
Shirin Honarbakhsh
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
Mojtaba Zaghari
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
Mahmood Shivazad
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 6 | Page No.: 860-864







ABSTRACT
This research tried to examine the effect of exogenous betaine as an osmoprotectant in broiler chicks under water salinity stress. An in vivo CRD experiment, in factorial arrangement of dietary betaine supplementation (0.000, 0.075, 0.150 and 0.225%) and levels of water TDS (375, 1375 and 2375 mg LG1) was investigated on 576 day-old male chicks which were distributed in four replicates. In the first 10 days they were raised without added betaine and TDS of their water was 375 mg LG1. The main experiment started at 11 days after unified the body weight mean of all floor pens. Betaine decreased packed cell volume (p< 0.05). Water consumption (28 and 42 days) and excreta moisture (28 days) were increased by elevating the level of water salinity (p< 0.01). Interaction between dietary betaine and saline water were significant on plasma osmolarity at 28 days and epithelia osmolarity of duodenum. Epithelia osmolarity was decreased from duodenum to ileum. It seems that body homeostatic regulation system defends against changes of blood osmotic pressure parameters and increased total volume of blood.
PDF Abstract XML References Citation
How to cite this article
Shirin Honarbakhsh, Mojtaba Zaghari and Mahmood Shivazad, 2007. The Effect of Betaine on Water Salinity Tolerance in Broiler Chicks. Journal of Biological Sciences, 7: 860-864.
DOI: 10.3923/jbs.2007.860.864
URL: https://scialert.net/abstract/?doi=jbs.2007.860.864
DOI: 10.3923/jbs.2007.860.864
URL: https://scialert.net/abstract/?doi=jbs.2007.860.864
INTRODUCTION
In many parts of the world most of water sources, which are available for poultry production, have high levels of TDS. Drinking water with high levels of TDS has negative effects on broiler performance. This response has relationship with osmotic regulation which is important for the optimal function of intracellular macromolecules (Kettunen et al., 2001). For example, intestinal cells that always have to cope with variable osmotic media since the luminal content of the intestine is hyper osmotic in relation to blood plasma (Eklund et al., 2005). Moreover, the process of nutrient digestion and absorption necessitate osmolytic protection mechanisms since intestinal cells mediate the exchange of water, small solutes such as ions, nutrients and macromolecules between plasma and intestinal fluid (Eklund et al., 2005). In order to get rid of this problem, glycine betaine is one of the most likely candidates for the task (Kettunen et al., 2001), because it’s thought to be an important organic osmolyte for controlling the osmotic pressure inside the intestinal cells (Eklund et al., 2005). Due to betaine chemical structure, it assists in cellular water homeostasis (Klasing et al., 2002). Tissues that rely on zwitterionic betaine as an osmolyte include the intestines, kidney, liver, brain and leukocytes (Klasing et al., 2002). This research investigated the effect of betaine as an organic compatible osmolyte, which was named one of the most likely candidates for osmoregulation according to the report of Kettunen et al. (2001). These researchers suggested that the presence of betaine helped the duodenal, but not jejunal, epithelium to maintain water balance in hyperosmotic conditions. Their results showed that betaine in the hyperosmotic saline, significantly decreased the rate of tritium accumulation into the tissue slices of intestine, indicating that betaine slowed down the influx of water to the epithelium. They concluded that betaine affects the movement of water across the small intestine epithelium of broiler chicks in vitro. To date, concerning to our literature review, no information has been published about the effects of betaine on water salinity tolerance in broiler chicks. The aim of present study was to examine the effect of exogenous betaine as an osmoprotectant in response to saline water consumption in broiler chickens.
MATERIALS AND METHODS General: This study was carried from 4 Jan to 14 Feb 2006 at the Research Center of Animal Science Department, Faculty of Agriculture, at the University of Tehran. Commercial male broiler chicks (Ross 308) were used in a factorial arrangement of treatments in a CRD experiment with 4 levels of added betaine (0.000, 0.075, 0.150 and 0.225%) x 3 levels of Total Dissolved Solids (TDS) in drinking water (375, 1375 and 2375 mg L-1) each in 4 replicates. Levels of TDS were made by adding NaCl to underground water source. TDS of the basal water was 375 mg L-1 which was measured according to A.O.A.C. (2000). Diet provided nutrient requirements for broiler chicks according to Ross 308 Management Manual (2002) and the met+cys levels were exactly met the requirement, which eliminates the methionine-sparing effect of betaine. Feed and water consumed ad libitum. The composition of the basal diet and the calculated nutrients content are presented in Table 1. All diets for each period were prepared with the same batch of ingredients and all diets within a period had the same composition except for the supplemental betaine (Betafin S1, 960 g kg-1 Betaine). Betaine and NaCl were added to basal diet and underground water source, respectively, from 11 days when the main experiment started. Thus, the experimental period was subdivided into grower (11 to 28 days) and finisher (29 to 42 days) periods. Prior to begin the trial, two birds of each replicate chosen randomly and wing banded for taken blood and intestinal samples during the experiment.
At 28 and 42 days water consumption was measured during 24 h. Plastic bell drinkers with one liter capacity were used (one drinker per pen). Four similar drinkers which were inaccessible to the birds were placed at various locations in the house for estimating evaporative water loss. Fecal samples were collected separately from each pen, immediately after excretion, at 28 and 42 days in order to measure excreta moisture. Parts of litter which were attached to excreta, separated by forceps. Excreta were subsequently dried in an oven at 65°C for 72 h (Maiorka et al., 2004). Two milliliter of blood was taken from the brachial vein of wing banded birds and centrifuged immediately at 2500 rpm for 10 min (Mirsalimi et al., 1992), at 28 and 42 days. Plasma was separated from the buffy coat and red blood cell sediment. Separated plasma was used for determination plasma osmolarity and plasma sodium (Na), K, chloride (Cl) and albumin concentration. Plasma Na and K concentration were measured by flame photometer and colorimetric procedure was used to determine the amount of Cl and albumin of plasma. At 42 days of age two wing banded birds of each pen were slaughtered and immediately intestinal samples were taken from these birds for evaluation osmolarity along their intestine. Segments (5 cm) were obtained from the duodenum, the jejunum at a position midway between Meckel’s diverticulum and the entrance of the bile ducts and the ileum at a position midway between Meckel’s diverticulum and the ileocaecal junction. The content of the lumen were flushed vigorously with saline to remove the digesta. Washed segments were opened longitudinally and scraped with glass slides to obtain the mucosa and measured how much we collected. Then add 10 times that measured scrapings as pure deionized water and the suspension was homogenized (Klasing et al., 2002).
| Table 1: | Composition of basal diet in starter, grower and finisher periods |
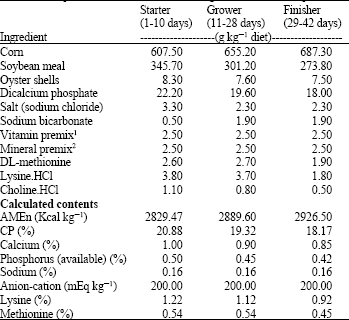 | |
| 1Provided the following per kilogram of broiler diet: Vitamin A,9000 IU; Cholecalciferol, 2000 IU; Vitamin E,18 IU; Vitamin K3, 2 mg; Vitamin B12, 0.15 mg; Biotin, 0.1 mg; Folacin, 1 mg; Niacin, 30 mg; Calcium pantothenat, 10 mg; Pyridoxine , 3 mg; Riboflavin, 6.6 mg; Thiamine 1.8 mg, choline 500 mg. 2Provided the following per kilogram of broiler diet: Copper (as cupric sulfate 5H2O), 10 mg; Iodine (as calciumiodate), 1 mg; Iron (as ferrous sulfate 7H2O), 50 mg; Manganese (as manganese oxide), 100 mg; Selenium (as sodium selenite), 0.2 mg; Zinc (as zinc oxide), 100 mg | |
Osmolarity was determined by using a freezing point osmometer (Cryoscopic osmometer, osmomat 030, Genotec). The dilution would then 10 fold so we would multiply the osmolarity that we measured by a factor of 10. Lack of any external liquid such as water, blood and etc, on the surface where the sampling took place was attended. Thus, the instruments were cleaned and dried regularly.
Statistical analysis: Data were analyzed by using the General Linear Models procedure of SAS (2001) software appropriate for completely randomized design. Significant difference among individual group means was determined with Duncan’s multiple range test option of the GLM procedure of SAS software.
RESULTS
Effect of betaine: Results presented in Table 2 shows that betaine supplementation, only increased plasma Na concentration at 28 days (p<0.05). Cubic relationship was observed between percentage of dietary added betaine and plasma Na concentration at 28 days:
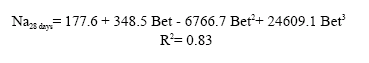 |
| Table 2: | Effect of dietary betaine and TDS of water on blood osmotic pressure parameters of broiler chicks † |
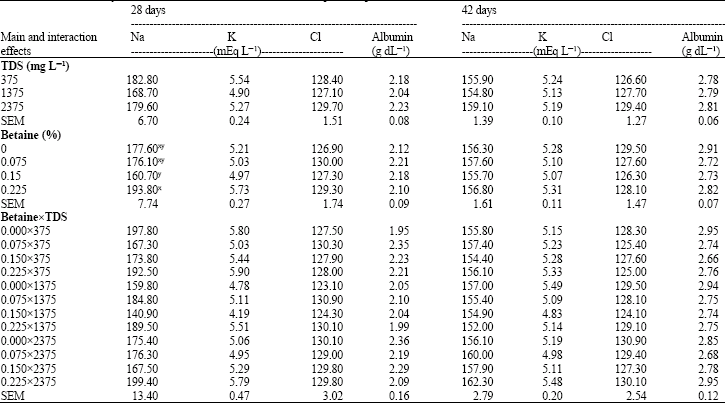 | |
| † x,y Means in a column with different superscripts differ significantly (p< 0.05) | |
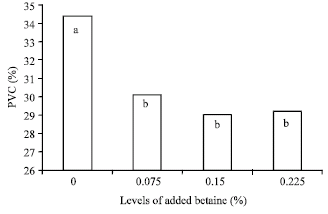 | |
| Fig. 1: | Effect of dietary added betaine on PCV in broiler chicks. Different letter above the columns denotes a significant difference between means (p<0.01) |
Figure 1 implies that dietary added betaine resulted to decrease Packed Cell Volume (PCV) at 42 days (p<0.01). PCV was significantly correlated to levels of betaine (R = -0.47). Obtained relationship between percentage of dietary added betaine and PCV (%) is following below:
 |
No significant effect of supplemented betaine on water consumption and excreta moisture was found (Table 3). At 42 days, excreta moisture had minus correlation with dietary added betaine (p<0.05).
| Table 3: | Water consumption and excreta moisture of chicks, fed different levels of dietary betaine and water TDS† |
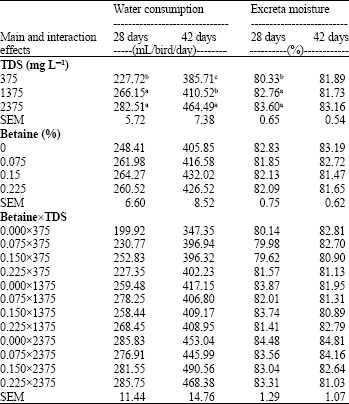 | |
| † a, b,c Means in a column with different superscripts differ significantly (p<0.01) | |
Betaine had no significant effect on plasma osmolarity and also epithelia osmolarity of small intestine (Table 4).
| Table 4: | Osmolarity of blood plasma and intestinal epithelia of broiler chicks fed different levels of dietary betaine and water TDS †, ‡ |
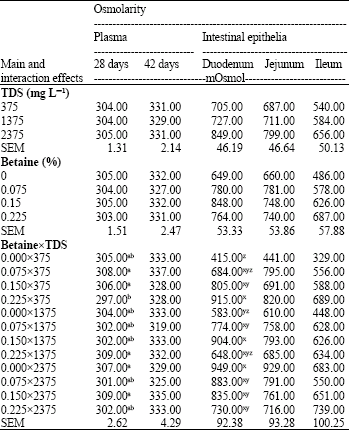 | |
| † a, b Means in a column with different superscripts differ significantly (p<0.01). ‡ x,y,z Means in a column with different superscripts differ significantly (p<0.05) | |
Effect of TDS: Water consumption was correlated to TDS levels significantly (R = 0.7). The obtained relationship between water consumption (42 days) and TDS, also excreta moisture (28 days) and TDS is following below:
According to the results, higher levels of TDS increased osmolarity of duodenum, jejunum, ileum (42 days) and plasma osmolarity at 28 days but these results were not significant (p>0.05). Interaction between dietary added betaine and drinking water TDS on plasma osmolarity (28 days) and duodenum epithelia osmolarity were significant (p<0.05).
DISCUSSION
Effect of betaine: Table 3 shows that excreta moisture of the groups which were supplemented by 0.075, 0.150 and 0.225% of betaine, was lower about 1.18, 0.85 and 0.89% compare with the control group at 28 days (p>0.05). Comparing the excreta moisture at 42 days for the same levels of betaine with the control group showed 0.56, 2.07 and 1.85 percent decrease, respectively (p>0.05).
Interestingly, betaine decreased PCV (Fig. 1). To present knowledge, decreased PCV duo to betaine has not been reported previously. It seems that body homeostatic regulation system defends against changes of blood parameters and increased total volume of blood. Data imply that betaine helped minimized water loss despite a prevailing osmotic gradient (betainexTDS: 0x2375, 0.075x2375, 0.150x2375 and 0.225x2375 in Table 4 for duodenum and jejunum). Eklund et al. (2005) noted that betaine exerts an osmoprotective effect by accumulation in cell organelles and cells exposed to osmotic and ionic stress, thereby replacing inorganic ions and protecting enzymes as well as cell membranes from inactivation by inorganic ions. Thus, water homeostasis is an important factor for cells exposed to different osmotic pressures. Osmotic protection would allow for the maintenance of water balance and intestinal cell volume, thereby facilitating secretion of digestive enzymes (Eklund et al., 2005). According to Eklund et al. (2005) if betaine stimulates cell proliferation in the intestinal tissue, the enlarged gut wall epithelium would provide an increased surface for nutrient absorption. In present study, osmolarity was measured by scraping the intestinal epithelium away from the underlying sub mucosa. Thus the values represent solute concentrations within the cells of the villi, as well as intestinal fluids and any luminal fluids trapped in the folds and not removed during washing. Overall mean of duodenum, jejunum and ileum epithelia osmolarity (760, 732 and 593 mOsm) showed that osmolarity was decreased from duodenum to ileum. This result is in agreement with those reported by Klasing et al. (2002). Present observed value of ~760 mOsmol in the duodenum is very hyper osmotic compared with normal plasma, which we found to be 330.5 mOsmol.
The chicks consumed feed ad libitum and the intestines were full of digesta; thus, the high osmolarity could be result of active absorption of nutrients (Klasing et al., 2002). The differences between the results obtained from present study, with Klasing et al. (2002) is due to different species, strain, individual variations, levels of added betaine and ages in which osmolarity were determined. Most of the betaine was absorbed in the duodenum and jejunum with little left for absorption in ileum. Betaine is transported by the Na+ dependent amino acid transport system A and by the Na+ and Cl- dependent betaine- γ-amino butyric acid (GABA) transporter (Klasing et al., 2002). Kettunen et al. (2001) reported the presence of Na+ dependent active transport system for betaine in the duodenum and jejunum of broiler chicks (Klasing et al., 2002). They also found that supplementation of betaine to the diet increased the Na+ dependent component in betaine uptake as well as the total quantity taken up by the duodenum (Klasing et al., 2002). The duodenum also had the highest osmolarity and because this tissue uses betaine for protection against a hyperosmotic environment (Klasing et al., 2002).
Effect of TDS: The effect of TDS was not significant on percentage of PCV although comparing amount of PCV in chicks consumed second and third levels of TDS with the control group showed 3.05 and 9.49% increment, respectively. Results showed an increasing effect of TDS on water consumption (at 28 and 42 days) and excreta moisture at 28 days (p<0.05) and also an increment in plasma osmolarity at 28 days (p>0.05). Decrease in blood volume is a powerful stimulus for rennin secretion. Thus, plasma angiotensin II (ANG II) concentration increased after consuming high levels of salinity. These results indicate that an increase in plasma osmolarity, a decrease in blood volume and ANG II are involved in the regulation of drinking. The osmotic, volaemic and ANG II stimuli are also dipsogenic in truly terrestrial species such as birds. In these truly terrestrial animals, cellular dehydration of osmosensitive neurons in the hypothalamus caused by osmotic stimuli is the strongest stimulus to elicit drinking. The potent osmotic effect is assessed by the fact that vigorous drinking occurs after the stimulus even through the other two dipsogenic stimuli, the volaemic stimulus and ANG II, are suppressed because of a simultaneous increase in blood volume. By contrast, hypo volaemia is only a week dipsogenic stimulus in birds even though plasma concentration of ANG II, which is it by itself a strong stimulus for drinking (Hazon and Flik, 2002). Obtained results indicated that consumed saline water caused hyperosmolarity condition (betainexTDS: 0x375, 0x1375 and 0x2375 in Table 4 for three parts of intestine).The increasing effect of TDS and interaction effect of TDS and betaine on the blood osmolarity at 28 days but not 42 days of age is also imply that young chicks are more susceptible to salt than older birds. Furthermore, because provided levels of TDS weren’t marginally limiting, in this research, blood osmolarity was in the ranges which were reported by Freeman (1983), Kettunen et al. (2001) and Klasing et al. (2002) for plasma osmolarity in normal chicks.
CONCLUSIONS
Betaine may be involved in the protection of intestinal epithelia against osmotic disturbance which can be caused by saline water but further researches is needed to investigate with higher levels of water salinity.
ACKNOWLEDGMENTS
We wish to thank Professor K.C. Klasing and Professor M. Eklund for their valuable comments. We also thank Dr. H. Mahravani Head of FMD Department of Razi Vaccine and Serum institute and A. Kamalzare the expert of this department for their technical help.
REFERENCES
- Eklund, M., E. Bauer, J. Wamatu and R. Mosenthin, 2005. Potential nutritional and physiological functions of betaine in livestock. Nutr. Res. Rev., 18: 31-48.
Direct Link - Kettunen, H., S. Peuranen and K. Tiihonen, 2001. Betaine aids in the osmoregulation of duodenal epithelium of broiler chicks and affects the movement of water across the small intestinal epithelium in vitro. Comp. Biochem. Physiol., 129: 595-603.
Direct Link - Klasing, K.C., K.L. Adler, J.C. Remus and C.C. Calvert, 2002. Dietary betaine increases intraepithelial lymphocytes in the doudenum of coccidia infected chicks and increases functional properties of phagocytes. J. Nutr., 132: 2274-2282.
Direct Link - Maiorka, A., N. Magro, H. Bartles, A. Kessler and J. Penz, 2004. Different sodium levels and electrolyte balances in Pre-starter diets for broilers. Braz. J. Poult. Sci., 6: 143-146.
Direct Link - Mirsalimi, S.M., P. O'Brien and R. Julian, 1992. Blood volume increase in salt-induced pulmonary Hypertension, heart failure and ascites in broiler and white leghorn chickens. Can. J. Vet. Res., 57: 110-113.
Direct Link