
P. Ghadam
Department of Biology, Alzahra University, Tehran, Iran
A. Abadi
Department of Health and Social Medicine, Shaheed Beheshti University of Medical Sciences, Tehran, Iran
A. Asadzadeh
Department of Biology, Alzahra University, Tehran, Iran
N. Safari
Department of Biology, Alzahra University, Tehran, Iran
A. Shabani
Department of Biology, Alzahra University, Tehran, Iran
M. Sharifian
Department of Biology, Alzahra University, Tehran, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 6 | Page No.: 1100-1103







ABSTRACT
In this study, we found that allelic polymorphism in Hp gene acts as a major determinant of susceptibility for the development of diabetic cardiovascular complications. We studied this gene in 122 Iranian diabetic cardiovascular patients. The results showed that distribution of the Hp phenotypes were found to be in Hardy-Weinberg equilibrium. By using the chi square test we determined the association between Hp allele and diabetic cardiovascular diseases (χ2 = 52.98 p<0.001) and this finding was independent of gender (χ2 = 0.39 p>0.05). The chance of having allele two in diabetic patients with cardiovascular disease was 22.31 (95% CI, 6.18 to 80.5) times more than those with allele one (p<0.0001).
PDF Abstract XML References Citation
How to cite this article
P. Ghadam, A. Abadi, A. Asadzadeh, N. Safari, A. Shabani and M. Sharifian, 2008. The Relationship of Haptoglobin Polymorphism and Cardiovascular Diseases in Some of Iranian Diabetic Patients. Journal of Biological Sciences, 8: 1100-1103.
DOI: 10.3923/jbs.2008.1100.1103
URL: https://scialert.net/abstract/?doi=jbs.2008.1100.1103
DOI: 10.3923/jbs.2008.1100.1103
URL: https://scialert.net/abstract/?doi=jbs.2008.1100.1103
INTRODUCTION
Haptoglobin (Hp) is a hemoglobin–binding polymorphic plasma glycoprotein present in all vertebrates. There are two classes of alleles for haptoglobin, denoted class one and class two and three potential genotypes denoted Hp 1-1, Hp 2-1 and Hp 2-2 (Quaye, 2008). Hp consists of two different polypeptide chains, α-chain and β-chain. β-chain is heavier than α-chain and is identical in all Hp phenotypes (Langlois and Delanghe, 1996).
Oxidation of low-density lipoprotein is catalyzed by heme and leads to vascular endothelial cell damage and atherosclerosis (Vercellotti et al., 1994). Haptoglobin by binding to hemoglobin and removing it from the circulation prevents iron –stimulated formation of oxygen radicals and has an important role as an antioxidant (Quaye, 2008). The antioxidant capacity of Hp 2-2 in circulation is lower than that of Hp 1-1 because Hp 2-2 binds to hemoglobin with lower binding affinity than Hp1-1 (Lange, 1992).
In some races individuals who were homozygous for Hp allele two (Hp 2-2) were found to be at significantly greater risk of developing myocardial infraction, stroke and cardiovascular death (UK Prospective Diabetes Study Group, 1998; Carter and Worwood, 2007 ). In this study, we found the relationship between Hp phenotypes and the risk of cardiovascular diseases (CVD) in some Iranian diabetic patients.
MATERIALS AND METHODS
In 2007, 122 consecutive patients (61 men, 61 women) who presented with diabetic cardiovascular disease were enrolled in this study. These patients were recruited from Alghadir Hospital (Tehran-Iran) for participation in this trial. In control group (50 cardiovascular patients) Hp genotyping was determined.
Blood samples were collected in the presence of EDTA and the plasma was stored at -70 °C. Genomic DNA was extracted from whole blood using DNGTM plus solution (Fermentas). Two sets of polymerase chain reaction (PCR) were performed (Koch et al., 2002):
| • | With primers A (5`- GAGGGGAGCTTGCC TTTCCATTG- 3`) (forward) and B (5-` GAGATTTTTG AGCCCTGGCTGGT-3`) (reverse) for the amplification of a 1757-bp Hp allele one -specific sequence and a 3481-bp Hp allele two -specific sequence (Annealing temperature 63 °C, cycle 30) |
| • | Primers C (5`-CCTGCCTCGTATTAACTGCACCAT-3`) (forward) and D (5` CCGAGTGCTCCA CATAGCCATGT3`) (reverse) amplified a 349-bp fragment that is generated only in the presence of Hp allele two template (Annealing temperature 61 °C, cycle 30) |
Statistical analysis: Chi-square test was used to determine association between Hp allele and diabetic CVD and CVD diseases and gender effect on distribution Hp genotypes in those patients. Odds ratio were calculated together with their 95% confidence intervals. The proportion test was used to compare percentage of two genotypes in diabetic cardiovascular disease. For all statistical analyses, p<0.05 was considered significant (the calculations were manual).
RESULTS
After electrophoresis of the reaction products in 1% agarose gel, Hp genotyping-specific banding patterns were obtained.With primers A and B, Hp 1-1 and Hp 2-2 genotypes were characterized by single bands representing the 1757 and 3481 bp products, respectively and Hp 2-1 genotype was characterized by the presence of both the 1757 and 3481 bp products (Fig. 1). Primers C and D were used for detecting the 349 bp Hp allele two-specific product (Fig. 2).
The distribution of three major Hp genotypes in diabetic cardiovascular patients was 1.6% (1-1), 52% (2-1), 45% (2-2) in the men and 3.27% (1-1), 54.09% (2-1), 42.62% (2-2) in the women. These prevalence were shown to be in Hardy-Weinberg equilibrium. In the no diabetic Iranian cardiovascular patients these prevalence were respectively 36% (1-1), 62% (2-1), 2% (2-2) (data not shown).
The Chi-square test determined the association between Hp allele and diabetic cardiovascular diseases (χ2 = 52.98 p<0.001) and this finding was independent of gender (χ2 = 0.39 p>0.05). Odds ratio shows that the chance of having Hp 2-2 phenotype in diabetic patients (men) with CVD were estimated about 504 (95% CI, 29.6 to 8577.7) times more than those with Hp 1-1 phenotype ( p<0.0001) and 27.12 (95% CI, 3.47 to 211.73) times more than those with Hp 2-1 phenotype (p = 0.0008). The chance of having Hp 2-1 phenotype in the same patients were calculated about 18.58 (95% CI, 2.33 to 147.74) times more than those with Hp 1-1 phenotype (p = 0.02). The chance of having Hp 2-2 phenotype in diabetic patients (women) with CVD were 234 (95% CI, 19.7 to 2779) times more than those with Hp 1-1 phenotype (p<0.0001) and 24.42 (95% CI, 3.12 to 190) times more than those with Hp 2-1 phenotype (p = 0.001). The chance of having Hp 2-1 phenotype in the same patients were calculated about 9.58 (95% CI, 2.05 to 44.7) times more than those with Hp 1-1 phenotype (p = 0.0019). The alterations of odds ratio in both sex were homogenous (Table 1).
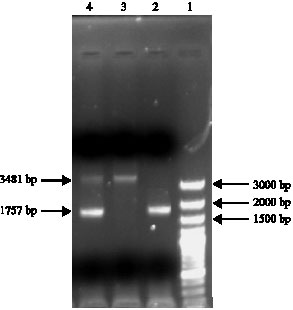 | |
| Fig. 1: | Haptoglobin genotyping with primer A and B. (Lane 1) DNA size marker (Lane 2) genotype Hp 1-1 (Lane 3) genotype Hp 2-2 (Lane 4) genotype Hp 2-1 |
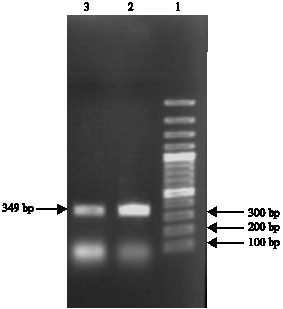 | |
| Fig. 2: | Haptoglobin genotyping with primer C and D. (Lane 1) DNA size marker, (Lanes 2 and 3) Hp 2-specific amplification product |
| Table 1: | The relationship between diabetic cardiovascular disorder and Hp genotypes |
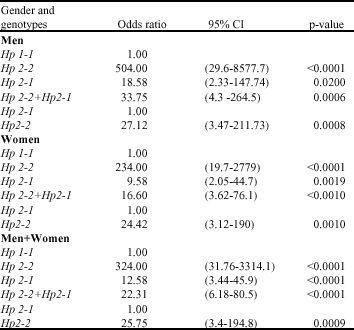 | |
It is concluded that there is no difference between percentage of Hp 2-2 and Hp 2-1 phenotype in diabetic patients with cardiovascular diseases (Z = 1, p>0.05).
DISCUSSION
The oxidative modification hypothesis of atherosclerosis states that oxidative modification of low-density lipoprotein (LDL) or other oxidative events within the blood vessel wall promotes the development and progression of atherosclerotic lesion (Brown and Goldstein, 1983; Steinberg and Witzum, 2002). Hemoglobin (Hb) is an oxidant of LDL by the Fenton reaction (Kristiansen et al., 2001; Mowri et al., 2000). Hb is released from red blood cells with intravascular hemolysis in endothelial cells and injury may enter it into the sub endothelial space (Fernandez et al., 2001).
The amount of hemoglobin penetrating into the blood vessel wall is increased in diabetes because of an increased turnover of red blood cells (Venerando et al., 2002) and an increase in endothelial cell dysfunction and injury (Laight et al., 2000).
So the generation of reactive oxygen species has an important role in the development of diabetic vascular complication (Melamed-Frank et al., 2001; Suzuki et al., 1999). Haptoglobin is an antioxidant that is able to prevent hemoglobin-driven oxidation (Langlois and Delanghe, 1996; Melamed-Frank et al., 2001). The mechanism by which haptoglobin serves as an antioxidant is believed to be via stabilization of the heme moiety within the hemoglobin protein (Quaye, 2008). Ability of haptoglobin to protect against hemoglobin-driven oxidative stress is lost when hemoglobin becomes heavily glycosylated. Glycosylated Hb is markedly increased in the diabetic state (Asleh et al., 2003). In diabetic white, South Asian and Afro-Caribbean patients have been shown that Hp 2-2 increases the risk of developing CVD (UK Prospective Diabetes Study Group, 1998; Carter and Worwood, 2007).
This study for the first time in Iran suggests that Hp genotype may contribute to developing CVD in diabetic patients and we showed that the presence of Hp allele two in diabetic individuals increases the risk of developing CVD.
ACKNOWLEDGMENTS
The authors wish to thank Alghadir Hospital and Alzahra University for their grateful support of this work.
REFERENCES
- Asleh, R., S. Marsh, M. Shilkrut, O. Binah and J. Guetta et al., 2003. Genetically determined heterogeneity in hemoglobin scavenging and susceptibility to diabetic cardiovascular disease. Circ. Res., 92: 1193-1200.
PubMed - Brown, M.S. and J.L. Goldstein, 1983. Lipoprotein metabolism in the macrophage: Implications for cholesterol deposition in atherosclerosis. Annu. Rev. Biochem., 52: 223-261.
PubMed - Carter, K. and M. Worwood, 2007. Haptoglobin: A review of the major allele frequencies worldwide and their association with diseases. Int. J. Lab. Hematol., 29: 92-110.
CrossRefPubMedDirect Link - Fernandez, A.Z., F. Lopez, A. Tablante, E. Romano and E. Hurt-Camejo et al., 2001. Intravascular hemolysis increases atherogenicity of diet induced hypercholesterolemia in rabbits in spite of heme-oxygenase-1 gene and protein induction. Atherosclerosis, 158: 103-111.
PubMed - Koch, W., W. Latz, M. Eichinger, A. Roguin, A.P. Levy, A. Schömig and A. Kastrati, 2002. Genotyping of the common haptoglobin Hp1/2 polymorphism based on PCR. Clin. Chem., 48: 1377-1382.
PubMed - Kristiansen, M., J.H. Graversen, C. Jacobsen, O. Sonne and H.J. Hoffman et al., 2001. Identification of the hemoglobin scavenger receptor. Nature, 409: 198-201.
PubMed - Laight, D.W., M.J. Carrier and E.E. Anggard, 2000. Antioxidants, diabetes and endothelial dysfunction. Cardiovasc. Res., 47: 457-464.
PubMed - Lange, V., 1992. Haptoglobin polymorphism: Not only a genetic marker. Anthropol Anz., 50: 281-302.
PubMed - Langlois, M.R. and J.R. Delanghe, 1996. Biological and clinical significance of haptoglobin polymorphism in humans. Clin. Chem., 42: 1589-1600.
PubMedDirect Link - Melamed-Frank, M., O. Lache, B.I. Enav, T. Szafranek and N.S. Levy et al., 2001. Structure-function analysis of the antioxidant properties of haptoglobin. Blood, 98: 3693-3698.
PubMed - Mowri, H.O., B. Frei and J.F. Keaney, 2000. Glucose enhancement of LDL oxidation is strictly metal ion dependent. Free Radic. Biol. Med., 29: 814-824.
PubMed - Quaye, I.K., 2008. Haptoglobin, inflammation and disease. Trans. R. Soc. Trop. Med. Hyg., 102: 735-742.
CrossRefPubMedDirect Link - Suzuki, D., T. Miyata, N. Saotome, K. Horie and R. Inagi et al., 1999. Immunohistochemical evidence for an increased oxidative stress and carbonyl modification of proteins in diabetic glomerular lesions. J. Am. Soc. Nephrol., 10: 822-832.
PubMedDirect Link - UK Prospective Diabetes Study Group, 1998. Ethnicity and cardiovascular disease: The incidence of myocardial infraction in white, South Asian and Afro-Caribbean patients type 2 diabetes. Diabetes Care, 21: 1271-1277.
PubMed - Venerando, B., A. Fiorilli, G. Croci, C. Tringali and G. Goi et al., 2002. Acidic and netural sialidase in the erythrocyte membrane of type 2 diabetic patients. Blood, 99: 1064-1070.
PubMed - Vercellotti, G.M., G. Balla, J. Balla, K. Nath, J.W. Eaton and H.S. Jacob, 1994. Heme and the vasculature: An oxidative hazard that induces antioxidant defenses in the endotheliume. Artif. Cells Blood Substit. Immobil. Biothecnol., 22: 207-213.
PubMed