
M. Sreenivasa Rao
of Chemistry, Chundi Ranganayakulu Post Graduate College, Chilakaluripet, Andhra Pradesh, 522 616, India
B. Rajitha
of Chemistry, Chundi Ranganayakulu Post Graduate College, Chilakaluripet, Andhra Pradesh, 522 616, India
E. Pavitra
of Chemistry, Chundi Ranganayakulu Post Graduate College, Chilakaluripet, Andhra Pradesh, 522 616, India
N. Anjaneyulu
of Chemistry, Chundi Ranganayakulu Post Graduate College, Chilakaluripet, Andhra Pradesh, 522 616, India
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 6 | Page No.: 989-992







ABSTRACT
In the present study, it was investigated the levels of copper in hepatopancreas, hemolymph and the ovary during the ovarian cycle of the white shrimp Litopenaeus vannamei. Ovarian development of L. vannamei was classified into four stages; the previtellogenic stage, vitellogenic stage I, vitellogenic stage II and vitellogenic stage III. The levels of copper in the hepatopancreas, hemolymph and ovary increased with increasing the ovarian index. The protein and hemocyanin concentration were significantly increased from the previtellogenic stage to vitellogenic stage III in the hemolymph. Similarly, the protein concentration significantly increased in the both hepatopancreas and ovary from previtellogenic stage to vitellogenic stage III. The results provide support to the hypothesis that the hepatopancreas is the source of ovarian vitellin (protein). The results also suggest that copper is an important element in the ovarian development of crustaceans.
PDF Abstract XML References Citation
How to cite this article
M. Sreenivasa Rao, B. Rajitha, E. Pavitra and N. Anjaneyulu, 2007. Identification and Changes of Copper Profile in Different Tissues During Vitellogenesis of White Shrimp Litopenaeus vannamei
. Journal of Biological Sciences, 7: 989-992.
DOI: 10.3923/jbs.2007.989.992
URL: https://scialert.net/abstract/?doi=jbs.2007.989.992
DOI: 10.3923/jbs.2007.989.992
URL: https://scialert.net/abstract/?doi=jbs.2007.989.992
INTRODUCTION
One of the most exigent problems in the field of aquaculture technology is that of obtaining mature gametes, since reproductive processes of most of the commercially important species are not fully understood and the mechanisms of ovarian maturation remain to be clarified.
Crustacean vitellogenesis occurs by massive incorporation of lipoglyco-carteno protein, which is synthesized in the female crustaceans during ovarian development (Tsukimura, 2001). It is expected that vitellogenin is synthesized in the hepatopancreas that has many biological functions including absorption, storage, metabolism and detoxification of minerals in crustaceans (Dall and Moriarty, 1983). The hepatopancreas periodically accumulates and releases copper during molting and starvation (Arumugam, 1989) and it has been shown to be the site of hemocyanin synthesis (Spindler et al., 1992). Hemocyanin is a copper containing, multi-subunit protein; it has evolved to carryout the specialized functions of oxygen transport in Arthropod species and Molluscans (Burmester, 2004). The O2 molecule is bound to two copper ions, each of which is coordinated by three histidines. Crustacean hemocyanins are highly variable in quaternary structures (Burmester, 2004). As the main protein component of hemolymph, hemocyanin classically represents up to 95% of the total amount of protein (Sellos et al., 1997). Hemocyanins also gave rise to non-respiratory proteins, which most likely have storage functions (Burmester, 2004).
There has previously been no discussion of a feasible role for copper during the process of vitellogenesis and the idea of this study was to examine variations in ovarian copper content and hemocyanin production in relation to the vitellogenin protein accumulation that occurs during ovarian development of white shrimp L. vannamei.
MATERIALS AND METHODS
Collection and maintenance of shrimps: Live female shrimp (L. vannamei) were collected from a market in Chirala (Andhra Pradesh, India). The shrimps were brought to the laboratory and maintained in the laboratory at 28±1°C in tubs partially filled with aged seawater. They were acclimatized to laboratory conditions (12:12 L : D) for at least 7 days before being used in experiments. The water in the tubs was changed daily. In all experiments only intact, uninjured female shrimps were selected. During their sojourn, the shrimps were fed on shrimp pellet ad libitum. Feeding was stopped one day before the commencement of experiment to avoid changes due to prandial activity.
Isolation of organs: The shrimps were immobilized by chilling on ice for 10 min and body weights the shrimps were determined nearest 1.2 g. The ovarian organs and hepatopancreas were isolated, immediately placed in ice-cold crustacean physiological saline (Van Harreveld, 1936) to scrap off adhesive tissue. The organs were removed from the saline and lightly blotted with the paper towels, weighed wet on an electronic balance. The reproductive stages of ovary in the shrimps were identified according to Nagaraju et al. (2004). The ovarian indices were determined using the standard formula:
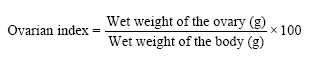 |
Measurement of total proteins: Total protein levels in the hepatopancreas, hemolymph and ovary source were estimated following the method of Bradford (1976).
Estimation of hemocyanin: Hemolymph samples were collected in micro tubes, placed on ice, allowed to clot and frozen at -70°C. The clotted hemolymph was homogenized with a polytron homogenizer and then centrifuged at 10000x g for 50 min and the resulting supernatant decanted and kept at 0°C. Hemocyanin was measured by a method based on that of Johnson et al. (1984). In brief, the hemolymph serum samples were diluted with buffer, 50 mM Tris/10 mM CaCl2 pH 8.0 and reading taken at 280 and 334 nm. The concentration of hemocyanin was calculated with E280 nm = 13.5 and E334 nm + 2.30 as determined for intact undissociated hemocyanin.
Estimation of copper concentration: For copper analysis, hemolymph samples were diluted four fold with deionized water; hepatopancreas and ovaries were wet digested with 4 mL of concentrated HNO3 and the residue was sand dried at 100°C, wet ashed and concentrations determined using flame atomic absorption spectrophotometry (Model 2380, Perkin-Elmer, Norwalk; Engel and Brouwer, 1987).
Statistical analysis: The data were analyzed using one-way ANOVA followed by Student-Newman-Keul’s test to determine the level of significance.
RESULTS AND DISCUSSION
Copper and protein concentrations were determined in hepatopancreas, hemolymph and ovary during vitellogensis and shown to significantly increase during development from the previtellogenic stage to the vitellogenic stage III (p<0.01; Table 1).
The hemocyanin concentrations in hemolymph were also significantly increased during vitellogenesis. The concentration of hemocyanin was significantly increased in hemolymph as the shrimp approached actual maturation (vitellogenic stage III) (Table 2). The relative weight of the ovary (ovarian index) also significantly increased during development from the previtellogenic stage to vitellogenic stage III. The concentrations of copper and hemocyanin from these shrimps were closely related to the stages of ovarian growth and ovarian index.
In shrimps, oogonial proliferation and ovarian differentiation takes place when the ovary is translucent to opaque white (previtellogenic ovary) (Table 1). During vitellogenesis the color of the ovary changes from pale yellow (vitellogenic stage I) to dark yellow (vitellogenic stage II) and then it becomes orange (vitellogenic stage III) to dark orange prior spawning. Maturation of the ovary also includes an increase in size of ovary as the oocytes proliferate and increase in diameter, due to yolk deposition (Table 1 and 2).
| Table 1: | Changes in the ovarian index and hemocyanin concentration (mg mL-1) in hemolymph during different reproductive stages of the white shrimp Litopenaeus vannamei |
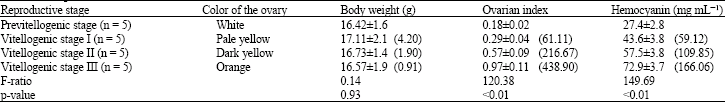 | |
| Values are mean±SD; Values in parentheses are percentage change from control shrimps | |
| Table 2: | Changes of copper levels and protein levels in different tissues during different reproductive stages of the white shrimp Litopenaeus vannamei |
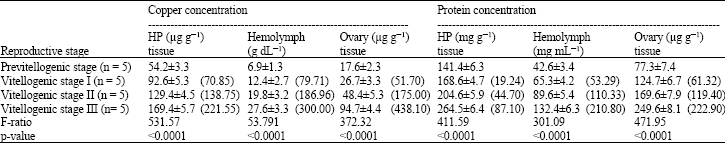 | |
| Values are mean±SD; HP = Hepatopancreas; Values in parentheses are % change from control shrimps | |
Vitellogenin is found in the hemolymph of most crustacean species, being synthesized by extra ovarian tissues and then transported via the hemolymph to the developing oocytes (Tsukimura, 2001; Warrier and Subramonian, 2002). The source of vitellogenin in crustacean ovaries has been the subject of debate for many years, but it is now usually accepted that the hepatopancreas is the main site for vitellogenin synthesis (Charniaux-cotton, 1985; Spindler et al., 1992). Recent use of molecular biology studies showed that expression of vitellogenin in crustaceans occurs in the hepatopancreas and the ovary (Abdu et al., 2002; Okuno et al., 2002). The results presented here are all consistent with the proposal that the source of ovarian vitellogenin in the shrimp is the hepatopancreas .
During the molt cycle, trace metal concentrations in the hepatopancreas and hemocyanin concentration in the hemolymph of blue crabs changes significantly (Engel and Brouwer, 1991). Similarly, the results presented here showed changes of copper and hemocyanin concentration in the shrimp during vitellogenesis. Hemocyanin is synthesized in the hepatopancreas confirming the results obtained in the crayfish using [35S] methionine incorporation (Gellissen et al., 1991), in the blue crab using hybridization of hepatopancreas mRNA with an oligonucleotide based on the amino acid sequence encompassing the copper B binding site (Rainer and Brouwer, 1993) and in prawns (Khayat et al., 1995). Changes of hemocyanin and protein concentrations observed in the white shrimp L. vannamei in relation to size and molt stage (Cheng et al., 2002). The changes of vitellogenin levels were observed in hemolymph and ovaries during ovarian maturation in some crustacean species (Chang and Shin, 1995; Lee and Chang, 1997). Similarly the copper profiles and protein concentration were significantly increased from previtellogenic stage to vitellogenic stage III in all three tissues (hepatopancreas, hemolymph and ovary) of shrimp. We hypothesized that the copper thus accumulated from the medium and released back into hemolymph in the form of hemocyanin (copper-bound protein). During vitellogenesis hemocyanins gave rise non respiratory protein and transformed to ovary.
On the basis of these first results on vitellogenesis in shrimps, we suggest that trace amount of copper may control, at least in part, the accumulation of protein (vitellin) in developing oocytes. Furthermore, this research provides strong supporting evidence to support the hypothesis that an ovarian vitellin source is the hepatopancreas.
ACKNOWLEDGMENTS
We are thankful to Correspondent Sree Chundi Subbarao, Chundi Ranganayakulu Post Graduate College, Chilakaluripet for providing facilities and encouragement. This work was generously supported by the University Grant Commission (UGC) grant, India.
REFERENCES
- Abdu, U., C. Davis, I. Khalaila and A. Sagi, 2002. The vitellogenin cDNA of Cherax quandrucarinatus encodes a lipoprotein with calcium binding ability and its expression is induced following the removal of the androgenic gland in a sexually plastic system. Gen. Comp. Endocrinol., 127: 263-272.
CrossRef - Arumugam, M., 1989. Mobilization of copper among tissues in the estuarine crab Scylla serrata (Forskal) under imposed starvation. Arch. Int. Physiol. Biochem., 97: 247-257.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Burmester, T., 2004. Evolutionary history and diversity of arthropod hemocyanins. Micron, 35: 121-122.
Direct Link - Charniaux-Cotton, C.H., 1985. Vitellogenesis and its control in malacostracan crustacean. Am. Zool., 25: 197-206.
Direct Link - Cheng, W., C.H. Liu, D.F. Yan and J.C. Chen, 2002. Hemolymph oxyhemocyanin, protein, osmolarity and electrolyte levels of white leg shrimp Litopenaeus vannamei in relation to size and molt stage. Aquaculture, 211: 325-339.
Direct Link - Engel, D.W. and M. Brouwer, 1987. Metal regulation and molting in the blue crab, Callinectes sapidus: Metallothionein function in metal metabolism. Biol. Bull., 173: 239-251.
CrossRefDirect Link - Engel, D.W. and M. Brouwer, 1991. Short-term metallothionein and copper changes in blue crabs at ecdysis. Biol. Bull., 180: 447-452.
CrossRefDirect Link - Gellissen, G., R. Henneke and K.D. Spindler, 1991. The site of synthesis of hemocyanin in the crayfish, Astacus leptodactylus. Experientia, 47: 194-195.
CrossRefDirect Link - Johnson, B.A., C. Bonaventura and J. Bonaventura, 1984. Allosteric modulation of Callinectes sapidus hemocyanin by binding of L-lactate. Biochemistry, 23: 872-878.
CrossRefDirect Link - Khayat, M., B. Funkenstein, A. Tietz and E. Lubzens, 1995. In vivo, in vitro and cell-free synthesis of hemocyanin in the shrimp Penaeus semisulcatus (de Haan). Comparat. Biochem. Physiol. Part B: Biochem. Mol. Biol., 112: 31-38.
CrossRefDirect Link - Nagaraju, G.P.C., N.J. Suraj and P.S. Reddy, 2004. Methyl farnesoate stimulates gonad development in Macrobrachium malcolmsonii (H. Milne Edwards) (Decapoda, Palaemonidae). Crustaceana, 76: 1171-1178.
CrossRefDirect Link - Okuno, A., W.J. Yang, V. Jayasankar, H. Saido-Sakanada and D.T.T. Houong et al., 2002. Deduced primary structure of vitellogenin in the gaint freshwater prawn, Macrobrachium rosenbergii and yolk processing during ovarian maturation. Exp. Zool., 292: 417-429.
Direct Link - Rainer, J. and M. Brouwer, 1993. Hemocyanin synthesis in the blue crab Callinectes sapidus. Comp. Biochem. Physiol., 104: 69-73.
CrossRef - Sellos, D., S. Lemoine and A.V. Wormhoudt, 1997. Molecular cloning of hemocyanin cDNA from Penaeus vannamei (Crustacea, Decapoda): Structure, evolution and physiological aspects. FEBS. Lett., 407: 153-158.
CrossRefDirect Link - Spindler, K.D., R. Hennecke and G. Gellissen, 1992. Protein production and the molting cycle in the crayfish Astacus leptodactylus (Nordmann, 1842): II. Hemocyanin and protein synthesis in the midgut gland. Gen. Comp. Endocrinol., 85: 248-253.
CrossRefDirect Link - Tsukimura, B., 2001. Crustacean vitellogenesis: Its role in oocyte development. Am. Zool., 41: 465-476.
CrossRefDirect Link - Van Harreveld, A., 1936. A physiological solution for freshwater crustaceans. Proc. Soc. Exp. Biol. Med., 34: 428-432.
Direct Link - Warrier, S. and T. Subramonian, 2002. Receptor mediated yolk protein uptake in the crab Scylla Serrata, crustacean vitellogenin receptor recognizes related mammalian serum lipoproteins. Mol. Reprod. Dev., 61: 536-548.
Direct Link