
G. E. Eriyamremu
Department of Biochemistry, University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
S.O. Asagba
Department of Biochemistry, Delta State University, P.M.B. 1, Abraka, Delta State, Nigeria
S.O. Uanseoje
Department of Biochemistry, University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
S.E. Omeregie
Department of Biochemistry, University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
C.O. Omofoma
Department of Medical Biochemistry, Faculty of Basic Medical Sciences,
University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 3 | Page No.: 526-531







ABSTRACT
The purpose of the present study was to evaluate the effect of Nigerian-Like Diet (NLD) and Western-Like Diets (WLD) on colonic mucosa lipid peroxidation, stress enzymes and nuclear membrane ATPase in the early stages of colon carcinogenesis in rats. Sixty male Wistar rats were grouped into three groups of 20 animals each; one group was fed with a Normal Diet (ND) and acted as the diet control. One group was fed with the NLD and the last group was fed with a WLD for 12 weeks. In each diet class, half of the animals were fed with the diet alone and the other half was fed with the diet containing cycas and after six weeks, a mid term study was carried out. Mid-term and full-term studies show a significant (p< 0.05) increase in nuclear ATPase activity of the rats fed with the WLD and a decrease in those fed the NLD compared with control. Feeding rats with the NLD significantly (p< 0.05) decreased the level of colonic mucosa lipid peroxidation compared with the WLD. A significant rise in the activity of Superoxide Dismutase (SOD) was observed in the rats fed the ND and NLD, while in the rats fed the WLD, SOD activity was compromised in both the mid- and full-term studies. The study shows that a NLD and WLD fed to rats alter mucosa plasma membrane lipid peroxidation, SOD and nuclear membrane ATPase in ways which suggest that the NLD may protect rats from colon carcinogenesis.
PDF Abstract XML References Citation
How to cite this article
G. E. Eriyamremu, S.O. Asagba, S.O. Uanseoje, S.E. Omeregie and C.O. Omofoma, 2007. Colonic Lipid Peroxidation, Nuclear Membrane ATPase and Stress Enzymes in Rats Fed a Nigerian-Like Diet and Cycas. Journal of Biological Sciences, 7: 526-531.
DOI: 10.3923/jbs.2007.526.531
URL: https://scialert.net/abstract/?doi=jbs.2007.526.531
DOI: 10.3923/jbs.2007.526.531
URL: https://scialert.net/abstract/?doi=jbs.2007.526.531
INTRODUCTION
Colon cancer is frequent in Western Europe, North America and Australia (Greelee et al., 2001) where the major risk factors involved in the development of the cancer is of genetic and dietary origin. Sporadic colorectal cancer represents about 95% of all cases of the disease and is mostly associated with dietary risk factors (Boutron-Ruault, 2002). Causal associations obtained in epidemiological studies between colon cancer development and diet is controversial and still subject to debate, however early observational studies showed that colorectal cancer risk is lower in African populations consuming a diet high in carbohydrates and in fibre than in Europeans who consume diets low in such foods (Burkitt, 1984). Exposure to chemical carcinogens present in roasted meats, are now widely accepted as risk factors for colorectal cancer (Lang et al., 1994), while diets high in carbohydrates and dietary fibre are generally accepted to prevent the disease (Eriyamremu and Adamson, 1994).
Frequently, the initiation of a cell to a cancerous one is induced by genotoxic carcinogens that directly damage the DNA (Shacter and Weitzman 2002; Pool-Zobel and Leucht, 1997). In mammalian cells, oxidative stress, resulting from a prevalence of oxidants, produces specific types of cellular damage to macromolecules (Toyokuni et al., 1995). The damage by oxidants and by other genotoxic agents is expected to play a role in the development and generation of cancer (Lindblom, 2001; Shih et al., 2001). Earlier studies have demonstrated the production of extracellular superoxide (O2¯) by the intestinal microflora as a source for generating these oxidants that can cause chromosomal instability and colon cancer (Huycke et al., 2002). The extent of damage occasioned by these oxidants is dependent on the activities of detoxifying enzymes such as superoxide dismutase and catalase. Earlier studies in rodents have demonstrated a reduction in the activities of these antioxidant enzymes (Remacle et al., 1992) in cancers.
There is general consensus that colon carcinogenesis arise from instability of the colonic tissue which leads to the development of polyps and then tumour. The mammalian hind gut is highly active and the contents of its microflora depend to a large extent on the diet pattern of the individual and changes in response to gross nutritional shifts. Earlier studies have shown that the hind gut microflora can alter with diet (Eriyamremu and Adamson, 1995; Mitsuoka, 1996). With the increasing, westernization of the Nigerian population, there is a shift from the traditional diets rich in carbohydrates and fibre to a more Western one which is rich in fat and protein. These changes in the diet pattern can result in changes in the colon microflora and would have consequences for the colon. Hence there is continuous interest on the influence of diet on the hindgut microflora, its influence on the colonic tissue and its implication for colon carcinogenesis. However there is scarcity of information on the influence of wholly compounded Nigerian diet on colonic membrane lipid peroxidation. This report is on the influence of a Nigerian-like diet on lipid peroxidation and some antioxidant enzymes in the colonic mucosa at early stages of carcinogenesis.
MATERIALS AND METHODS
Experimental design and collection of tissues: Sixty male Wistar albino rats (6 weeks old) from the same stock weighing 45-55 g were used in this experiment. The animals were obtained from the Department of Pharmacology, of the University of Benin, Benin City, Nigeria, at least one month before the commencement of the experiment in our laboratory during which time they were fed with rat chow. The animals were divided into three diet class of 20 rats per class. One class received the Normal Diet (ND), another class received a high carbohydrate and high fibre diet (Nigerian-Like Diet, NLD) and the third class received a high protein and high fat, Western-Like Diet (WLD) (Table 1). Each diet class was further sub-divided into two groups of 10 rats each; with one group fed on the diet, while the other was first tube fed 5 g kg-1 body weight cycas before been allowed free assess to the diet. The animals were grouped such that the weight difference between all the groups was about 0.1 g. Before the commencement of the study period the animals were first acclimatized for two weeks with their respective experimental diet. The study period lasted for twelve weeks during which time the animals were allowed water and the diets ad libitum. After six weeks, half of the animals in each group (5 animals) were sacrificed for a mid-term study of the parameters studied. Food intake and dry faecal output were measured daily and weight gain data were recorded weekly. In all experiments, animals were fasted overnight before being killed. All these animal treatment were carried out in accordance with the principles of laboratory animal care of the NIN guide for Laboratory Animal Welfare as contained in the NIN Guide for Grants and Contracts, vol. (1985).
| Table 1: | Composition of the study diets |
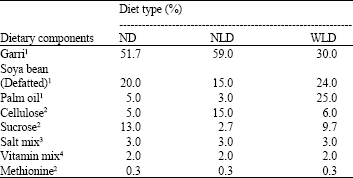 | |
1From a local market in Benin City. 2Product from Merck, Darmstadt, Germany. 3Product from Tuco Products Company, Ontario, Canada. 4AIN-93-VX vitamin mix recommended for use with the AIN-93G and AIN-93M diet formulations Garri is a cassava (Manihot esculentum) | |
Colonic preparation: The colon (after the first 10 cm, then the next 10 cm) was removed and used immediately for isolation of cells. The colon was slit open and rinsed thoroughly with cold normal saline and the mucosa was removed by scraping with glass slides. The colonic tissue was immediately homogenized and centrifuged at 5000 g for 10 min under cold conditions, while the residue was recovered for ATPase assay, the supernatant was used to analyze for Superoxide dismutase (SOD), catalase and lipid peroxidation.
Preparation of nuclear membranes: Colonic nuclear membranes were prepared by extensive digestion of nuclei of the colonic mucosa homogenate with DNase I (Widnell and Tata, 1964). The nuclei were used to prepare nuclear membranes by modifications of the procedure of (Kay et al., 1972).
ATPase, superoxide dismutase and catalase assays: The nuclear ATPase reaction was carried out according to Agguter and McCaldin (1978) and colonic Na+/K+ ATPase was estimated by the method of Adam-Vizi and Seregi (1982). The inorganic phosphate released in both the nuclear and colonic assays were determined (Fiske and Subarrow, 1925).
Superoxide Dismutase (SOD) activity of the samples was assayed according to the method described by Misra and Fridovich (1972). One unit of the enzyme was defined as the amount of the enzyme required for 50% inhibition of oxidation of epinephrine to adrenochrome in one minute.
Catalase (CAT) activity was assayed by the method of Kaplan and Groves (1992) and was determined as residual H2O2 after incubation with the enzyme.
The protein content of all the samples was estimated by the method of Lowry et al. (1951).
Membrane lipid peroxidation: Estimation of colonic membrane lipid peroxidation was by the method of Gutteridgre and Wilkins (1982). Values for thiobarbituric acid reactive substances (TBARS) are reported as malondialdehyde (MDA) and quantified using a Molar extinction coefficient of 1.5x105 M cm-1 and expressed as mmole MDA/g tissue weight.
Statistical analysis: The values are reported as Means±SEM. Statistical difference was determined using ANOVA and differences in the means were tested by Duncan’s multiple range tests (1969).
RESULTS
Statistical analysis reveal a significant increase in the weight gain of the rats which consumed the WLD compared with the rats which consumed the other diets, despite significantly low food consumption (Table 2). These animals fed with the WLD also excreted the least faeces. Cycas supplementation of the diets increased weight gain, food intake and dry faecal output. This trend was observed in the rats after six and twelve week’s exposure to the various diets. The rats fed with the NLD though gained the least weight, consumed the most food and excreted the most faeces compared with the other diet groups. The data suggest that the NLD diet induces weight loss and increases dry faecal output while the WLD diet increased weight and reduced dry faecal output. Cycas inclusion in the diets increased these data.
After six weeks of feeding, a statistical significantly increased activity of the ATPase of the rats fed with the WLD diet was observed when compared with the rats in other diet groups. In converse the NLD diet significantly decreased nuclear membrane ATPase when compared with the other diet groups. Inclusion of cycas in the experimental diets significantly increased the nuclear ATPase activity. A similar trend was observed after feeding the rats for twelve weeks (Table 3).
Feeding rats with the NLD significantly (p<0.05) increased the level of colonic lipid peroxidation compared with the normal diet (Table 4) Colonic lipid peroxidation was further significantly (p<0.05) increased by the WLD relative to the NLD and ND.
| Table 2: | Average weight gain, food intake and dry faecal output |
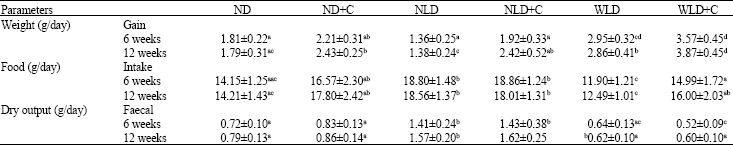 | |
Values are expressed as mean±SEM, n = 5. ND is the normal diet fed group, the HCF is the group fed with a high carbohydrate high fibre diet and HPF represent the group fed with a high protein and high fat diet. ND+C; HCF+C and HPF+C-Diets supplemented. with cycas. Means of the same row followed by different superscripts differ significantly (p<0.05) | |
| Table 3: | The level of colonic membrane lipid peroxidation and the activities of superoxide dismutase and catalase of rats |
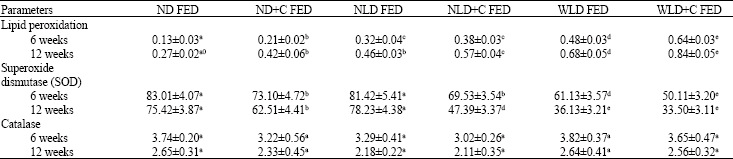 | |
Values are expressed as mean±SEM, n = 5. Means of the same row followed by different superscripts differ significantly (p<0.05). Lipid peroxidation is expressed as mmole MDA/g tissue. Catalase activity is expressed as μmoles/min/mg tissue. SOD activity is expressed as Units/g tissue. ND is the normal diet fed group, the HCF is the group fed with a high carbohydrate high fibre diet and HPF represent the group fed with a high protein and high fat diet. ND+C; HCF+C and HPF+C represents the respective diets supplemented with cycas | |
| Table 4: | Nuclear total ATPase activity of the colonic nuclear membrane of the rats |
 | |
Values are expressed as mean±SEM, n = 5. Means of the same row followed by different superscripts differ significantly (p<0.05). ATPase activity is represented as nmol Pi released/min/mg protein, ND is the normal diet fed group, the HCF is the group fed with a high carbohydrate high fibre diet and HPF represent the group fed with a high protein and high fat diet. ND+C; HCF+C and HPF+C represents the respective diets supplemented with cycas | |
The inclusion of cycas to all the experimental diets enhanced lipid peroxidation and these however reached a significant (p<0.05) level in the rats fed the normal diet and the WLD. The same trend was observed in the data taken after 12-weeks of feeding, although there was a significant (p<0.05) increase in lipid peroxidation in rats fed with the NLD+cycas relative to those fed NLD alone. There appears to be an inverse relationship between the data on SOD and the level of lipid peroxidation; the WLD lowered SOD activity compared with the NLD. Addition of cycas to both diets significantly reduced the activity of SOD after 6 and 12 weeks of feeding. Statistical analysis did not reveal any significant change in the activity of catalase in the colonic of the rats fed with the experimental diets compared with the control. This study reveals that colonic membrane lipid peroxidation and SOD activity is responsive to diet changes.
DISCUSSION
Cycas contain cycasin, which is modified by the colonic microflora to dimethylhydrazine (a potent carcinogen). The level of cycas plant fed to the rats and the duration of the feeding protocol in this study will relate to very early events in the initiation stage of colon carcinogenesis. Thus this study assessed the role of a NLD and a WLD on some biochemical indices in early colonic carcinogenesis. The biochemical events studied were colonic membrane lipid peroxidation, antioxidant enzymes and nuclear membrane ATPase.
As we have previously reported (Eriyamremu and Adamson, 1994; Eriyamremu and Adamson, 1995), the NLD which is high in carbohydrate and fibre decreased weight gain and increased dry faecal output (Table 2). Potential mechanisms by which dietary fibre can protect against the development of colorectal cancer is thought to include increases in stool bulk, dilution or binding of potential carcinogens and decrease in transit time (Burkitt, 1984). As the NLD increased faecal bulk, it would protect the rats from colorectal carcinogenesis. In contrast the WLD decreased dry faecal output which will increase transit time and concentrate potential carcinogens and thus predispose the rats to colorectal carcinogenesis. From the calculated metabolizable energy, the WLD contains more energy than the NLD and so animals consuming the WLD do not need to consume large quantities of the diet to meat their energy demand and this would have accounted for the low food intake observed in the animals on this diet.
The increased lipid peroxidation level in the rats fed with the NLD compared with the control diet (Table 3) agrees with those of an earlier study which also fed rats with a NLD (Asabga et al., 2004). The diet been low in protein but high in carbohydrate may induce protein malnutrition and this has been shown to cause oxidative stress (Tatli et al., 2000) and may enhance lipid peroxidation. The significantly increased lipid peroxidation in the rats fed with the WLD (high in protein and fat) compared with the NLD (Table 3) suggests that oxidative stress may not result from protein malnutrition alone. A high fat diet, as contained in the WLD, will increase enterohepatic circulation and thus improve the chances of loss of bile acids to the colon where they can be modified into secondary bile acids, some of which have been reported to increase lipid peroxidation (Sipos et al., 2002). n increase in lipid peroxidation by the WLD may be an important event in the role of this diet in early colon carcinogenesis.
Studies by Huycke et al. (2002) had shown that Enterococcus faecalis enhances the production of superoxide in the hind gut. As diet can alter the colonic microflora, a diets that favour the production of this bacterium would improve superoxide production in the colon. The increased lipid peroxidation observed in the rats fed with the WLD compared with the NLD may be a likely indication that the diet supports a microflora that can generate free radicals and thus cause high lipid peroxidation. Molecular pathways to colorectal cancer involve multiple genetic changes which may be acquired and/or inherited mutations or hypermethylation in cancer relevant genes. Persistent oxidative stress has been implicated to cause mutations in cancer related genes leading to increased proliferation and tumourigenesis (Zock and Katan, 1998). Thus the result of an elevated level of lipid peroxidation occasioned by the WLD would be a predisposition to mutation in cancer genes.
SOD is a metalloenzyme that play a vital rolein the protection of aerobic cells against oxygen free radical toxicity (Sun, 1990). Altered activities of SOD were shown to be important in multistage carcinogenesis of both rodents and humans. When compared to their appropriatenormal cell counterparts, tumor cells are almost always lowin SOD activity (Sun, 1990). This observation was made in a number of different cell types and is independent of the mechanism of cell transformation (McCormick et al., 1991; Sun et al., 1993). Furthermore, increased amounts of SOD were shown to be protective against cancer. Cycas inclusion in the various diets reduced total SOD activity (Table 3) and thus indicates that compromise in SOD activity is important for the onset of cycas induced colon carcinogenesis.
The observation that the WLD decreased SOD activity in rats (Table 3) is not surprising since the diet elaborated lipid peroxidation would have damaged colonic cells with its attendant loss of enzymes. Also diets high in fat have long been known to form mineral soaps with fatty acids in the intestine preventing their absorption. Heart MnSOD activity in rats is significantly reduced when fed diets deficient in manganese compared to those fed adequate or high dietary manganese (Davis et al., 1992). So if the fat in the WLD interfere with manganese, it may in part account for the observed low SOD of the rats fed this diet. So the NLD which decreased lipid peroxidation and increased SOD activity compared with the WLD may delay the onset of colon carcinogenesis.
Catalase is among the enzymes that help the cell to handle hydrogen peroxide though it should be noted that this enzyme is not the major route of hydrogen peroxide catabolism (Doroshow et al., 1986). This study did not observe any significant change in catalase activity in all the experimental groups (Table 3) which suggests that changes in catalase is not a likely manifestation or event in early colon carcinogenesis.
Several authors have reported the importance of the nuclear membrane in nucleocytoplasmic exchange of large macromolecules including proteins and RNA species (Ammouche et al., 1994) and these processes require ATP. One high energy phosphate bond is hydrolyzed to transport one nucleotide of mRNA (Clawson et al., 1980). Nuclear membrane ATPase is responsive to diet changes and thus alter functions central to the activity of the cell (Ammouche et al., 1994). The transport of RNA to the cytoplasm will thus affect activities such as protein synthesis and eventually cell proliferation. This study shows that the NLD reduced the nuclear membrane ATPase (Table 4) and would thus reduce nucleocytoplasmic exchange of RNA and may protect against colon cancer. On the contrary, the WLD increased the nuclear membrane enzyme activity and may support improved transport of RNA from the nucleus and thus promote colon carcinogenesis. The possible low nucleocytoplasmic exchange in the rats fed the NLD may help in part account for the observed low SOD activity in these rats compared with the ND fed ones. It should be expected that a possible high nucleocytoplasmic exchange in the rats fed the WLD should increase SOD activity, but its effect on lipid peroxidation and the possibility of its fat content interfering with the absorption of minerals important for the enzyme action may cancel out this effect.
In this study, we did not examine the luminal epithelium of the animals for the degree of lesions or dysplasia. Despite this drawback, this study provides data on the effects of a NLD and WLD on colonic lipid peroxidation, SOD and NTPase activities in early colon carcinogenesis. The study shows that the NLD compared with the WLD alter these biochemical events in ways which suggest that it is protective against colon carcinogenesis and that change in these parameters are some of the early biochemical events in colon carcinogenesis. A study of the microflora supported by the NLD and WLD and their relationship to the degree of colonic epithelium dysplasia would be desirable.
REFERENCES
- Asagba, S.O., G.E. Eriyamremu, M.A. Adaikpo and A. Ezeoma, 2004. Levels of lipid peroxidation, superoxide dismutase and Na+/K+ ATPase in some tissues of rats exposed to a Nigerian-like diet and cadmium. Biol. Trace Element Res., 100: 75-86.
Direct Link - Davis, C.D., T.L. Wolf and J.L. Greger, 1992. Varying levels of manganese and iron affect absorption and gut endogenous losses of manganese by rats. J. Nutr., 122: 1300-1308.
Direct Link - Eriyamremu, G.E. and I. Adamson, 1994. Early changes in energy metabolism in rats exposed to an acute level of deoxycholate and fed on a Nigerian-like diet. Ann. Nutr. Metab., 38: 174-183.
Direct Link - Eriyamremu, G.E. and I. Adamson, 1995. Alterations in rat colonic faeces exposed to an acute level of deoxycholate and fed on a Nigerian-like diet. Nutr. Res., 15: 869-870.
Direct Link - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Gutteridge, J.M.C. and S. Wilkins, 1980. Copper-dependent hydroxyl radical damage to ascorbic acid: Formation of a thiobarbituric acid-reactive product. FEBS Lett., 137: 327-330.
PubMed - Huycke, M.M., V. Abrams and D.R. Moore, 2002. Enterococcus faecalis produces extracellular superoxide and hydrogen peroxide that damages colonic epithelial cell DNA. Carcinogenesis, 23: 529-536.
CrossRefDirect Link - Kaplan, J.H. and J.N. Groves, 1992. Liver and blood cell catalase activity of tumour-bearing mice. Cancer Res., 32: 1190-1194.
PubMed - Lang, N.P., M.A. Butler, J. Massengell, M. Lawson,R.C. Stotts, M. Hauer-Jensen and F.F. Kadlubar, 1994. Rapid metabolic phenotypes for acetyltransferase and cytochrome P4501A2 and putative exposure to food-borne heterocyclic amines increases the risk of colorectal cancer or polyps. Cancer Epidemiol. Biomarkers Prev., 3: 675-682.
Direct Link - Lindblom, A., 2001. Different mechanisms in the tumorigenesis of proximal and distal colon cancers. Curr. Opin. Oncol., 13: 63-69.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - McCormick, M.L., T.D. Oberley, J.H. Elwell, L.W. Oberley, Y. Sun and J.J. Li, 1991. Superoxide dismutase and catalase levels in renal tumors and their autonomous variants in the Syrian hamster. Carcinogenesis, 12: 977-984.
Direct Link - Misra, H.P. and I. Fridovich, 1972. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem., 247: 3170-3175.
CrossRefPubMedDirect Link - Pool-Zobel, B.L. and U. Leucht, 1997. Induction of DNA damage in human colon cells derived from biopsies by suggested risk factors of colon cancer. Mutat. Res., 375: 105-116.
Direct Link - Remacle, J., D. Lambert, M. Raes, E. Pigeolet and C. Michiels, 1992. Importance of various anti-oxidant enzymes for cell stability: Confrontation between theoretical and experimental data. Biochem. J., 286: 41-46.
Direct Link - Shacter, E. and S.A. Weitzman, 2002. Chronic inflammation and cancer. Onchology, 16: 217-226.
Direct Link - Shih, I.M., W. Zhou, S.N. Goodman, C. Lengauer, K.W. Kinzler and B. Vogelstein, 2001. Evidence that genetic instability occurs at an early stage of colorectal tumorigenesis. Cancer Res., 61: 818-822.
Direct Link - Sun, Y., 1990. Free radicals, antioxidant enzymes and carcinogenesis. Free Radical Biol. Med., 8: 583-599.
CrossRefDirect Link - Sun, Y., Y. Li and L.W. Oberley, 1993. Superoxide dismutase activity during dimethylhydrazine colon carcinogenesis and the effect of cholic acid and indole. Free Radic. Res. Commun., 4: 299-309.
Direct Link - Tatli, M.M., H. Vural, A. Koc, M. Kosecik and A. Atas, 2000. Altered antioxidant status and increased Lipid peroxidation in marasmic children. Pediatr. Int., 42: 289-292.
Direct Link - Toyokuni, S., K. Okamoto, J. Yodoi and H. Hiai, 1995. Persistent oxidative stress in cancer. FEBS Lett., 358: 1-3.
PubMed - Zock, P.L. and M.B. Katan, 1998. Linoleic acid intake and cancer risk: A review and meta-analysis. Am. J. Clin. Nutr., 68: 142-153.
Direct Link