
H. H. Musa
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
J.H. Cheng
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
X.S. Wu
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
H.P. Ju
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
D.M. Mekki
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
G.H. Chen
College of Animal Science and Technology, Yangzhou University, Yangzhou, 225009, China
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 4 | Page No.: 693-696







ABSTRACT
Present study was focus to compare LDL receptor mRNA expression, total cholesterol, triglyceride, lipoprotien levels and abdominal fat weight in genetically fat and lean chickens. Genetically lean (Rugao) and fat (Anka) chickens were reared in the same environmental condition, at 12 weeks of age samples of liver tissue were collected and abdominal fat weight was determined. Similarly, total cholesterol, triglycerides and high density lipoprotein were assayed using a commercial enzymatic kit, very low density lipoprotein and low density lipoprotein were estimated using the Friedewald equation. Total RNA from liver tissues were isolated using the standard Trizol methods and then total RNA was reverse transcribed by moloney murine Leukemia virus. Semi-quantitative RT-PCR was developed to quantify the levels of LDL receptor mRNA expression. The level of LDL receptor expression was significantly (p< 0.05) difference between lean and fat chicken. In addition, lean and fat chickens were significantly differed on triglyceride, very low density lipoprotein and abdominal fat weight. The expression of LDL receptor mRNA in liver of fat chicken was negatively correlated with abdominal fat weight. However, in lean chicken was negatively correlated with total cholesterol, triglyceride, lipoprotein concentration, abdominal fat weight and percentage of abdominal fat weight. In addition, within two breeds LDL receptor mRNA expression in liver was negatively correlated with low density lipoprotein, abdominal fat weight and percentage of abdominal fat weight.
PDF Abstract XML References Citation
How to cite this article
H. H. Musa, J.H. Cheng, X.S. Wu, H.P. Ju, D.M. Mekki and G.H. Chen, 2007. Analysis of LDL Receptor mRNA Expression, Serum Biochemical and Abdominal Fat Weight in Fat and Lean Chickens. Journal of Biological Sciences, 7: 693-696.
DOI: 10.3923/jbs.2007.693.696
URL: https://scialert.net/abstract/?doi=jbs.2007.693.696
DOI: 10.3923/jbs.2007.693.696
URL: https://scialert.net/abstract/?doi=jbs.2007.693.696
INTRODUCTION
The Low Density Lipoprotein Receptor (LDLR) is the first member of a still increasing family of structurally and functionally related cell surface receptors reviewed by (Nykjaer and Willnow, 2002). The fact that LDL receptor relatives can be found in species ranging from nematodes to insects and mammals indicates that the genes have originated from an evolutionary ancestor gene. The low density lipoprotein receptor is the key component in the feed-back regulated maintenance of cholesterol homeostasis in the body. The LDL receptor family consists of cell-surface receptors that recognize extracellular ligands and internalize them for degradation by lysosomes (Bajari et al., 2005). The LDL receptor is the prototype of this family, which also contains Very Low Density Lipoprotein Receptors (VLDL-R), apolipoprotein E receptor 2, LRP and megalin. The primary legends for the receptor is low density lipoprotein, or LDL which contains a single copy of apolipoprotein B-100; approximately 65-70% of plasma cholesterol in human circulates in form of LDL (Hyesung and Stephen, 2003). The family members contain four major structural modules: The cysteine-rich complement-type repeats, epidermal growth factor precursor-like repeats, a transmembrane domain and a cytoplasmic domain. Each structural module serves distinct and important functions. These receptors bind several structurally dissimilar ligands. It is proposed that instead of a primary sequence, positive electrostatic potential in different ligands constitutes a receptor binding domain. This family of receptors plays crucial roles in various physiologic functions. Due to the lack of studies of this gene in chicken present study was focus to compare LDL receptor mRNA expression, total cholesterol, triglyceride, lipoprotein levels and abdominal fat weight in genetically fat and lean chickens and to study the relationship between LDL receptor expression, serum biochemical and abdominal fat weight.
MATERIALS AND METHODS
Animals and tissue collection: Genetically lean (Rugao) and fat (Anka) chickens were from Jiangsu Poultry Institute (Yangzhou, China). Birds were reared in floor pens in a light-tight facility, feed and water were provided ad-libitum at all times during the study periods from December to March 2006. At 12 weeks of age the birds were fasted for overnight and slaughter, approximately 1.5 g of liver tissue was collected from each breed in 1.5 mL microfuge tube. The excised specimens were immediately snap frozen in liquid nitrogen and transferred to college of animal Science and Technology, Yangzhou University; stored at 80°C until RNA extraction. Similarly, carcasses were eviscerated and dissected manually to determine abdominal fat weight (Musa et al., 2006).
Serum biochemical analysis: Five milliliter blood samples were taken from the wing vein of fasting chicken, serum was harvested by centrifugation at 3000 rpm for 10 min and then serum was frozen and stored at 20°C for future analysis. Total serum cholesterol and triglycerides were assayed using a commercial enzymatic kit supplied by (Zhe jiang Dongou Biological Engineering Co., Ltd.). High-density lipoprotein cholesterol was detected enzymatically after precipitation of (LDL and VLDL) by heparin and manganese. Very low density lipoprotein cholesterol and low density lipoprotein cholesterol were estimated using the Friedewald equation (Friedewald et al., 1972).
RNA extraction: Total RNA was isolated from liver tissue using the standard Trizol reagent methods according to the manufacturer’s protocol (Invitrogen/Life Technologies, Carlsbad, CA) (Chomczynski, 1993). Briefly 50-100 mg of tissues was homogenized with 1 mL Trizol. The homogenized samples were extracted with chloroform and then precipitated by isopropyl alcohol. The resultant RNA pellets were washed once with 75% ethanol and the dried RNA pellets were dissolved in 30 μL of 100% formamide deionzed.
RNA integrity and purity: The RNA samples were resuspended into 100% formamide and quantified fluorimeterically at 260 nm. All RNA isolates had an OD260:OD280 between 1.8 and 2.0 were selected for this study. RNA quality or integrity was checked by 1% agarose gel electrophoresis, stained with 1 μg mL-1 gold view. Samples were adjusted by 100% formamide deionzed to a concentration of 100 ng μL-1 and stored in 20°C.
Primer design: Low density lipoprotein receptor messenger RNA primer was designed by Oligo 6.0 software, according to chicken genomic sequence in Gen Bank database (accession number AJ515243). Forward was 5CGC GTC CGG CTC CAT ATC3 and reverse was 5CTC GCA GCC CCA CTC ATC C3. In addition one pair of chicken β-actin primer was used forward was 5TGC CAG GGT ACA TTG TGG TA3 and reverse was 5 TGC GTG ACA TCA AGG AGA AG3.
First strand cDNA synthesis: The cDNA was synthesized using first strand cDNA synthesis kits supplied by (TakaRa Biotechnology Dalian Co., Ltd.). Briefly 100 ng of total RNA was reverse transcribed using 10 pmol μL-1 of oligo (dT) 18 primer. The final reaction volume was 20 μL-1, containing 5x M-MLV buffer, 10 mM of dNTP Mixture, 40 U μL-1 of RNAse inhibitor, 200 U μL-1 RTase M-MLV (RNAse H-) and RNAse free water. The cDNA was synthesis at 42°C for 1 h and extended at 70°C for 15 min, thereafter cDNA stock was stored at 20°C. The yield of cDNA was measured according to the PCR signal generated from the internal standard house-keeping gene β-actin amplified under 25 μL reaction volume contains 2.5 μL PCR buffer, dNTP 2 μL, Tag DNA polymerase 0.2, 10 pmol of each β-actin primer 1, 1.5 μL cDNA template and 16.8 μL sterilized water. The reaction condition was 94C for 5 min, 30cycles of 94°C for 30 sec, 57.5°C for 30 sec and 72°C for 60 sec and finally extended for 10 min at 72°C and then PCR products were separated on 1% agarose gel.
RT-PCR analysis of LDL receptor mRNA expression in liver: Semi-quantitative RT-PCR for LDL receptor primer and chicken beta actin primer were carried out in a total volume of 25 μL of solution containing 2.5 μL of 1xPCR reaction buffer, 2 μL of (each 2.5 μ mol) dNTP mix, 0.5 U of Taq DNA polymerase, 5pmol of each gene specific primers and 5 pmol each of the primers specific for β-actin 1, 1.6 μL cDNA template and 14.6 μL sterilized water. The amplification was performed in Biometra PCR System (Applied Biosystems). Thermal cycling parameters were as follows: 1 cycle 94°C for 5 min, followed by 35 cycles, 94°C for 30 sec, 64.5°C for 30 sec, 72°C for 1 min with a final extension at 72°C for 10 min., the samples were then run on the same agarose gel (1%) for quantitation.
RNA quantification: Images of the RT-PCR gold view stained agarose gels were acquired with GENE-GENIUS (Bio-imaging system) and quantification of the bands was performed by Image Quant TL (Amersham Biosciences, USA). Band intensity was expressed as relative absorbance units. The ratio between the sample RNA to be determined and β-actin was calculated to normalize for initial variations in sample concentration and as a control for reaction efficiency.
Statistical analysis: Values are presented as the mean and standard error of mean, significant differences were obtained using a paired Student's t-test (Steel and Torrie, 1980). The relation between LDL receptor mRNA expressions, lipoprotein concentration and abdominal fat weight was determined using Pearson bivariate coefficient correlation, performed by SPSS software.
RESULTS AND DISCUSSION
Semi-quantitative RT-PCR: A 1.6 μL CDNA were used in semi-quantitative RT-PCR with LDL receptor primer and Chicken β-actin as internal control (Fig. 1). The image acquired with GENE-GENIUS was quantified by Image Quant TL; band intensity was expressed as relative absorbance unit. Semi-quantitative RT-PCR is the most sensitive method for detecting gene transcription products; however, it can be highly variable and may not accurately reflect gene activity. In each quantitative RT-PCR based methods, specific errors will be introduced due to minor differences in the starting amount of RNA or differences in efficiency of cDNA synthesis and PCR reaction. Therefore, a reliable quantitative RT-PCR method required normalization for experimental variations by using an endogenous control (Dheda et al., 2004). Housekeeping genes, which are expected to be expressed at a constant level and should not be affected by experimental conditions, are most often used as control for variation of input RNA and an efficiency of reverse transcription. The liver is a key element in the control of plasma cholesterol, which is largely determined by the rate of removal of Low Density Lipoproteins (LDLs) from the circulation, regulated by hepatic LDL receptors (Myant, 1990; Brown and Goldstein, 1986).
Comparison and correlation analysis of LDL receptor expression, serum biochemical and fat weight in fat and lean chickens: The level of LDL receptor expression in this study was significantly (p<0.05) difference between lean and fat chicken.
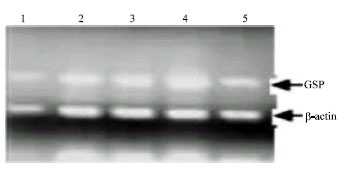 | |
| Fig. 1: | Semi-quantitative RT-PCR analysis of LDL receptor mRNA expression, 1, 2 and 3 are for fat chicken and 4 and 5 are for lean chicken; GSP, gene specific primer of LDL receptor gene; β-actin is chicken beta actin |
In addition, lean and fat chickens were significantly differed on triglyceride, very low density lipoprotein and abdominal fat weight (Table 1). The benefits from LDL receptor mediated regulatory system are the coordination of utilization of intra-and extracellular sources of cholesterol at systemic level (Brown and Goldstein, 1986). Because mRNA levels do not always correlate directly with the amount of functional protein produced, determining the functional relationship between LDL receptor and serum biochemical is one way to gauge the relevance of the expression data to actual physiological and biochemical effects. In this study expression of LDL receptor in liver of fat chicken was negatively correlated with abdominal fat weight. However, in lean chicken was negatively correlated with total cholesterol, triglyceride, lipoprotein concentration, abdominal fat weight and percentage of abdominal fat weight. In addition, within breeds LDL receptor mRNA expression in liver was negatively correlated with low density lipoprotein, abdominal fat weight and percentage of abdominal fat weight (Table 2). Elevated serum LDL-cholesterol concentrations in the general population result, in part, from a high intake of saturated fats and cholesterol. The potential effect of dietary cholesterol to increase LDL concentrations is offset by the inhibition of hepatic cholesterol synthesis. The saturated fatty acids from the chylomicron remnants, or those that are eventually mobilized from adipose tissue, appear to down regulate the LDL receptor, leading to an increase in LDL concentrations. Richards et al. (2003) found significant (p<0.05) correlation among the expression levels of various lipogenic genes.
| Table 1: | Comparison between fat and lean chickens on LDL receptor mRNA expression, serum biochemical and abdominal fat weight |
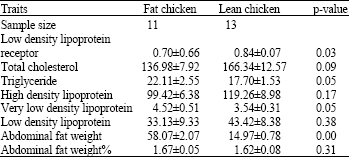 | |
Data was presented as means and standard error of means; P value is significant at (p<0.05) | |
| Table 2: | Relation between LDL receptor mRNA expression, cholesterol, triglyceride, lipoprotein and abdominal fat weight in genetically fat and lean chickens |
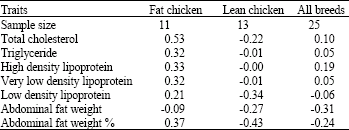 | |
Table explains the correlation between the level of LDL receptor mRNA expression in liver and levels of serum biochemical and abdominal fat weight of genetically fat and lean chicken. Correlation is significant at (p<0.05) | |
In addition, LPL expression was positively correlated with all of the major lipogenic enzyme genes, possibly indicating a functional coordination in the expression of this gene affecting lipid deposition in adipose tissue with those genes regulating hepatic lipid production (Richards et al., 2003). The expression of mRNA for LDL receptor and VLDL receptor were correlated. In addition, the expressions of both receptors were negatively correlated with cholesterol concentration in follicular fluid (Argov et al., 2004). On the other hand, insulin is a potent stimulator of LDL receptor expression (Wade et al., 1988) which is the primary mechanism by which chylomicron remnants are cleared in vivo (Havel, 1995). In contrast to a defect in lipolysis, decreased LDL receptor expression would lead to the accumulation of smaller, denser remnant lipoproteins. Although a number of studies have provided evidence for a positive association between central obesity, insulin resistance and glucose metabolism, it is less clear whether and by what mechanism insulin per se regulates the metabolism chylomicrons in vivo (Sattar et al., 1998). Finally from the comparison analysis we can conclude that the levels of LDL receptor mRNA expression can be consider as a candidate for triglyceride, very low density lipoprotein and abdominal fat weight in chickens. In addition, correlation analysis shows that decrease in LDL receptor expression was associated with increased abdominal fat weight in fat chicken. However, increase in LDL receptor expression was associated with decreased serum biochemical levels and abdominal fat weight.
REFERENCES
- Argov, N., U. Moallem and D. Sklan, 2004. Lipid transport in the developing bovine follicle: Messenger RNA expression increases for selective uptake receptors and decreases for endocytosis receptors. Biol. Reprod., 71: 479-485.
Direct Link - Bajari, T.M., V. Strasser, J. Nimpf and W.J. Schneider, 2005. LDL receptor family: Isolation, production and ligand binding analysis. Methods, 36: 109-116.
Direct Link - Chomczynski, P., 1993. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques, 15: 532-537.
PubMed - Dheda, K., J.F. Huggett, S.A. Bustin, M.A. Johnson, G. Rook and A. Zumla, 2004. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques, 37: 112-119.
Direct Link - Friedewald, W.T., R.I. Levy and D.S. Fredrickson, 1972. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem., 18: 499-502.
CrossRefPubMedDirect Link - Havel, R.J., 1995. Chylomicron remnants: Hepatic receptors and metabolism. Curr. Opin. Lipidol., 6: 312-316.
Direct Link - Hyesung, J. and C.B. Stephen, 2003. An Intramolecular Spin of the LDL receptor b-propeller. Structure, 11: 133-136.
Direct Link - Musa, H.H., G.H. Chen, J.H. Cheng, B.C. Li and D.M. Mekki, 2006. Study on carcass characteristics of chicken breeds raised under the intensive condition. Int. J. Poult. Sci., 5: 530-533.
CrossRefDirect Link - Nykjaer, A. and T.E. Willnow, 2002. The low-density lipoprotein receptor gene family: A cellular Swiss army knife. Trends Cell Biol., 12: 273-280.
Direct Link - Richards, M.P., S.M., Poch, C.N. Coon, R.W. Rosebrough, C.M. Ashwell and J.P. McMurtry, 2003. Feed restriction significantly alters lipogenic gene expression in broiler chicken. J. Nutr., 133: 707-715.
Direct Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Brown, M.S. and J.L. Goldstein, 1986. A receptor-mediated pathway for cholesterol homeostasis. Science, 232: 34-47.
CrossRefDirect Link