabotropic glutamate receptors can be attenuated by zinc whereas AMPA induced exitary transmission is increased in the presence of zinc. The inhabitation of established long term- potent ion (LTP) by zinc may be accounted for by blocking nerve- terminal calcium channels causing a depression in excitatory post synaptic potential (EPSP) amplitude due to a reduction in glutamate released (Turner and Soliman., 2000). A down- regulate glutamate response for zinc releasing can be performed into two ways: (1) by inhibiting the pre- synaptic release of glutamate via GABA released from interneuron and (2) by reducing the ability of glutamate to activate NMDA receptors, via direct interaction whit NMDA receptors at the post- synaptic membrane level (Mocchegiani et al., 2005). Zinc concentration of the hippocampus and cerebrum, both of which serve important functions in short- term memory, were reduced only in the zinc — restricted rats that demonstrated impair short- term memory (Keller et al., 2001). Zinc is known to inhibit NMDA receptor activity in the hippocampus (Keller et al., 2001; Filinn et al., 2005). Zinc may reduce NMDA receptor- activated channel currents on cortical neurons by acting at 2 different sites, one outside the membrane field and affecting opening frequency and the other inside the channel and interfering directly with the passage of ions (Christine and Choi, 1990). The sensitivity of NMDA receptor to zinc inhibitions depends on its subunit composition. The NR1/NR2A complex has a 2000 fold greater sensitivity to zinc that the NR1/NR2B complex. NR1/NR2B is highly expressed in the immature rat brain and is replaced by the NR1/NR2A complex in the adult brain (Keller et al., 2001; Chen et al., 1997; Herin and Aizenman, 2004). NR1/NR2A, However, exhibits two distinct binding sites for zinc, a voltage- independent, high potency site and a lower potency, voltage dependent site, whereas NR1/NR2B seems to have one, largely voltage-independent, relatively low potency binding site for zinc inhabitation (Chen et al., 1997; Vogt et al., 2004; Molnar and Nader, 2001). Chowanadisai (2005) have shown regulation of NMDA receptor that was controlled by zinc. Zinc deficiency can impair learning and memory in life my be by reduce NMDA receptor (Chowanadisai et al., 2005). However effects of zinc on the regulation of NMDA receptor activity are not well understood.
REFERENCES
- Castellano, C., V. Castari and A. Ciamei, 2001. NMDA receptors and learning and memory processes. J. Curr. Drug Targets., 2: 273-283.
PubMedDirect Link - Chen, N., A. Moshaver and L.A. Ramond, 1997. Differential sensitivity of recombinant N-methyl-d-aspartate receptor subtype to zinc inhibition. J. Mol. Pharm., 51: 1015-1023.
PubMedDirect Link - Chen, Y.G., 2005. Specific tau phosphoryl site hippocampus correlater with impairment of step-down inhibitory avoidance task in rats. J. Behav. Brain. Res., 158: 277-284.
CrossRefDirect Link - Chowanadisai, W., S.L. Kelleher and M. Lonnerdal, 2005. Maternal zinc deficiency reduces NMDA recertor expression in neonatal rat brain, which persists into early, adulthood. J. Nerochem., 94: 510-519.
CrossRefDirect Link - Christine, C.W. and D.W. Choi, 1990. Effects of zinc on NMDA receptor mediated channel currents in cortical neurons. J. Neuro. Sci., 10: 108-116.
Direct Link - Cole, T.B., A. Martyanova and R.D. Palmiter, 2001. Removing zinc from synaptic vesicles dose not impair spation learning, memory, or sensorimotor function in the mouse. J. Brain. Res., 891: 253-265.
Direct Link - De oliveria, F.S., M.R. Viana and A.R. Antoniolli et al., 2001. Differential effects of lead and zinc on inhibitory avoidance learning in mice. J. Biol. Med. Behav., 34: 117-120.
PubMedDirect Link - Filinn, J.M., D. Hunter and D.H. Kinkous et al., 2005. Enhanced zinc consumption causes memory deficits and increased brain levels of zinc. J. Phis. Behav., 83: 793-803.
CrossRefDirect Link - Flecknell, P.A., 1996. Laboratory Animal Anesthesia. 2nd Edn., Academic Press Inc., USA., ISBN: 978-0122603617.
Direct Link - Herin, A.G. and E. Aizenman, 2004. Amino terminal domain regulation of NMDA receptor function. J. Phrmacol., 500: 101-111.
CrossRefPubMedDirect Link - Homayovn, H., M.R. Stefani and B.W. Adams et al., 2004. Functional interaction between NMDA and mGlu5 receptors: Effects on working memory instrumental learning, motor behaviors and dopamine release. Neuro. Psychol. Pharmacol. J., 29: 1259-1269.
Direct Link - Jafari-Sabet, M., 2006. NMDA receptor blocker prevents the facilitatory effects of post-trainig intra-dorsal hippocamal NMDA and physostigmin on memory retention of passive avoidance learning. J. Behav. Brain. Res., 169: 120-127.
CrossRefDirect Link - Kawabe, K., T. Iwasaki and Y. Ichitani, 2007. Repeated treatment with N-methyl D-aspartate antagonists in neonatal, but not adult, rats causes Long- term deficits of radial-arm maze learning. J. Brain. Res., 1169: 77-86.
CrossRef - Keller, K.A., A. Grider and J.A. Coffield, 2001. Age dependent influence of dietary zinc restriction on short term memory in male rats. J. Physiol. Behav., 72: 339-348.
CrossRefDirect Link - Xu, L.S., L.X. Yang, W.W. Hu, X. Yu, L. Ma, L.Y. Liu, E.Q. Wei and Z. Chen, 2005. Histamine ameliorates spatial memory deficits induced by MK-801 infusion into ventral hippocampus as evaluated by radial maze taske in rats. J. Pharmacol. Sci., 26: 1448-1448.
CrossRefDirect Link - Maureen, M., S. Sunil and K. Sonu et al., 2004. Cognative and motor development among small-for-gestational-age infants: Impact of zinc supplementation, birth weight and caregiving practices. J. Pediatr., 113: 1297-1304.
Direct Link - Mocchegiani, E., C.B. Freddari and F. Marcellini et al., 2005. Brain aging and neurodegeneration: Role of zinc ion availability. J. Neuro. Biol., 75: 367-390.
Direct Link - Molnar, P. and J.V. Nader, 2001. Synaptically released zinc inhibits N-methyl D-aspartate receptor activation at recurrent mossy fiber synapses. J. Brain. Res., 910: 205-207.
CrossRefDirect Link - Noêmia, M.M.DL., D.C. Laranja, E. Bromberg, R. Roesler and N. Schröder, 2005. Pre-or post-training administration of NMDA receptor blocker MK-801 impairs object recognition memory in rats. J. Behav. Brain. Res., 156: 139-143.
CrossRefDirect Link - Riedel, G., B. Platt and J. Micheau, 2003. Glutamate receptor functions in learning and memory. J. Behav. Brain. Res., 140: 1-47.
PubMedDirect Link - Takeda, A., S. Takefuta and S. Takefuta et al., 2000. Relationship between brain zinc and transient learning impairment of adult rats fed zinc-deficiencent dite. J. Brain. Res., 859: 352-357.
CrossRefDirect Link - Turner, T.Y. and M. Soliman, 2000. Effects of zinc on spatial reference memory and brain dopamine (D1) receptor binding kinetics in rats. J. Nero. Psychol. Pharma., 24: 1203-1217.
CrossRefDirect Link - Vogt, K., J. Mellor and G. Tong et al., 2004. The action of synaptically released zinc at hippocampal mossy fiber synapses. J. Neur., 26: 187-196.
PubMedDirect Link