
Adel Ramadan El-Shanshoury
Department of Botany, Faculty of Science, Tanta University, Tanta, Egypt
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 6 | Page No.: 918-924







ABSTRACT
Seed protein banding patterns of twenty accessions of V. sativa subspecies sativa were revealed by SDS-PAGE. These have been used to characterize infraspecific genetic variation between accessions collected from different countries. Protein patterns have indicated the existence of low dissimilarity distance within accessions of the same country and a higher degree of dissimilarity between accessions of different countries.Electrophoretic analyses of five isozyme systems (Est, Cbp, Per, Alp and Got) have been carried out on the twenty accession investigated of this subspecies. Variation in allele frequency and number of loci was recorded revealing the existence of homozygosity and heterozygosity. Accessions of the same origin nearly exhibited the same number of alleles and similar intensity of bands in contrast to those of different origin. The results have been discussed with references to the genetic variability within this subspecies as related to difference in origin.
PDF Abstract XML References Citation
How to cite this article
Adel Ramadan El-Shanshoury, 2007. Characterization of Infraspecific Electrophoretic Genetic Variation within Vicia sativa Subspecies sativa Population. Journal of Biological Sciences, 7: 918-924.
DOI: 10.3923/jbs.2007.918.924
URL: https://scialert.net/abstract/?doi=jbs.2007.918.924
DOI: 10.3923/jbs.2007.918.924
URL: https://scialert.net/abstract/?doi=jbs.2007.918.924
INTRODUCTION
The genus Vicia L. belongs to tribe Vicieae of the family Fabaceae. It comprises 180-210 species, some minor food crops and more forage plants. The most common vetch, V. sativa, is cultivated in many countries in Old and New World. The species of Vicia were placed in two subgenera (Cracca and Vicia). The second subgenus Vicia is smaller and can be defined as monotypic that pocesses some synapomorphic characters. The species of this group are more limited and cover mostly the Mediterranean region and near the Middle East countries. Species of this subgenus seem to be a rather newly branch. Its evolution must have been correlated with the development of the more arid region and Mediterranean type of climate. V. sativa aggregates comprise the majority of the taxa of this section. The annual species of this complex are wide spread in Mediterranean countries. This aggregate includes, V. segetales, V. sativa, V. angustifolia, V. incisa and V. cordata. V. sativa is the most widespread and morphologically diversified annual species in Mediterranean countries and Middle East. The taxa of this species are sympatric to a large extent and weedy types and not isolated ecologically. The isolation enforced by their breeding system, spontaneous hybridization and recombination between different karyotypes, as well as chromosomal reconstruction was responsible for the be-wildering chromosomal polymorphism of the species (Mettin and Hanelt, 1973; Holling and Stace, 1974; Zohary and Plitmann, 1979). Vicia sativa subspecies have been object of karyological, cytological and molecular genetic studies. Biosystematic of this species is understood as the introgression of traditional taxonomy by cytology, cytogenetics and ecology. The most striking species diversity is to be found in the Mediterranean region and Caucasus Minor Centers in South Siberia and South America (Hanelt and Mettin, 1989).
Many characters vary considerably within the species but few prove useful for infraspecific grouping. Among these are characters of the morphology of the flower, the leaf, the pod shape and the seed (Kupicha, 1976). Biochemical data would be still more useful if the analyses covered a wide range of samples distribution (Yasuit et al., 1987; Perrino et al., 1989). Karyological characters (x = 7, 6, 5) exhibit a large variability between accessions of V. sativa especially in regard to the morphology of chromosomes and the DNA content of the genome. These are most useful on the species level for the characterization of smaller groups and as an indicator of evolutionary tendencies (Kupichia, 1976). A bivalent formation with high percentage within V. sativa species implies a considerable chromosomal homology between its taxa (Yamamoto, 1977, 1980).
There is a direct relation between phenotype and genotype. Crossing is necessary to demonstrate that a band on an electrophoretic gel does indeed correspond to an allelic form of a single gene. Counts of genotypes lead to simple measures of variation such as number of protein bands or alleles per locus. The number of alleles per locus may be sufficient for the purpose of establishing existence of variation and frequency of heterozygotes could be related to population size (Laurie-Ahlberg and Weir, 1979).
Evolution at the morphology and molecular levels is uncoupled (Wake, 1981), so protein studies often reveal discordant geographic patterns between levels of isozyme divergence and taxonomic boundaries inferred from morphological criteria. Isozyme studies can mesh genetic and ecological information to strengthen inference about specific aspects of population structure, especially breeding structure and effective gene flow (Hamrick and Godt, 1997). Haider and El-Shanshoury (2000) studied the variability of storage seed proteins and esterase isozyme among 20 samples belongs to six subspecies of V. sativa. Results revealed a high degree of similarity between accessions of the same subspecies. The isozyme characters of gene number can be of systematic value only if they vary at a taxonomic level (Whitt, 1987). These characters could be useful for intraspecific, intrageneric or intrafamilial composition. Changes in allelic frequencies in accessions could be attributed to specific mechanism of selection, migration or mutation acting at different stages of population (Chowdhury and Slinkard, 2000).
In this study, electrophoretic techniques on protein and isozyme are used for discriminating genetic heterogeneity among population belonging to the same subspecies reflected by variation in mating structure.
MATERIALS AND METHODS
Materials of 20 accessions representing Vicia sativa subspecies sativa were obtained as seeds from the International Center for Agricultural Research in Dry Area (ICARDA), Aleppo, Syria (Table 1). For seed protein electrophoresis, seeds were milled under cooling at 4 °C by using electric mill for 1 min and the total proteins were extracted for 3 h at 4 °C in 20% sucrose. The extract was centrifuged at 15,000 rpm for 15 min and the supernatant (protein extract) was used directly for electrophoresis or kept in deep freezer (-18 °C) until use. Sixty to eighty microlitre of the protein extract were mixed with equal volume of sample buffer (0.125 M Tris/HCl, pH 6.8, 2% (w/v) SDS, 10% sucrose, 1% (v/v) B-mercaptoethanol, 0.15 (m/v bromophenol blue), denaturation by heating in a boiling water bath for four min, 3 μL of bromophenol blue were then added and 40 μL of the mixture were loaded for electrophoresis.
Electrophoresis was carried out using 12% SDS-poyacrylamide, gel slabs prepared as described by Laemmli (1970). Equal amounts of protein extracts were loaded per track. Tris/glycine-SDS running buffer was used at 100 voltages for 30 min followed by 150 Voltage until the indicator dye reached the bottom of the gel. The gels were directly placed in commassie brilliant blue staining solution overnight and de-stained several times in the de-staining solution.
| Table 1: | Origin of Vicia sativa subspecies sativa accessions, their strain numbers, seed size, seed coat and cotyledon color as obtained from ICARDA |
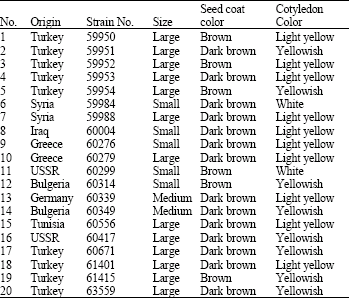 | |
| Large seed = Weight of 100 seeds = More than 6 g. Medium seed = Weight of 100 seeds = Ranged from 4.6 to 6 g. Small seed = Weight of 100 seeds = Ranged from 3 to 4.5 g. | |
The gels were then photographed. The seed protein bands produced by each sample were counted and compared according to its relative mobility with those of other accessions. The presence or absence of each band was treated as binary character in data matrix (coded 1-0) for computation. The data matrix was standardized and analyzed using the NTSYS-pc (Rohlf, 1993). For isozyme electrophoresis, the same protein extract was used. Aliquots of this extract were electrophoresed in 10% slab polyacrylamide gels. The gels of esteraee (Est), calcium binding protein (Cbp), peroxidase (Per), alkaline phosphatase (Alp) and glutamate oxaloacetate transaminase (Got) were stained for the desired enzymes according to the protocols described by Soltis et al. (1983). After staining the gels, bands having very close value of (Rf) were grouped as members of distinct zone (locus). When these zones were expressed independently, they were considered as an expression of a single locus and the bands within this zone were dealt with as an expression of alleles within that locus. Calculation of allele frequency was carried out to find a correlation of individual traits. The data matrix was analysed using the NTSYS-pc Rohlf (1993).
RESULTS
Seed proteins of twenty accessions of V. sativa subspecies sativa were analysed. These accessions represented eight countries and exhibited different seed size and coat and cotyledon colours (Table 1). Fourteen bands were revealed under reducing condition. More stability of some bands was scored within accessions collected from the same country. This reflects high similarity in genetic construction of these accessions. The total number of bands in each accession ranged from 13 bands in accession (18); collected from Turkey; to 4 bands in accession (6); collected from Syria; as shown in Fig. 1. NTSYS depends on number of protein bands revealed as a degree of dissimilarity distance between accessions. The dendrogram based on protein banding patterns demonstrating delimitation and relationships of the examined accessions is given in Fig. 2. Accessions under investigation are delimited into
two major groups (a and b) at 1.22 dissimilarity distance. Group (a) included 9 accessions (1, 3, 4, 10, 18, 2, 5, 6 and 7) six of which originating from Turkey, two from Syria and one accession originating from Greece.
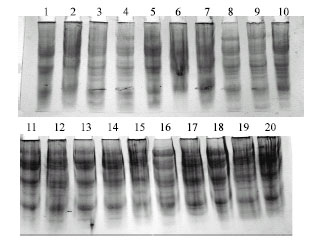 | |
| Fig. 1: | Electrophenogram of 20 accessions of V. sativa subspecies Sativa showing SDS-PAGE patterns of seed proteins, accessions are numbered as Table 1 |
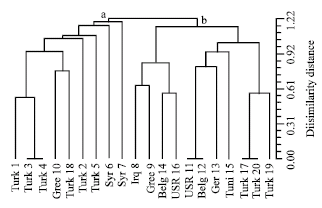 | |
| Fig. 2: | Phenogram cluster analysis of 20 accessions of V. sativa subspecies sativa (numbered in Table 1) based on dissimilarity distance matrix of SDS-PAGE protein banding pattern |
Accession 1, 3 and 4 (brownish seed coat, large seed, light yellow cotyledon) are grouped nearly together, while accessions 10 and 18 with large seeds, dark brown seed coat and light yellow cotyledon are grouped together. Accessions 2, 5 and 6 delimited as a single branch nearly below 1.22 dissimilarity distances. Accession 6 which has small seeds and white cotyledon is separated alone as a branch, while accessions 2 and 5 each with large seeds, yellowish cotyledon and varied only in color of seed coat from dark brown to brown respectively. Accession 7 collected from Syria, is delimited as a single branch in this group. This could be because it has large dark brown seeds with light yellow cotyledon when compared with accession 6, collected from the same country. Group (b) included eleven accessions (8, 9, 14, 16, 11, 12, 13, 15, 17, 20 and 19). Three of these accessions 17, 18 and 20 originate from Turkey and delimited together. Their seeds are large with dark brown coat and yellowish cotyledons and share most of protein bands. Under the same group the accessions (11, 12, 13 and 15) are delimited almost together in a separate subgroup. Accessions 11 and 12 each is small seeded with brown coat but slightly differ in color of cotyledons. Accessions 13 and 15 both with dark brown seed coat and light yellow cotyledons but differ in seed size. The remaining accessions (8, 9, 14 and 16) were delimited together at dissimilarity distance less than 0.92. Grouping of 8 and 9 accessions each is small seeded with dark brown coats and light yellow cotyledons. While accessions 14 and 16 grouped at nearly below 0.6 dissimilarity distance varied in seed size and shared cotyledon and seed coat color.
The obtained results depending on the number of alleles and the number of loci of five isozymes systems studied as the products of gene expression in V. sativa subspecies sativa accessions were analyzed using NTSYS (Fig. 3) and their patterns are shown in Fig. 4. These results can be summarized as follow:
Esterase (Est): Six zones of activity were detected. Est1, Est2 and Est3 showed a monomer heterozygote represented by two alleles in some accessions and one allele in the others. On the other hand, the two loci (Est4 and Est6) were exhibited as monomer heterozygote locus represented by one allele. Only Est5 was represented as a dimmer heterozygous locus with three alleles.
Calcium binding protein (Cbp): Six zones of activity were detected in all accessions. Only Cbp1 was represented by one allele as a monomer heterozygote band. Loci of Cbp2, Cbp3 and Cbp6 were detected in accessions as a monomer heterozygote locus expressed by two alleles. The loci Cbp4 and Cbp5 showed a dimer heterozygote with variable triallelic locus each.
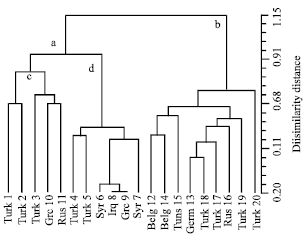 | |
| Fig. 3: | Phenogram cluster analysis of 20 accessions of V. sativa subspecies sativa (numbered in Table 1) based on dissimilarity distance matrix of allele frequencies of different isozyme loci of five isozyme systems [Est, Cbp, Per, Alp and Got) |
Alkaline phosphatase (Alp): Two zones of activity in this isozyme were detected Alp1 and Alp2. This isozyme was detected as a monomer heterozygote in all accessions under study.
Peroxidase (Per): This isozyme showed the simplest patterns of expression as a single locus showing one zone of activity produced by one allele. This isozyme was a monomer and homozygote.
Glutamate oxalacetate transferase (Got): Three zones of activity of this isozyme were detected. These are, Got1, Got2 and Got3 in three loci. All the three loci were made up of diallelic products as a monomer heterozygote.
The NTSYS analysis based on the number of alleles of the five isoenzyme system of the 20 accessions under investigation is shown in Fig. 3. The dendrogram delimited all accessions into two main groups (a and b). Group (a) included 11 accessions and divided into two clusters (c and d). Cluster (c) included 5 accessions (1, 2, 3, 10 and 11), 3 of which (1, 2 and 3) originating from Turkey nearly delimited together and two from Greece and USSR also delimited together. Cluster (d) includes 6 accessions (4, 5, 6, 8, 9 and 7). Two accessions (4 and 5) originate from Turkey and 4 accessions (6, 8, 9 and 7) originate from Syria, Iraq and Greece. The second group (b) included 9 accessions. Four of which originating from Turkey (18, 17, 19 and 20) were grouped together with two accessions from Germany and USSR (13 and 16).
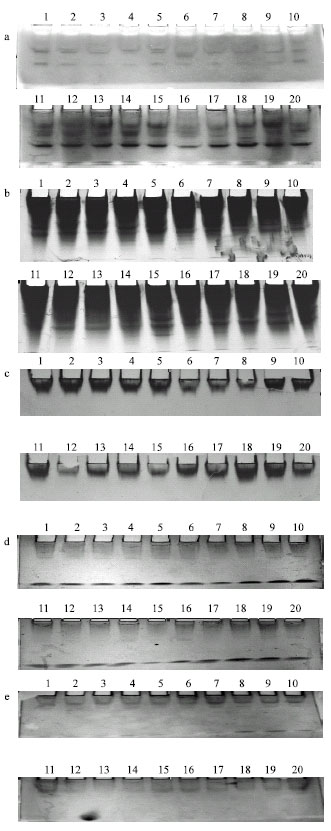 | |
| Fig. 4: | Zygograms of five isozyme systems investigated in accessions of V. sativa subspecies sativa. (a = Est, b = Cbp, c = Per, d = Alp and e = Got) |
The remaining three accessions (12, 14 and 15) which originating from Bulgaria and Tunisia were delimited together.
DISCUSSION
Genetic variation was observed among accessions of V. sativa subspecies sativa based on both intra and inter geographical variation in protein and isozyme patterns. This variation observed in accessions from different geographical regions suggests a high genetic diversity among genotypes of this subspecies. This genetic diversity could be reflected in the ability of this subspecies to grow in wide range in Middle East and Mediterranean of geographical and climatic variables. The protein banding pattern of a plant species reflects its genetic constitution and any slight change in the latter is expressed as deviation from the base banding pattern of storage seed protein. Therefore protein banding technique has been applied to elucidate both intra and inter population genetic variation in various plant species. The genetic variation could be accumulated due to different factors. The presence of genetic diversification in bimolecular analysis of protein and isozyme patterns within a single subspecies reflect the broad distribution of this subspecies in temperate and Eurasian. The annularity of this subspecies correlated with the reduction in vegetative growth of inflorescence and flower tendency to autogamy suppose the origin of annual from the ancestral perennial species. These confirm that karyotype variation may be attributed to the variation in chiasms frequency leading to geographical morphological diversification (Ehrendorfer, 1988). The genomic contents (1c) value of DNA in V. sativa ranging from 2.0-13.9 pg. The variable amount of DNA contents correlated within the total genome length and changes in duration of cell cycle due to different proportion of heterochromatin contents (Chooi, 1971). Variation in meiotic cell cycle and genome size is congruent with morphological variation as well as life cycle (Badr, 2006) could contribute to the adaptability of the plant species to a wide range of environmental conditions and hence lead to establishment and maintenance of the diversity upon which evolution feed.
Reproduction of Vicia species happens by means of seeds, seeds production results from cross pollination. Variation in size of seeds may be related to variation in genome size (Chung et al., 1998) as well as postulated in the nucleotype hypothesis (Bennett, 1972). Vicia flowers are adapted to pollination by hymenoptera. A shift from allogamy to autogamy occurred indepently in annual species groups of both subgenera of Vicia. Certain amount of cross pollination may occur for V. sativa and it has been estimated at approximately 10% (Hanelt and Mettin, 1970). This confirms that widely distributed species tends to maintain more variation (allogamy) than more narrowly distributed species. This variation results in diversification in gene products (protein and isozyme patterns). This variation found in V. sativa subspecies sativa also correlated with variation in Geographic range.
Isozyme is practical and useful genetical and ecological information to strengthen inference about specific aspects to estimate genetic variability in plant population (Hamrick and Godt, 1997). The present results isozymes especially on Est and Cbp revealed a high difference in allelic locus and difference in allele per locus.
The phenogram based on the number of alleles of the five isozyme systems for the 20 accession under investigation is shown in Fig. 3. Delimitation of these accessions into groups having a degree of dissimilarity coefficient revealed a genetic variability within this subspecies. This is may be attributed to geographic isolation as one of the important factors responsible for genetic distance disparity as reported by Tadesse and Bekele (2001). The wide polymorphism of the enzyme system from monomer heterozygote expressed by one allele and dimmer heterozygote by two or three alleles in some loci for some accessions and differential staining intensities may be attributed to difference in ploidy level (Dessaure and Cdle, 1984) which mean gene dosage and ploidy level could be inferred accurately from staining intensities because subunit interactions were additive. This alternative alleles are selectively equivalent and may represent transient stages of replacement with fixation probably being a function of specific mechanism of selection, migration or mutation acting at different stages of population (Kimura, 1983; Yunus et al., 1991; Alfaro et al., 2000; Alba et al., 2001).
On the other hand, the presence of a relatively genetic diversity restricted to homozygosity or heterozygosity in structure of alleles is due to the dynamics of the hybrid zone (Halkka et al., 1987) and reveal the phylogenetic relationships between accessions (Crawford, 1983). These variation might have resulted from variation in type of pollination (this species can be pollinated either by its own pollen or by foreign pollen which depends on environmental conditions. The observed allelic differentiation between sites in inter-population under study reflects the dispersal and diversity of this subspecies in the Mediterranean region and Middle East. The presence of some population genetic surveys revealed heterozygotic deficiency; this could result from strong selection from heterozygous genotypes and inbreeding type (Knight and Waller, 1987). These results showed that low level of gene flow in the annual plant species correlated with population sub-structuring.
To conclude the most significant factors that affect genetic diversity in this subspecies, include, geographical distribution, level of gene flow in this annual subspecies, type of pollination (autogamy and allogamy), chromosomal change in number and mutation leading to new chromosomal rearrangements and change in gene order along chromosomes. These factors promote the dynamics of diversification and sub-structuring of intra-population.
REFERENCES
- Badr, S.F., 2006. Karyotype analysis and chromosome evolution in species of Lathyrus (Fabaceae). Cytologia, 71: 447-455.
Direct Link - Bennet, M.D., 1972. Nuclear DNA content and minimum generation time in herbaceous plants. Proc. R. Soc. Lond. B, 181: 109-135.
CrossRefDirect Link - Chowdhury, M.A. and A.E. Slinkard, 2000. Natural outcrossing in grass pea (Lathyrus sativus L.) Genet. Resour. Crop Evol., 47: 163-169.
Direct Link - Haider, A.S. and A.R. El-Shanshoury, 2000. Variability of storage proteins and esterase isozymes in Vicia sativa subspecies. Biol. Plant, 43: 205-209.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link