
W.M. Muiru
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 29053, Nairobi, Kenya
E.W. Mutitu
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 29053, Nairobi, Kenya
D.M. Mukunya
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 29053, Nairobi, Kenya
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 6 | Page No.: 1021-1026







ABSTRACT
Three antagonistic actinomycetes coded as 14P, 28P and CS35 were isolated from the soil and identified based on morphological, physiological and color formation on Czapeks Dox agar, inorganic salt starch agar and glycerol asparagine agar. The antibiotic metabolites from these isolates were characterized by separation in different solvent systems, performing bioautography, partial purification and freeze drying to enhance their antimicrobial activity. The three isolates showed differences in the carbon utilization tests and isolate coded 14P utilized all the carbon sources tested, isolate 28P utilized all the carbon sources tested except, cellulose, whereas isolate CS35 utilized all the carbon sources except inositol, raffinose and rhamnose. Isolate 28P had rectus type of spore bearing hyphae while isolate 14P formed both rectus and flexuous type whereas isolate CS35 formed only the flexuous type of hyphae. Melanin production was observed only from isolate 14P and CS35 and all the isolates were identified as belonging to the genus Streptomyces. Concentration of the antibiotic metabolites by partial purification and freeze drying resulted in enhanced antimicrobial activity with an overall increase in activity of 21.5 and 20.1%, respectively. The antibiotic metabolites were separated by the various solvent systems with Hexane-Methanol-Water (4:3:3) giving the best separation. The bioautograms revealed the presence of one active compound in isolates CS35 and 14P and two active compounds in isolate 28P. Actinomycetes are a diverse group of organisms and they produce antimicrobial compounds that vary greatly in terms of composition and biological activity.
PDF Abstract XML References Citation
How to cite this article
W.M. Muiru, E.W. Mutitu and D.M. Mukunya, 2008. Identification of Selected Actinomycete Isolates and Characterization of Their Antibiotic Metabolites. Journal of Biological Sciences, 8: 1021-1026.
DOI: 10.3923/jbs.2008.1021.1026
URL: https://scialert.net/abstract/?doi=jbs.2008.1021.1026
DOI: 10.3923/jbs.2008.1021.1026
URL: https://scialert.net/abstract/?doi=jbs.2008.1021.1026
INTRODUCTION
Actinomycetes are a group of organisms that morphologically resemble fungi and physiologically resemble bacteria (Sultan et al., 2002) and they exhibit a very wide range of morphological forms extending from cocci through fragmenting hyphal to permanent and highly differentiated branched mycelium (Sultan et al., 2002; Mustafa et al., 2004; You et al., 2005). Actinomycetes can utilise a variety of organic nutrients but special media are often preferable (Sathi et al., 2001). Their growth is characterized by small compact, soft to leathery colonies tenaciously adhering to the medium, the surface being either flat or elevated (Sathi et al., 2001).
The outer zones of the colonies are smooth but fringes of minute hyphae are observed under the low power microscope. Colonies of some actinomycetes have smooth surfaces while others their surfaces are much folded (Mutitu et al., 2008). The colony surface is usually dry with conical or elevated appearance and it is either free from mycelium or covered with mycelium. The colour of the mycelia can range from nearly colourless to white, chalky red, or grey or olive (Mustafa et al., 2004). The colour is due to the pigment production and examples of pigments produced are phenazines, phenoxazinones and prodiginines (Rahman et al., 2000). Some species forms fairly rings consisting of concentric spore bearing and spore free rings disposed in zones (Sathi et al., 2001).
Many characteristics of actinomycetes have been employed for the purpose of classification and ideally, these should be constant under the same cultural conditions (Sathi et al., 2001). These include shape of the spores, branching manner of the spore chains, nature of the spore chain, form or structure of sporophores (You et al., 2005), cultural characteristics and spore color on various media and carbon utilization (Rahman et al., 2000; Sultan et al., 2002), whole cell sugar and lipid composition (Rahman et al., 2000) and antibiotic production.
Actinomycetes are well known for the production of antibiotics with the group producing over half of the known antibiotics (Hongjuan et al., 2006). Use of antibiotics and other antimicrobial products in plant disease management has been gaining popularity due to the realization of the adverse effect to the human health and environment occasioned by use of the synthetic chemical pesticides (Agrios, 2005). The antagonistic metabolites are normally composed of heterogenous and biologically active compounds (Mustafa et al., 2004). To fully make use of these metabolites, it is necessary to characterize them by separating them in solvent systems, conducting bioassays and ascertaining their stability under different physical and chemical conditions. Enhancement of antimicrobial activity can be achieved through partial purification, freeze drying among other techniques (Mutitu et al., 2008).
Though the actinomycetes are the largest group of actinobacteria with the group producing over 70% of all the naturally occurring antibiotics (Hongjuan et al., 2006), their identification is still short of the universal standard and there is a need to undertake more studies on this. The objectives of this study were to identify actinomycete isolates based on morphology, utilization of various carbon sources, pigment production and characterizing the antimicrobial metabolites through partial purification, freeze drying, chromatographic and bioautographic techniques.
MATERIALS AND METHODS
Identification of actinomycete isolates 14P, 28P and CS35: The experiment was carried out in the Plant Pathology Laboratory in the Department of Plant Science and Crop Protection, University of Nairobi. The actinomycetes were isolated from soils collected at the University of Nairobi and the three isolates were selected after conducting primary screening for the production of antagonistic metabolites using the method by Getha et al. (2004). Identification was done by determining the morphological characteristics, conducting carbon utilization tests and assessment of pigment production.
Determination of morphological characteristics: The media used were (i) Czapeks Dox Agar (NaNO3 2 g, K2HPO4 1 g, MgSO4.7H2O 0.5 g, KCl 0.5 g, FeSO4 0.01 g, Sucrose 30 g, Agar 20 g, distilled water 1 L), (ii) Inorganic salt starch agar (Solution 1). Ten grams of soluble starch was used to make a paste of starch with a small amount (5 mL) of cold distilled water. Using distilled water, this volume was brought to a volume of 500 mL (Solution 2); K2HPO4 (anhydrous basis) 1 g, MgSO4.7H2O 1 g, NaCl 1 g (NH4)2SO4 2 g, CaCO3 2 g, distilled water 500 mL, Trace salts solution (FeSO4.7H2O 0.1 g, MnCl2.4H2O 0.1 g, ZnSO4.7H2O 0.1 g) 1 mL, distilled water 100 mL) pH was adjusted to between 7.0 and 7.4. Starch suspension and salts solution was mixed and 20 g agar was added and liquefied by steaming at 100 °C for 15-20 min. The media was sterilized in conical flasks at 121 °C at one bar pressure for 15 min and (iii) Glycerol asparagine agar [L-asparagine (anhydrous basis) 1 g, Glycerol 10 g, K2HPO4 (anhydrous basis) 1 g, distilled water 1000 mL and trace salts solution 1 mL]. The pH of the solution was adjusted to be between 7.0-7.4. and 20 g of agar was added and liquefied by steaming at 100 °C for 15-20 min. The media was sterilized in the autoclave at 121 °C at one bar pressure for 15 min). The media was cooled to 50 °C and 25 mL of each media was dispensed in sterile 9 cm diameter plates. The media was held for a minimum of 24 h to promote moderate drying and to check for sterility before inoculation.
A turbid suspension of actinomycetes was used to make 5 equally spaced streaks across each plate using a sterile wire loop. Cross hatch streaks were then made on the media and the plates incubated in the dark at 25-28 °C. For each actinomycete isolate, two plates of each media were prepared and observations made after 7, 14 and 21 days. During each observation, determination of the characteristics of the spore bearing hyphae, spore chains and the number of spores at the end of mature hyphae was done by direct microscopic observation of the cross hatch culture surfaces of the opened plates. The sporulating culture surfaces were described in terms of morphological groups of Odakura et al. (1984) and Getha et al. (2004).
Determination of physiological characteristics (carbon utilization tests): The basal medium used was Pridham and Gottlieb medium (Getha et al., 2004) and the carbon sources tested were sucrose, mannitol, I-inositol, cellulose, rhamnose, arabinose and raffinose. D-glucose was used for the positive control and for the negative control, no carbon sources were added. Ten per cent of each carbon source was made by dissolving 1 g of carbon source in 10 mL of sterile distilled water. Apart from I-inositol and cellulose, the other carbon sources were sterilized by filtering using bacteriological filters (0.22 μm). I-inositol and cellulose were sterilized by spreading the sugars in sterile flasks then adding acetone free ethyl agar and then allowing the ether to evaporate overnight at room temperature in a laminar flow hood.
The basal media was autoclaved and cooled to 60 °C and each sterile carbon source was added to give a concentration of approximately 1%. After agitating the mixture, 25 mL of the media were dispensed per plate and two plates of each carbon source for each isolate was prepared. Two drops of washed inoculum (0.05 mL) were placed at opposite sides on each plate. Using a sterile wire loop, 2 parallel streaks were made from these drops of washed inoculum. Plates were incubated at room temperature and observations made for 10-16 days and during each observation, growth on different carbon sources was compared to the positive and the negative control.
Determination of colour or pigment production by the various isolates: For color determination, the media which had been prepared for morphological characterization was used. Munsel color chart was used and color determination was done for mature sporulating aerial surface growth, color of the substrate mycelium as seen from the reverse side of the plate and for diffusible pigments other than melanin. The color of the aerial surface growth was determined as white, gray, yellow, red, blue, green and violet. The presence of other soluble colors other than melanin pigmentation was determined if soluble colors other than brown or black was produced on any medium or if brown was distinctively modified with red, yellow, green, blue or violet. In case where these soluble colors were present, their response to pH change was determined by adding a drop of 0.05 N HCl and 0.05 N NaOH to the colored agar and observation made immediately and after 10-15 min.
Characterization of the antibiotic metabolites: The antibiotic metabolites were produced in shaken liquid media and bioassay conducted using Pythium spp. as the test pathogen following the method by Mondal et al. (2001).
Movement of antibiotic metabolites in specific solvent systems: The movement of the antibiotics in specific solvent systems was used to develop the chromatograms following the method by Odakura et al. (1984). Using a micropipette, 5 μL of the antibiotic filtrates were applied 3 cm from the lower edge of the chromatography paper strips (Whatman No. 1) and then air dried. The chromatography paper strips were immersed to a depth of 1 cm in the solvents and the upper ends attached to the inner surface of the lids using adhesive tapes. The different solvents tested for elution of the antibiotic were methanol, butanol, water, acetic acid and n-hexane in different combinations. Fifty milliliter of each solvent were placed in chromatography tanks and ascending development was allowed without preliminary saturation of the chromatograms with the vapours of the solvents. The ascending development of the chromatograms was stopped when the solvent fronts reached a distance of 15 cm from the origin. The solvent fronts were marked, measured and the paper strips aseptically folded with aluminium foil for use in bioautography work.
Bioautography of the antibiotic metabolites using paper chromatography: The solvent systems which managed to move and separate the crude antibiotics into their different components were selected and used to make solvent combinations that were used to separate the crude antibiotics. The solvent systems used were hexane-methanol-water 4:3:3 (v/v), hexane-butanol-water (65:25:10 v/v), methanol-n-heaxane (60:40 v/v) and butanol-acetic acid-water (50:25:25 v/v). The solvent systems were used to develop ascending chromatograms as explained above. Bioautography set up consisted of a base layer of 20 mL of sterile Czapeks agar in sandwich boxes (14x9 cm) left for a while to set and solidify. Ten milliliter of Czapeks agar seeded with 105 propagules mL-1 of the Pythium sp. was evenly spread on the basal layer.
The seeded layer was allowed to set and then the developed chromatogram strips were placed on the surface of the seeded agar ensuring good contact to allow the antibiotic to diffuse from the paper to the agar to inhibit the growth of the test organism. The lids of the sandwich boxes were replaced aseptically and the boxes were incubated at 24-26 °C for 24-48 h. The presence of inhibition as evidenced by the clear zones around where active components were present was determined and the distance from the point of antibiotic application to the centre of the clear zones were measured and recorded.
Partial purification of the antibiotic metabolites in culture filtrates: For partial purification of the antibiotic metabolites, a modification of the method by Mutitu et al. (2008) was adopted. Charcoal (500 g) was dried and ground coarsely and put in a canister and activated by heating at 200 °C for 1 h in the oven and then cooled to room temperature (22 ± 2 °C). The cell free culture filtrates were mixed with 10% of the powdered charcoal (w/v) and stirred for 30 min to allow absorption of the antibiotics onto the charcoal particles. Whatman No. 1 filter paper was used to filter the mixture. The antibiotic containing charcoal left in the funnel was eluted with 60 mL absolute methanol. These eluate which contained antibiotics dissolved in methanol was then concentrated in vacuo by the use of a rotary vacuum evaporator at 70 °C to about 2 mL. This partially purified antibiotic metabolites were then bioassayed using the paper disc method as described by Mondal et al. (2001).
Concentrating the antibiotics through freeze drying: Fifteen milliliter of each cell free culture filtrate were placed in sterile universal bottles and kept in a slanting position in the deep freezer and frozen overnight. The caps of these vials were then removed and a muslin cloth fastened at the opening of the vials. The frozen culture filtrates were freeze dried for a second time at a vacuum pressure of 760 mm Hg and at a temperature of -40 °C. The freeze drying was done to a dryness of 10-2 torr until a powder was obtained. This powder obtained from an original 15 mL of cell free culture filtrate was reconstituted with 10.5 mL of distilled water to give a 30% concentrated culture filtrate. The same powder was reconstituted with 7.5 mL of sterile distilled water to give rise to a 50% concentrated culture filtrate. These concentrated antibiotic metabolites were then assayed for antibiotic activity using the paper disc method following the method reported by Mondal et al. (2001).
RESULTS
The three media used namely Czapeks Dox agar, inorganic salts starch agar and glycerol asparagine agar, supported good growth of the three actinomycete isolates. Observations under magnification x400 and x1000 showed the presence of substrate mycelia and sporulating aerial hyphae in all the 3 isolates. The spores occurred in chains and in isolate CS35, spores were occurring in chains of less than 10 whereas in isolate 14P and 28P, the spores were occurring in chains of more than 10. Using an identification guide by International Streptomyces Project (ISP) (Getha et al., 2004), the characteristic of the spore bearing hyphae of the isolate 28P was found to be rectus or straight that of CS35 was flexuous, whereas that of 14P was both flexuous and rectus.
Observations made after 21 days showed a great number of spores scattered on the surface of the media. Germination and new mycelial growth was observed from some of the spores scattered on the surface of the media. Dense aerial growth and sporulation was observed after 21 days making it difficult to do microscopic observations except at the edge of the colonies. Only isolate 14P showed melanoid production and this was on Czapeks Dox agar. The three isolates showed variable results, in the utilization of the carbon sources tested. Isolate 14P utilized all the carbon sources tested, isolate 28P utilized all the carbon sources tested except, cellulose, whereas isolate CS35 utilized all the carbon sources except inositol, raffinose and rhamnose (Table 1).
Production of pigments in the solid media by the actinomycete isolates: The colour of the aerial surface for the isolates 14P and 28P was white and for the isolate CS35, it was light grey in the 3 media tested namely inorganic starch salts agar, glycerol asparagine agar and Czapeks Dox agar. The colour of the substrate mycelium as seen from the reverse side of the plate was pale yellow and pinkish white for the isolates CS35 and 28P respectively in the 3 types of media. Isolate 14P did not produce the reverse color in the inorganic salt starch agar and glycerol asparagine agar but it produced dark reddish brown color on Czapeks Dox Agar. The presence of soluble colour other than melanoid production was observed in isolates CS35 and 14P and the colours noted for the two isolates were yellow and red. Soluble colour other than melanoid production was not observed in isolate 28P. In all the isolates, there was no response of colour to pH change on addition of 0.05 N HCI or 0.05 N NaOH.
| Table 1: | Utilization of carbon sources by the various actinomycete isolates |
 | |
| ++: Strongly positive utilization. Growth on tested carbon in basal medium is equal to or greater than growth on basal medium + glucose, +: Positive utilization. Growth on tested carbon source is significantly better than on basal medium without carbon but somewhat less than on glucose, ± : Utilization doubtful. When growth on tested carbon is only slightly better than on the basal medium without carbon and significantly less than with glucose, -: Utilization negative. When growth is similar to or less than growth on basal medium without carbon | |
From the morphological and physiological characteristics, the actinomycete isolates CS35 was identified to be Streptomyces sahachiroi, actinomycete isolate 14P was identified to be Streptomyces neyagawaensis. The isolate 28P was identified to be Streptomyces hygrostaticus.
Paper chromatography of the antibiotic metabolites: The solvents tested namely methanol, butanol, water, n-hexane and acetic acid eluted all the antibiotics in culture filtrates of isolates 28P and CS35 but n-hexane did not elute the antibiotics in the culture filtrate of actinomycete isolate 14P (Table 2). The results of the bioautography showed that, Rf values were different for each of the antibiotic. Two clear zones were observed in culture filtrates from isolate 28P indicating that this culture filtrate contained two active compounds that were active against Pythium sp. Only one clear zone was observed from culture filtrates from isolate 14P and CS35 indicating that these culture filtrates were each having one compound active against Pythium sp.
| Table 2: | Rf values of antibiotic metabolites from the actinomycete isolates in the various solvent systems |
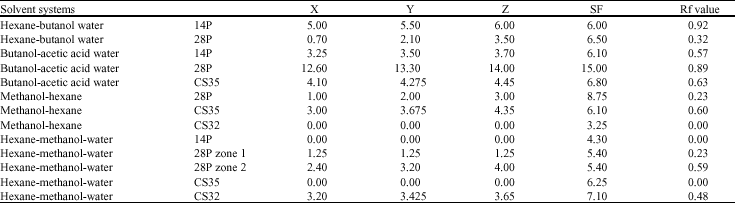 | |
| X: Distance from the origin to the lower end of the clear zone, Y: Distance from the origin to the geometrical centre of the clear zone, Z: Distance from the origin to the upper end of the clear zone, SF: Distance from the origin to the solvent front, Rf value = y/SF | |
| Table 3: | Mean inhibitory diameters of antibiotic metabolites after concentration through freeze drying and partial purification |
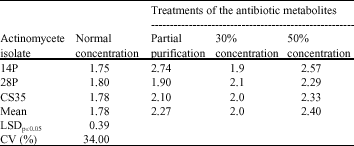 | |
Partial purification of the antibiotic metabolites: There was an increase in activity as evidenced by the increase in size of the colony diameters when partial purification of the culture filtrates of the 3 isolates was done. The activity of the isolate 14P had the highest increase in activity followed by isolate CS35 and 28P in that order (Table 3). This represented an increase of 40.5, 18.3 and 5.6%, respectively as compared to the activity of the normal antibiotic metabolites. Partial purification had a significant effect on the enhancement of activity of the antibiotics in the culture filtrates at p ≤ 0.05.
Concentrating antibiotic metabolites through freeze drying: There was increase in antimicrobial activity when 50% of water was removed as compared to when 30% of water was removed. This increase in activity was evidenced by the increase in the size of the diameters of inhibition zones (Table 3). Concentrating antibiotic metabolites from 30 to 50% represented a percentage increase in antimicrobial activity of 16.5, 9 and 35% for the isolates CS35, 28P and 14P, respectively. The level of activity when the antibiotics were concentrated from 30 to 50% was significantly different at 5% level for all isolates.
DISCUSSION
The three actinomycete isolates were identified as belonging to the genus Streptomyces and this confirms reports that most of the actinomycetes that produce antibiotics are in the Streptomyces genus (Mustafa et al., 2004). Other researchers have also demonstrated that most of the actinomycetes producing potent antibiotics belong to the Streptomyces genus. Although it was possible to identify these actinomycete isolates to the species level based on pigment production, morphological and physiological characteristics, additional features such as studying electron micrographs of the spore surface and biochemical properties can provide more details that can be used for identification purposes as reported by Mustafa et al. (2004). Other advanced methods such as gene analysis of 16S rRNA and fourier transform infrared spectroscopy are more reliable, however these are more expensive and technically challenging (You et al., 2005).
The different media did not have an effect on morphological characteristics of the isolates but it had an effect on the physiological characteristics such as melanin production. Mutitu et al. (2008) also found that, a variety of pigments and colony types are produced by the same organism on different media. For the carbon utilization tests, profuse growth of the actinomycete colonies showed that, the particular carbon source was effectively utilized by the isolate as also reported by Mustafa et al. (2004). Slight or poor growth is an indication that, the particular carbon source is not an adequate source of carbon or the material may contain traces of other compounds (Sathi et al. (2001).
The spore bearing hyphae of isolate coded 28P was found to be both rectus and flexuous hyphae. All the three isolates showed morphology typical of Streptomyces. Concentration of the antibiotic metabolites by partial purification and freeze drying resulted in enhanced antimicrobial activity as shown by the bioassay tests. Related study by Rahman et al. (2000) and Sathi et al. (2001) have shown similar findings. Partial purification elutes the antibiotic from the inert components such as water, media residues and the actinomycete cells thus increasing the antibiotic activity (Mutitu et al., 2008). In this study, concentrated culture filtrates demonstrated increased activity as shown by the in vitro tests.
The antibiotic metabolites showed a high antagonistic potential to the test pathogen and this potential could be enhanced by concentrating the metabolites through partial purification and freeze drying. You et al. (2005) also reported that actinomycetes has a high potential as biocontrol agents for controlling plant diseases. The Rf values from the bioautograms showed the presence of two active compounds from isolate 28P and one active compound from each of the isolates CS35 and 14P and thin layer chromatography should be conducted to confirm these findings as recommended by Hongjuan et al. (2006). The sizes of the inhibition zones varied with the most antagonistic isolate producing bigger zones in the bioautograms. This corroborates reports by Asha Devi et al. (2006) that the size of the inhibition zone positively correlates with the amount of antibiotics produced. Actinomycetes especially those in the genus Streptomyces, produces novel products such as antimicrobial products which have antagonistic potential to plant pathogenic organisms and these can be used in the management of diseases induced by such pathogens. The improvement of antagonistic activity of these metabolites after partial purification and concentration through freeze drying is a further proof of these great potential and more work is needed to evaluate these metabolites in plant disease control under field conditions.
REFERENCES
- Agrios, G.N., 2005. Plant Pathology. 5th Edn., Elsevier, Cambridge, Massachusetts, ISBN-13: 9780080473789, Pages: 922.
Direct Link - Asha Devi, N.K., M. Jeyarani and K. Balakrishnan, 2006. Isolation and identification of marine actinomycetes and their potential in antimicrobial activity. Pak. J. Biol. Sci., 9: 470-472.
CrossRefDirect Link - Getha, K., S. Vikineswary, W.H. Wong, T. Seki, A. Ward and M. Goodfellow, 2004. Characterization of selected isolates of indigenous Streptomyces species and evaluation of their antifungal activity against selected plant pathogenic fungi. Malaysian J. Sci., 23: 37-47.
Direct Link - Hongjuan, Z., L.P. Rachel, I.E. David, W.G. Gareth and G. Royston, 2006. The rapid differentiation of streptomyces using fourier transform infrared spectroscopy. Vibrational Spectroscopy, 40: 213-218.
CrossRefDirect Link - Mutitu, E.W., W.M. Muiru and D.M. Mukunya, 2008. Evaluation of antibiotic metabolites from actinomycete isolates for the control of late blight of tomatoes under greenhouse conditions. Asian J. Plant Sci., 7: 284-290.
CrossRefDirect Link - Oskay, A.M., T. Usame and A. Cem, 2004. Antibacterial activity of some actinomycetes isolated from farming soils of Turkey. Afr. J. Biotechnol., 3: 441-446.
Direct Link - Odakura, Y., H. Kase, S. Itoh, S. Satoh and S. Takasawa et al., 1984. Biosynthesis of astromicin and related antibiotics. ii biosynthetic studies with blocked mutant of Micromonospora olivasterospora. The J. Antibiot., 37: 1670-1680.
Direct Link - Abdur Rahman, M.A., A.T.M. Zafrul Azam and M.A. Gafur, 2000. In vitro antibacterial principles of two flavonoids and extracts from Clerodendrum indicum linn. Pak. J. Biol. Sci., 3: 1769-1771.
CrossRefDirect Link - Sathi, Z.S., M.A.A. Rahman and M.A. Gafur, 2001. Identification and in vitro anti microbial activity of a compound isolated from Streptomyces species. Pak. J. Biol. Sci., 4: 1523-1525.
Direct Link - Sultan, M.Z., N.A. Khatune, Z.S. Sathi, S.A.M.D. Bhuiyan and G.M. Sadik et al., 2002. In vitro antibacterial activity of an active metabolite isolated from Streptomyces species. Biotechnology, 1: 100-106.
CrossRefDirect Link - You, J.L., L.X. Cao, G.F. Liu, S.N. Zhou, H.M. Tan and Y.C. Lin, 2005. Isolation and characterization of actinomycetes antagonistic to pathogenic Vibrio spp. from nearshore marine sediments. World J. Microbiol. Biotechnol., 21: 679-682.
CrossRefDirect Link