
U.A. Naher
Department of Land Management, Universiti Putra Malaysia (UPM), 43400, Serdang, Selangor DE, Malaysia
O. Radziah
Department of Land Management, Universiti Putra Malaysia (UPM), 43400, Serdang, Selangor DE, Malaysia
M.S. Halimi
Department of Agricultural Technology,Universiti Putra Malaysia (UPM), 43400, Serdang, Selangor DE, Malaysia
Z.H. Shamsuddin
Department of Land Management, Universiti Putra Malaysia (UPM), 43400, Serdang, Selangor DE, Malaysia
I.M. Razi
Department of Crop Science, Universiti Putra Malaysia (UPM), 43400, Serdang, Selangor DE, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 6 | Page No.: 1008-1014







ABSTRACT
A study was conducted in in vitro condition to determine the specific growth rate, sugar consumption and generation time of four diazotrophic bacterial species, Burkholderia, Rhizobium and two Corynebacterium sp. in four different carbon substrates. Growth of all bacterial strains was significantly affected by the sugars. Burkholderia sp. (Sb13) rapidly attained maximum population in glucose, fructose and sucrose and the specific growth rate best fitted with exponential model. The cell generation times of Burkholderia sp. (Sb13) in glucose, fructose and sucrose substrate were 0.4, 0.55 and 0.1 h, respectively. Rhizobium sp. (Sb16) followed an exponential growth model in fructose and its generation time was 0.62 h. The growth of Corynebacterium sp. (Sb26) was higher in arabinose and followed logarithmic growth model with generation time of 0.68 h. Corynebacterium sp. (Sb35) showed faster generation time (1 h) in sucrose substrates and the specific growth rate in all four carbon substrates followed logarithmic model of growth phase. Burkholderia sp. and Corynebacterium sp. (Sb26) consumed 100% of arabinose after 36 h of incubation period. The total sugar consumption by the diazotrophs were higher in logarithmic model of growth phase compared to exponential, power and polynomial model.
PDF Abstract XML References Citation
How to cite this article
U.A. Naher, O. Radziah, M.S. Halimi, Z.H. Shamsuddin and I.M. Razi, 2008. Specific Growth Rate and Carbon Sugar Consumption of Diazotrophs Isolated from Rice Rhizosphere. Journal of Biological Sciences, 8: 1008-1014.
DOI: 10.3923/jbs.2008.1008.1014
URL: https://scialert.net/abstract/?doi=jbs.2008.1008.1014
DOI: 10.3923/jbs.2008.1008.1014
URL: https://scialert.net/abstract/?doi=jbs.2008.1008.1014
INTRODUCTION
Diazotrophs are nitrogen fixing bacteria and carbohydrates sugars, sugar alcohols, organic and amino acids are generally considered as carbon and energy sources for the rhizospheric diazotrophs (Chan, 1986). Diazotrophs utilized rhizosphere carbon substrates as their energy and fix nitrogen form the atmosphere and form natural association with plants. About 64-86% of the carbon released in to the rhizosphere is respired by microorganisms (Hutsch et al., 2002). Diazotrophs activity is higher in the vicinity of plant roots due to rhizospheric deposition. A 10-100 fold more microbial population found in the rhizosphere as compared with the surrounding bulk soil (Weller and Thomashow, 1994).
Carbon substrate utilization pattern are definite biochemical characteristics of microorganisms. Diazotrophs shows differences in terms of minimal doubling time, specific growth rate and consumed different amount of carbon substrates as their energy sources. The carbon substrate utilization pattern also differed when there is a mixture carbon compounds instead of sole carbon sources. In mix carbon conditions bacteria often utilized one carbon source preferentially and further carbon sources being consumed when the preferred carbon source utilized (Wendisch et al., 2000). The cell generation time depends on carbon substrates. This may vary from 10 min to several days. Some of the uncultivated bacteria have longer doubling times (Couturier and Rocha, 2006). Bacteria consumed carbon substrates and increases in number following different growth model. The population growth increased in specific substrates and reached in maximum number and after then decreased. The metabolites produced in growth culture media inhibit further populations.
Diazotrophs mostly depends on rhizosphere carbon compounds. Carbon-substrate availability is the most common limiting factor of microbial activity in soil (Paterson et al., 2007). The aim of the present study was to determine the sole carbon substrate utilization pattern of diazotrophs and population growth on the specific carbon substrates. The information can be used to apply specific carbon sources to support and enhance a particular diazotrophs that can be used as a plant growth promoting inoculants to maximize the potential of biological nitrogen fixation in rice cultivation system.
MATERIALS AND METHODS
Burkholderia sp. (Sb13), Rhizobium sp. (Sb16) and two different Corynebacterium spp. (Sb26 and Sb35) capable of high nitrogenases enzyme activity and IAA production (previously isolated from rice root environment) were tested for carbon substrate utilization in glucose, fructose, sucrose and arabinose substrates. The carbohydrate sugars were used as sole carbon source for bacterial growth. Sugar consumption, specific growth rate and doubling time were determined. The experiment was laid out in completely randomized design with 4 replications. Data were analyzed using SAS (9.1version) statistical software.
Substrate preparation: Carbon and nitrogen free nutrient culture solution was modified from Egener et al. (1999) with 15 g of individual sugar (glucose, fructose, sucrose and arabinose) were added and used as growth culture medium. Composition of the medium in 1 L were as follows: KH2PO4, 1.5 g; K2HPO4, 0.33 g; K2SO4, 0.2 g; ferric citrate, 13 mg; CaCl2.2H2O, 0.4 g; MgCl2., 0.4 g; Na2MoO4.2H2O, 2 mg; H3BO3, 3 mg: MnSO4.H2O, 2 mg; ZnSO4.7H2O, 0.2 mg; CuSO4.5H2O, 0.1 mg.
Inoculum preparation: Four diazothrophic species were grown in nutrient agar broth for 48 h and cells were harvested by centrifugation at 13500 rev min-1 for 10 min in eppendorf tube and washed with 0.85% sterilized Phosphate Buffer Saline (PBS) according to Bacteriological Analytical Manual (2001). Composition of PBS (g L-1): 7.650 g NaCl, 0.724 g Na2HPO4 and 0.210 g KH2PO4. After washes the bacterial cells were immediately suspended into PBS solution. Optical density (OD 600) of washed cells were checked and adjusted to 0.1. The population was confirmed by cell enumeration in drop plate method on nutrient agar. Approximately 104 live bacterial cells were applied to each sugar substrates. The inoculated conical flasks were incubated at 28°C temperature on a rotary shaker.
Population growth determination: The bacterial cells were harvested after 6, 12, 18, 24 and 36 h of incubation. Optical density was checked by spectrophotometer at 600 nm wave length. 1 mL of culture solution from each conical flask was diluted 10 fold up to 10-10 and 0.1 mL of each dilution was dropped on nutrient agar plates. Bacterial populations were counted using drop plate count method.
Specific growth rate: Specific growth rate of each bacterial culture at different sampling time was determined as:
where, μ is specific growth rate of individual culture. Value of Z and Z0 represents the amount of any bacterial component of the culture at times t and t0, respectively.
Doubling time: The mean doubling time or generation time were determined as:
where, g is time required for the culture to increase by a factor of 2.
Sugar consumption: After every sampling, the bacterial culture solution of each treatment were filtered through 0.2 μM pore syringe filter and kept in -20°C temperature until analysis. A 20 μL aliquot of samples were injected in high performance liquid chromatography (HPLC, model Jasco Brownian). The amount of residual glucose, fructose, sucrose and arabinose, were determined using Apex column (60°C) and Refractive Index (RI) detector. Acetonitrile (75%) was used as mobile phase with a flow rate of 1.8 mL min-1. A standard series of 0.25, 0.50, 0.75 and 1% sugars were prepared and sugar concentrations were determined using standard graph. The amounts of sugar consumption were determined as:
C = Sto−St |
where, C is Amount of substrate consumption, Sto represent substrate added at initial time and St is the substrate remaining in the culture solution at each sampling time.
RESULTS
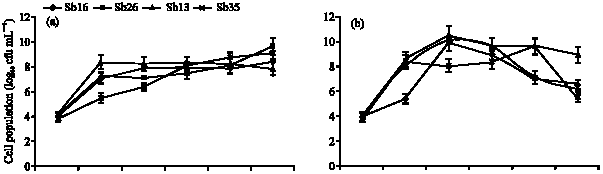
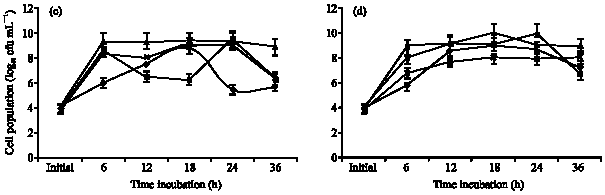
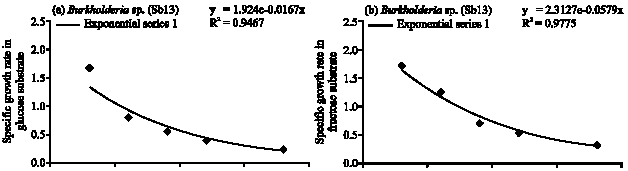
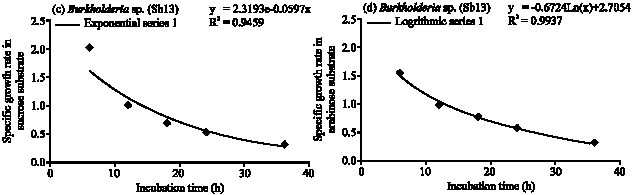
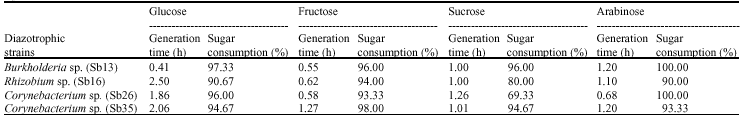
Burkholderia sp. (Sb13): Diazotrophic populations showed a large variance in terms of population growth, generation time and followed different specific growth rate models. The Burkholderia sp. (Sb13) produced maximum population within 6 to 18 h of incubation period in glucose, fructose, sucrose and followed exponential model of specific growth rate (Fig. 2a-c). In glucose Burkholderia sp. obtained maximum population growth at 6 h of incubation period and after then population gradually decreased (Fig. 1). The cell generation time was 0.41 h and consumed 97% of glucose within 36 h (Table 1) of incubation period. In fructose Burkholderia sp. obtained maximum population growth at 12 h. The cell generation time 0.55 h and after 36 h of incubation period consumed 96% of applied fructose. The maximum population obtained at 18 h in sucrose. The doubling time 1 h and consumed 96% of applied sucrose. In arabinose Burkholderia sp. showed a gradually increased of population growth and at 36 h obtained maximum population. The cell generation time was 1.2 h and specific growth rate followed logarithmic model (Fig. 2d). After 36 h of incubation period 100% of arabinose was consumed by the Burkholderia sp.
| Fig. 1: | Comparison growth of different strains in (a) glucose, (b) fructose, (c) sucrose and (d) arabinose substrate. Values are means of 4 replications. Error bars are standard errors of 4 replications, Sb13; Burkholderia sp., Sb16; Rhizobium sp., Sb26; Corynebacterium sp., Sb35; Corynebacterium sp. and Burkholderia sp. (Sb13) |
| Fig. 2: | Specific growth rate (h) of Burkholderia sp. (Sb13) in (a) glucose, (b) fructose, (c) sucrose and (d) arabinose substrates |
| Table 1: | Cell generation time and percent sugar consumption of diazotrophs in different carbon substrates (values were means of 4 replications) |
 | |
| Fig. 3: | Specific growth rate (h) of Rhizobium sp.(Sb16) in (a) glucose, (b) fructose, (c) sucrose and (d) arabinose substrates |
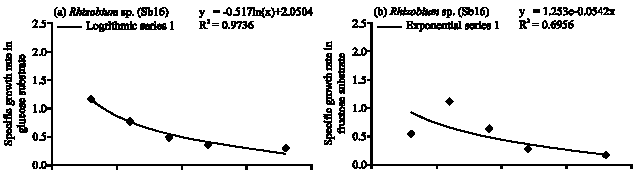
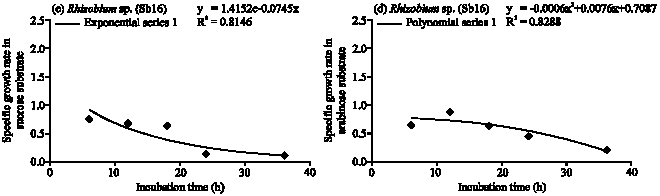
Rhizobium sp. (Sb16): The Rhizobium sp. population gradually increased in glucose substrate and maximum population obtained at 36 h (Fig. 1) of incubation period. The cell doubling time 2.5 h and specific growth rate followed logarithmic model of growth phase (Table 1, Fig. 3a). At 36 h of incubation period Rhizobium sp. consumed 90% of applied glucose. In the fructose, Rhizobium sp. showed comparatively faster growth rate and followed exponential model (Fig. 3b). The maximum population obtained at 12 h and after then population decreased. The cell generation time 0.62 h and consumed 94% of applied fructose (Table 1). The Rhizobium sp. obtained maximum population growth at 18 h of incubation period in sucrose and followed exponential model for specific growth rate (Fig. 3c). The cell doubling time 1 h and after 36 h of incubation period it consumed 80% of applied sucrose. In arabinose substrate, Rhizobium sp. showed polynomial model of growth phase (Fig. 3d) and generation time 1.1 h. It consumed 98% of arabinose after 36 h of incubation period.
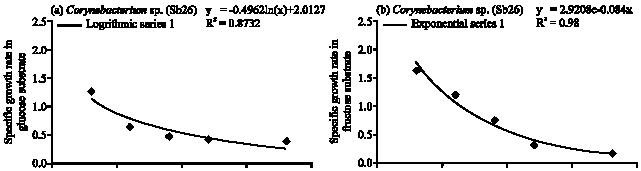
Corynebacterium sp. (Sb 26): The population growth of Corynebacterium sp. (Sb26) in glucose substrate gradually increased and maximum population obtained at 36 h of incubation period following logarithmic model of specific growth rate (Fig. 1, 4a). The cell doubling time 1.86 h and consumed 96% of applied glucose. In the fructose Sb26 showed faster growth rate and maximum population obtained at 12 h. The specific growth rate in fructose followed exponential model. The cell generation time 0.58 h and consumed 93.33% of applied fructose. In the sucrose Sb26 produced maximum population at 24 h of incubation period. For specific growth rate it followed power model. The generation time 1.26 h and consumed 69.33% of applied sucrose. In arabinose, Sb26 obtained maximum population at 24 h and followed logarithmic model for specific growth rate. The cell generation time 0.68 h and it consumed 100% of applied arabinose (Table 1, Fig. 4d).
| Fig. 4: | Specific growth rate (h) of Corynebacterium sp. (Sb26) in (a) glucose, (b) fructose, (c) sucrose and (d) arabinose substrates |
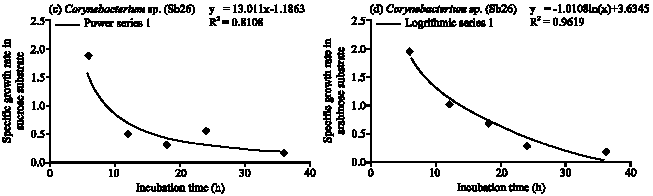
| Fig. 5: | Specific growth rate (h) of Corynebacterium sp.(Sb26) in (a) glucose, (b) fructose, (c) sucrose and (d) arabinose substrates |
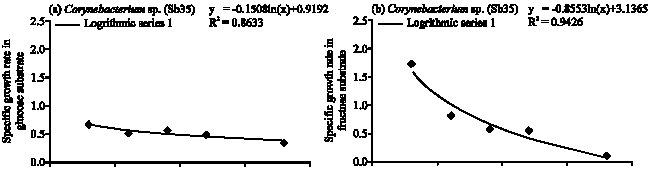
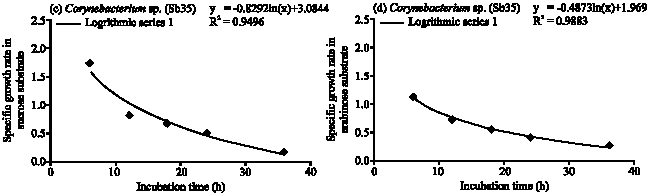
Corynebacterium sp. (Sb35): The specific growth rate of Corynebacterium sp. (Sb 35) followed logarithmic model in four different carbon substrates (Fig. 5a-d). In glucose substrate population increased gradually and reached maximum at 36 h of incubation period (Fig. 1). The cell generation time 2.06 h and consumed 94.64% of applied glucose (Table 1). In the fructose substrate the maximum population obtained at 24 h of incubation period (Fig. 1). The generation time 1.27 h and consumed 98% of applied fructose (Table 1). In the sucrose substrate the maximum population obtained at 24 h and generation time 1.01 h. After 36 h of incubation period Sb35 consumed 94.67% of applied sucrose. In arabinose the generation time 1.2 h and consumed 93.33% of applied arabinose.
DISCUSSION
The population of four diazotrophs showed a large variation in growth rate and generation time in different carbon substrates. Generally eukaryotes followed logarithmic growth model of growth phase. But the specific growth rate highly depended on the substrates and presence of nutrient elements. The generation time of diazotrophs even faster in the presence of nitrogen element in the growth culture media (Chan, 1986). The generation time also depends on substrate affinity. The Burkholderia sp. (Sb13) showed high affinity to glucose and fructose and followed exponential model of growth phase. The cell generation in four carbon substrates were glucose > fructose > sucrose > arabinose. Although the specific growth rate and cell generation was faster in other three substrates but arabinose consumption was higher which indicates the gradual increased in population which followed logarithmic model of growth phase maintained living cells for longer time. The exponential growth phase cause early maximum population growth and further population growth limited by accumulation of inhibitory metabolites or end products. Limitation of biological space and available nutrients also caused rapid death of cells. At the death phase, the number of viable cells decreased exponentially, the reverse growth of log phase (http://www.textbookofbacteriology.net/).
Rhizobium spp. (Sb16) grows rapidly in glucose, arabinose, fructose and sucrose substrates (Vincent et al., 1979). In the present study Sb16 showed high affinity to fructose compared to glucose. The cell generation time were fructose > sucrose > arabinose > glucose. Neal and Walker (1935) found superior growth of some Rhizobium spp. in arabinose and other pentose sugar compared to glucose. Sb16 showed logarithmic model of growth phase in glucose where as in fructose and sucrose it showed exponential growth phase. Among four substrates sucrose consumption is lower this may be due to the rapid breakdown of sucrose in the solution and formed glucose and fructose. Sucrose consumption may not reflect the actual consumption and generation time of microbes in that particular disaccharide sugar.
Corynebacterium spp. (Sb26 and Sb35) showed a large variation in population growth in different substrates. The generation time of Sb26 was fructose > arabinose > sucrose > glucose. Sb26 followed logarithmic model of growth phase in glucose and arabinose. While in fructose it followed exponential model for specific growth rate. Girbal et al. (2000) recorded accelerating growth of C glutanimicum in fructose substrate instead of a constant growth rate on glucose. Seibold et al. (2007) also found early exponential growth of Corynebacterium glutamicum in fructose substrate. The higher arabinose consumption may be due to the gradual increased in population growth which produced less inhibitory metabolites and maintained longer stationary phase of population growth. The Corynebacterium sp. (Sb35) showed prolonged growth period in four carbon substrates compared to other tested diazotrophs. The cell generation time in different substrates were sucrose > arabinose > fructose > glucose. Although the substrate affinity is different but followed logarithmic model of growth phase and consumed almost similar amount of carbon substrates.
The faster generation time and amount of substrate consumption not followed the same pattern. The exponential growth phase doesn`t longer time. The inhibitory product cause rapid cell death in closed culture system. The cell division in a constant rate only depends upon the composition of the growth medium and the conditions of incubation. In the stationary phase viable cells can be determined but some of them may be stopped further generation. Sb16 and Sb26 consumed lower amount of sucrose. The sucrose consumption concealed with glucose and fructose and in mix carbon compound bacteria showed different consumption as well as growth rate than sole carbon source. In the sucrose the cell generation time was ± 1.0 for all four diazotrophs.
CONCLUSION
The diazotrophs exhibited differences in the specific growth rate, generation time and utilized sugar as energy and carbon sources. Burkholderia, Rhizobium and Corynebacterium spp. (Sb26) showed rapid growth in fructose and followed exponential growth model for specific growth rate. From the above study we found that logarithmic growth model sugar consumption were high compared to other exponential, power and polynomial growth model. Burkholderia sp. showed shorter generation time and the Corynebacteria sp. (Sb35) showed longer generation time (>1.0 h) in all four carbon substrates. The application of glucose in Burkholderia sp. (Sb13), fructose in Rhizobium sp. (Sb16) and Corynebacteriaum sp. (Sb26) and sucrose in Corynebacteriaum sp. (Sb35) may accelerate the growth of these diazotrophs in rice production system.
ACKNOWLEDGMENTS
The author is grateful to the Universiti Putra Malaysia and Third World Organization for Women in Science (TWOWS) for providing the financial supports for the Ph.D project.
REFERENCES
- Chan, Y.K., 1986. Utilization of simple phenolics for dinitrogen fixation by soil diazotrophic bacteria. Plant Soil, 90: 141-150.
CrossRefDirect Link - Couturier, E. and P.C.E. Rocha, 2006. Replication-associated gene dosage effects shape the genomes of fast-growing bacteria but only for transcription and translation genes. Mol. Microbiol., 59: 1506-1518.
Direct Link - Egener, T., T. Hurek and B. Reinhold-Hurek, 1999. Endophytic expression of nif genes of Azoarcus sp. strain BH72 in rice roots. Mol. Plant-Microbe Int., 12: 813-819.
CrossRefDirect Link - Girbal, L., L.J. Rols and D.N. Lindley, 2000. Growth rate influences reductive biodegradation of the organophos pesticide demeton by Corynebacterium glutamicum. Biodegradation, 11: 371-376.
Direct Link - Hutsch, B.W., J. Augustin and W. Merbach, 2002. Plant rhizodeposition: An important source for carbon turnover in soils. J. Plant Nutr. Soil Sci., 165: 397-407.
CrossRefDirect Link - Neal, O.R. and R.H. Walker, 1935. Physiological studies on Rhizobium IV. Utilization of carbonaceous materials. J. Bacteriol., 30: 173-187.
PubMedDirect Link - Paterson, E., T. Gebbing, C. Abel, A. Sim and G. Telfer, 2007. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytologist., 173: 600-610.
CrossRefDirect Link - Seibold, G., S. Dempf, J. Schreiner and J.B. Eikmanns, 2007. Glycogen formation in Corynebacterium glutamicum and role of ADP-glucose pyrophosphorylase. Microbiology, 153: 1275-1285.
CrossRefPubMedDirect Link - Weller, D.M. and L.S. Thomashow, 1994. Current Challenges in Introducing Beneficial Microorganisms in to the Rhizosphere. In: Molecular Ecology of Rhizosphere Microorganisms. Biotechnology and the Release of GMO's, O'Gara, F., D.N. Dowling and B. Boesten (Eds.). VCH-Verlagsgesellschaft mbH., Weinheim, Germany, pp: 1-18.
CrossRefDirect Link - Wendisch, F.V., A.A. De Graaf, H. Sham and J.B. Eikmanns, 2000. Quantitative determination of metabolic fluxes during coutilization of two carbon sources: Comparative analyses with Corynebacterium glutamicum during growth on acetate and/or glucose. J. Bacteriol., 182: 3088-3096.
PubMedDirect Link