
B. Prapagdee
Laboratory of Environmental Biotechnology, Faculty of Environment and Resource Studies, Mahidol University, Salaya, Nakhonpathom 73170, Thailand
U. Akrapikulchart
Laboratory of Environmental Biotechnology, Faculty of Environment and Resource Studies, Mahidol University, Salaya, Nakhonpathom 73170, Thailand
S. Mongkolsuk
Department of Biotechnology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1187-1192







ABSTRACT
The aims of this study are to isolate the antagonist from rhizosphere soil and evaluate its potential for biocontrol of anthracnose disease in orchid. The potential microbial antagonist, designated SRA14, was isolate and identified as Streptomyces hygroscopicus. Cell-free culture filtrates of Streptomyces hygroscopicus SRA14 inhibited the growth of Colletotrichum gloeosporioides. The percentage of growth inhibition by the stationary culture filtrate was significantly higher than that of exponential-culture filtrate. Additionally, morphological changes such as hyphal swelling and abnormal shapes were observed in fungi grown on potato dextrose agar that contained the culture filtrates. Application of culture filtrates was able to prevent the expression of anthracnose disease on orchid, indicating that disease inhibition was due to antifungal metabolites in the culture filtrates. No significantly the prevention of anthracnose development was observed in the stationary culture filtrate and mancozeb-treated leaves. Present data suggested the stationary culture filtrate of SRA14 can be used as biofungicide for control of anthracnose diseases in Dendrobium orchid.
PDF Abstract XML References Citation
How to cite this article
B. Prapagdee, U. Akrapikulchart and S. Mongkolsuk, 2008. Potential of a Soil-Borne Streptomyces hygroscopicus for Biocontrol of Anthracnose Disease Caused by Colletotrichum gloeosporioides in Orchid. Journal of Biological Sciences, 8: 1187-1192.
DOI: 10.3923/jbs.2008.1187.1192
URL: https://scialert.net/abstract/?doi=jbs.2008.1187.1192
DOI: 10.3923/jbs.2008.1187.1192
URL: https://scialert.net/abstract/?doi=jbs.2008.1187.1192
INTRODUCTION
Dendrobium orchid is a one of the important economical cut-flower plants in many countries. However, orchid cultivation has faced a lot of problems, particularly fungal diseases. Colletotrichum gloeosporioides (Penz.) causes anthracnose disease in a wide variety of agricultural crops in worldwide (Bautista-Banos et al., 2003; Gamagae et al., 2003). Chemical fungicides are extensively used in current agriculture. However, excessive use of chemical fungicides in agriculture has led to deteriorating human health, environmental pollution and development of pathogen resistance to fungicide. Microbial antagonists are widely used for the biocontrol of fungal plant diseases due to lack of induction of pathogen resistance and reduction of chemical fungicide residues in the environment. Antagonism may be accomplished by competition, parasitism, antibiosis, lysis or by a combination of these modes of action (Chernin and Chet, 2002).
Many species of Streptomyces, are well known as antifungal biocontrol agents that inhibit several plant pathogenic fungi e.g., Gauemannomyces graminis var. tritici (Coombs et al., 2004), Phytophtora capsici (Joo, 2005), Fusarium oxysporum f. sp. cubense (Cao et al., 2005), Sclerotium rolfsii (Errakhi et al., 2007). The antagonistic potentials of Streptomyces to pathogenic fungi involved the production of extracellular hydrolytic enzymes and moreover, secondary antifungal compounds were also reported by Trejo-Estrada et al. (1998a), El-Tarabily et al. (2000) and Ouhdouch et al. (2001). However, data related to the potential of the extracellular metabolites of Streptomyces strains to control the anthracnose disease caused by C. gloeosporioides are limited. In this study, we report the powerful Streptomyces strain that is able to control the anthracnose disease caused by C. gloeosporioides in Dendrobium orchid. The antifungal metabolites produced by this antagonist could be applied for theirs future use as a biofungicide for the control of anthracnose disease in several agricultural plants.
Antifungal activity of cell-free culture filtrates produced by a potent antagonistic streptomyces against C. gloeosporioides: Twenty-four random rhizosphere soil samples were collected from paddy fields and orchards in the central part of Thailand. The soil suspension was spread onto starch-casein-agar plates and incubated at 28°C for 12-14 days. Colonies of Streptomyces on the agar plates were picked on the basis of their morphological characteristics and purified on ISP-2 agar (Shirling and Gottlieb, 1966). All the isolates were screened for their in vitro antagonism against C. gloeosporioides by dual culture assay, according to the modified method of Crawford et al. (1993).
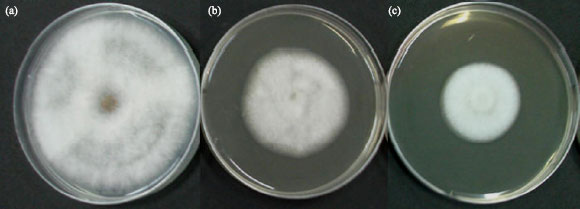 | |
| Fig. 1: | Antifungal activity of cell-free culture filtrates of the SRA14 against C. gloeosporioides on PDA plates. In vitro antagonism of the strain SRA14 to C. gloeosporioides was evaluated on PDA plates. The strain SRA14 was grown on PDA plate amended with either 20% (v/v) exponential (b) or stationary (c) culture filtrates. Control culture of C. gloeosporioides was cultivated on culture filtrate-free PDA (a). Growth inhibitory activity was observed after 6 days of incubation in comparison to the controls |
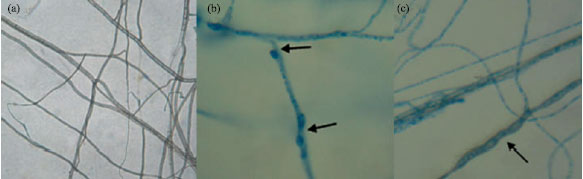 | |
| Fig. 2: | Effects of cell-free culture filtrates on the hyphal morphology of C. gloeosporioides. Hyphal morphology of 3 day-old C. gloeosporioides grown on PDA amended with 20% (v/v) exponential (b) and stationary (c) culture filtrates was observed under light microscopes (40X) compared with the fungal hyphal cultured on PDA plate (a) |
The SRA14 had a strong antagonistic activity to C. gloeosporioides. Analysis of the 16S rDNA gene sequences showed that the SRA14 is closely related to Streptomyces hygroscopicus (98% similarity) with GenBank database accession number AB184760. It has been reported that Streptomyces violaceusniger G10 showed a strong antagonism toward F. oxysporum f. sp. cubense by producing extracellular antifungal metabolites (Getha and Vikineswary, 2002). Fermentation broth of the SRA14 cultured in ISP-2 broth (Shirling and Gottlieb, 1966) was collected at 1 and 3 days to obtain the exponential phase and stationary phase cultures, respectively. The cell-free supernatant was filtered aseptically through a sterile membrane with 0.45 μm pore size. Culture supernatant was added to Potato Dextrose Agar (PDA) to yield a final concentration of 20% (v/v). The radial growth of C. gloeosporioides was inhibited by both the 20% (v/v) exponential (Fig. 1b) and stationary (Fig. 1c) culture filtrates in comparison with the control culture (Fig. 1a). The results implied that the culture filtrates of the SRA14 contain the antifungal metabolite(s).
The finding showed that extracellular metabolites in the culture filtrates of the SRA14 inhibited the growth of pathogenic fungi prompted the investigation of the effects of culture filtrates on the fungal hyphal structure. Light microscope investigation revealed that growth inhibition of C. gloeosporioides as a response to the exponential (Fig. 2b) and stationary (Fig. 2c) culture filtrates was accompanied by marked cellular changes including hyphal swelling, distortion and bulbous roundedness on hyphal structure. None of these abnormal shapes were observed in the control experiments (Fig. 2a). Abnormal hyphal structures of the inhibited fungal hyphae resulting from diffusible antifungal secondary compounds have been previously reported by Getha and Vikineswary (2002) and Taechowisan et al. (2005). In addition, the antifungal mechanism of antagonists has been attributed to the action of hydrolytic enzymes such as chitinase, β-1,3-glucanase and protease (De Boer et al., 1998; Wang et al., 1999, 2002).
Production of extracellular chitinase enzyme involved in the antifungal potential of S. hygroscopicus SRA14: As stated in many previous reports, the production of chitinase enzyme by Streptomyces was related to fungal growth inhibition and the biological control of fungal pathogens was possible because of the ability of Streptomyces to degrade fungal cell walls (Mahadevan and Crawford, 1997, 1999; El-Tarabily et al., 2000; Mukherjee and Sen, 2006). The SRA14 was grown in ISP-2 broth and collected cell-free supernatants for chitinase determination. The estimation of chitinase activity was carried out by the procedure described by Wang et al. (2002) using colloidal chitin as substrate. The SRA14 produced relatively high levels of chitinase (5.2 U mg-1 protein) at day 1 of the incubation period. The level of chitinase activity was sharply increased during the exponential phase and dramatically declined when the cells entered the stationary phase (Data not shown). According to Joo (2005), Streptomyces halstedii AJ-7 produces extracellular chitinase and causes abnormal hyphal morphology. The culture filtrate of Pseudomonas aeruginosa K-187 causes growth aberration, hyphal swelling and lysis of many fungi due to its high content of chitinase enzyme (Wang et al., 1999). The antifungal potential of the exponential culture filtrate was probably related to the increased production of chitinase enzyme during the exponential phase. It should be noted that one of the possible antifungal mechanisms of the SRA14 may be associated with the production of extracellular chitinase enzyme.
Stationary culture filtrate possessed a higher antifungal potential than the exponential culture filtrate: Growth inhibitory experiment was then performed to find out whether exponential or stationary culture filtrates play the major role in the fungal growth inhibition. Culture supernatants from both the growth phases of the SRA14 were collected and aseptically added to Potato Dextrose Broth (PDB) to yield a final concentration of 20% (v/v). Spore suspension (104 spores mL-1) of C. gloeosporioides was inoculated into PDB medium incubated on incubator shaker at 120 rpm, 28°C for 1, 2, 3, 5 and 7 day, respectively. The fungal growth monitored by dry weight determination.
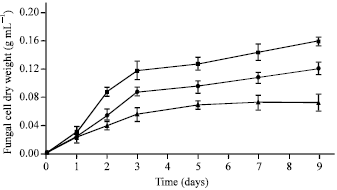 | |
| Fig. 3: | Antifungal effects of cell-free culture filtrates obtained from the strain SRA14 as observed by the submerged growth inhibition of C. gloeosporioides. C. gloeosporioides was grown on PDB, amended with either 20% (v/v) exponential (●) or stationary (▲) culture filtrates. Control culture of C. gloeosporioides (■) was cultivated on culture filtrate-free PDB. Mycelial growth of the fungus was monitored by measuring the cell dry weight at 1 day intervals. The experiment was independently repeated at least three times and representative data are shown |
The result found that the submerged growth of C. gloeosporioides was inhibited by both the 20% (v/v) exponential and stationary culture filtrates in comparison with the control culture (Fig. 3). These results indicated that fungal growth suppression resulted from the presence of extracellular antifungal metabolites in culture filtrates. The percentages of the submerged growth inhibition of C. gloeosporioides by the exponential and stationary culture filtrates on the day 3 of incubation were 26.58 and 53.77%, respectively. El-Abyad et al. (1993) reported that the culture filtrate of either Streptomyces pulcher or Streptomyces canescens at a concentration of 80% (v/v) inhibited the mycelial growth of F. oxysporum f. sp. lycopersici, Verticillium albo-atrurn and Alternaria solani.
The percentage of growth inhibition by the stationary culture filtrate was significantly higher than that of exponential culture filtrate. These findings imply that the antifungal potential of the culture filtrates is not solely due to the increased chitinase production, as shown by the increase in percentage of fungal growth inhibition by the stationary culture filtrate. There is a possibility that the increased antifungal activity against C. gloeosporioides by the stationary culture filtrate of the SRA14 is a consequence of the production of extracellular secondary antifungal compound(s). The production of secondary antifungal compound(s) as antibiotics has been already reported in many species of Streptomyces (Xiao et al., 2002; Fguira et al., 2005; Taechowisan et al., 2005).
 | |
| Fig. 4: | Biocontrol of anthracnose disease on Dendrobium leaves by cell-free culture filtrates of the SRA14. Orchid leaves were treated with different treatments including, (T1) no treatment as negative control, (T2) C. gloeosporioides as positive control, (T3) 200 μL of 1 mg mL-1 of mancozeb as chemical fungicide control, (T4) 200 μL of the exponential culture filtrates, (T5) 200 μL of the stationary culture filtrates and (T6) 200 μL of overnight culture (105 spores mL-1) of the SRA14. The 100 μL of spore suspension (104 spores mL-1) of C. gloeosporioides was placed on abaxial surface of each orchid leaf in all treatments, except T1. The visible symptom appearance of all orchid leaves was observed for 5 days after fungal infection |
The degree of fungal growth inhibition corresponded to the increase in concentration of the culture filtrate (data not shown). The growth inhibitory effects of culture filtrates decreased with the extension of fungal incubation period. Other previous studies reported that the decreases in the degree of growth inhibition were associated with the increases in the incubation period of the fungal culture (El-Abyad et al., 1993; Chang et al., 2007).
Preventive application of culture filtrates from antagonistic rhizobacteria on anthracnose disease expression in Dendrobium leaves: According to the in vitro antagonism against C. gloeosporioides, S. hygroscopicus SRA14 was evaluated for in vivo biocontrol of anthracnose disease in 1 year-old Dendrobium plants. The experiment used a Completely Randomized Design (CRD) which was divided into 6 treatments with 5 replications. The abaxial surface of orchid leaf was scratched with a sterile needle to make a little wound. Then, 100 μL of spore suspension (104 spores mL-1) of C. gloeosporioides was applied on abaxial surface of each orchid leaf in all treatments, except negative control treatment (T1). The sizes of visible lesion on orchid leaves were measured for 5 days after fungal infection. The results in greenhouse study showed that the Disease Severity Index (DSI) was significantly (p≤0.05) reduced on orchid leaflets treated with the exponential (T4), stationary (T5) culture filtrates and cell culture of the SRA14 (T6) compared with the absence of the biocontrol agent (T2) (Fig. 4). Application of the culture filtrates of the SRA14 inhibited anthracnose disease on orchid leaves, indicating that suppression was due to antifungal compounds in the culture filtrate. The stationary culture filtrate was more suppress the symptom expression than that of exponential culture filtrate. There was no significantly different in the control of anthracnose development on orchid leaves between the stationary culture filtrate and chemical fungicide treatment (T3). The results indicated that the stationary culture filtrate controlled anthracnose disease as effectively as the chemical control. Several studies have demonstrated that many of the secondary compounds produced by Streptomyces strains have a broad spectrum activity against plant fungal pathogens (Trejo-Estrada et al., 1998; Fguira et al., 2005; Taechowisan et al., 2005). Antibiotic production by S. hygroscopicus can inhibit a broad range of fungal pathogens e.g., Rhizoctonia solani, Pythium ultimum, F. oxysporum and Sclerotinia homeocarpa (Rothrock and Gottlieb, 1984; Chamberlain and Crawford, 1999). In conclusion, there is good potential for using the stationary culture filtrate of S. hygroscopicus SAR14 to as biofungicide to control the anthracnose disease in orchid cultivation.
ACKNOWLEDGMENTS
The authors thank K. Duangmal, C. Kuekulvong and W. Tanboon for their technical assistance. This research was supported by the Grant (MRG4980130) from Thailand Research Fund (TRF) to B. Prapagdee.
REFERENCES
- Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Cao, L., Z. Qiu, J. You, H. Tan and S. Zhou, 2005. Isolation and characterization of endophytic streptomycete antagonists of fusarium wilt pathogen from surface-sterilized banana roots. FEMS Microbiol. Lett., 247: 147-152.
CrossRefDirect Link - Chamberlain, K. and D.L. Crawford, 1999. In vitro and in vivo antagonism of pathogenic turfgrass fungi by Streptomyces hygroscopicus strains YCED9 and WYE53. J. Ind. Microbiol. Biotechnol., 23: 641-646.
CrossRefDirect Link - Chang, W.T., Y.C. Chen and C.L. Jao, 2007. Antifungal activity and enhancement of plant growth by Bacillus cereus grown on shellfish chitin wastes. Bioresour. Technol., 98: 1224-1230.
CrossRefDirect Link - Coombs, J.T., P.P. Michelsen and C.M.M. Franco, 2004. Evaluation of endophytic actinobacteria as antagonists of Gaeumannomyces graminis var. tritici in wheat. Biol. Control, 29: 359-366.
CrossRef - Crawford, D.L., J.M. Lynch, J.M. Whipps and M.A. Ousley, 1993. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Applied Environ. Microbiol., 59: 3899-3905.
Direct Link - De-Boer, W., P.J.A.K. Gunnewiek, P. Lafeber, J.D. Janse and B.E. Spit et al., 1998. Antifungal properties of chitinolytic dune soil bacteria. Soil Biol. Biochem., 30: 193-203.
CrossRefDirect Link - El-Abyad, M.S., M.A. El-Sayad, A.R. El-Shanshoury and S.M. El-Sabbagh, 1993. Towards the biological control of fungal and bacterial diseases of tomato using antagonistic Streptomyces sp. Plant Soil., 149: 185-195.
CrossRef - El-Tarabily, K.A., M.H. Soliman, A.H. Nassar, H.A. Al-Hassani, K. Sivasithamparam, F. McKenna and G.E. Hardy, 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol., 49: 573-583.
Direct Link - Errakhi, R., F. Bouteau, A. Lebrihi and M. Barakate, 2007. Evidences of biological control capacities of Streptomyces sp. against Sclerotium rolfsii responsible for damping-off disease in sugar beet (Beta vulgaris L.). World J. Microbiol. Biotechnol., 23: 1503-1509.
CrossRefDirect Link - Fguira, L.F., S. Fotso, R.B. Ameur-Mehdi, L. Mellouli and H. Laatsch, 2005. Purification and structure elucidation of antifungal and antibacterial activities of newly isolated Streptomyces sp. strain US80. Res. Microbiol., 156: 341-347.
CrossRefDirect Link - Gamagae, S.U., D. Sivakumar, R.S.W. Wijeratnam and R.L.C. Wijesundera, 2003. Use of sodium bicarbonate and Candida oleophila to control anthracnose in papaya during storage. Crop Prot., 22: 775-779.
CrossRefDirect Link - Getha, K. and S. Vikineswary, 2002. Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4: Indirect evidence for the role of antibiosis in the antagonistic process. J. Ind. Microbiol. Biotechnol., 28: 303-310.
CrossRefDirect Link - Joo, G.J., 2005. Production of an anti-fungal substance for biological control of Phytophthora capsiciI causing phytophthora blight in red-peppers by Streptomyces halstedii. Biotechnol. Lett., 27: 201-205.
CrossRefDirect Link - Mahadevan, B. and D.L. Crawford, 1997. Properties of the chitinase of the antifungal biocontrol agent Streptomyces lydicus WYEC108. Enzyme Microb. Technol., 20: 489-493.
CrossRefDirect Link - Ouhdouch, Y., M. Barakate and C. Finance, 2001. Actinomycetes of Moroccan habitats: Isolation and screening for antifungal activities. Eur. J. Soil Biol., 37: 69-74.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Taechowisan, T., C. Lu, Y. Shen and S. Lumyong, 2005. Secondary metabolites from endophytic Streptomyces aureofaciens CMUAc130 and their antifungal activity. Microbiology, 151: 1691-1695.
CrossRefDirect Link - Trejo-Estrada, S.R., A. Paszczynski and D.L. Crawford, 1998. Antibiotics and enzymes produced by the biocontrol agent Streptomyces violaceusniger YCED-9. J. Ind. Microbiol. Biotechnol., 21: 81-90.
CrossRefDirect Link - Wang, S.L., W.J. Hsiao and W.T. Chang, 2002. Purification and characterization of an antimicrobial chitinase extracellularly produced by Monascus purpureus CCRC31499 in a shrimp and crab shell powder medium. J. Agric. Food Chem., 50: 2249-2255.
CrossRefDirect Link - Wang, S.L., T.C. Yieh and I.L. Shih, 1999. Production of antifungal compounds by Pseudomonas aeruginosa K-187 using shrimp and crab shell powder as a carbon source. Enzyme Microb. Technol., 25: 142-148.
CrossRef - Xiao, K., L.L. Kinkel and D.A. Samac, 2002. Biological control of Phytophthora root rots on alfalfa and soybean with Streptomyces. Biol. Control, 23: 285-295.
CrossRefDirect Link - Mukherjee, G. and S.K. Sen, 2006. Purification, characterization and antifungal activity of chitinase from Streptomyces venezuelae P10. Curr. Microbiol., 53: 265-269.
CrossRef