
C. Guruvayoorappan
Department of Immunology, Amala Cancer Research Centre, Amala Nagar,
Thrissur, Kerala State 680 555, India
Girija Kuttan
Department of Immunology, Amala Cancer Research Centre, Amala Nagar,
Thrissur, Kerala State 680 555, India
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 8 | Page No.: 1511-1519







ABSTRACT
The antiangiogenic activity of rutin was studied using in vivo as well as in vitro models. In vivo angiogenic activity was studied using B16F-10 melanoma cell-induced capillary formation in C57BL/6 mice. Administration of rutin significantly inhibited (43.35%) the number of tumor directed capillaries induced by injecting B16F-10 melanoma cells on the ventral side of C57BL/6 mice. Rutin at non-toxic concentrations (5-25 μg mL-1) inhibited the vessel sprouting in rat aortic ring assay. Moreover, rutin was able to inhibit the proliferation, migration and capillary-like tube formation of human endothelial cells, the key steps of angiogenesis. Present studies using Tumor Associated Macrophages (TAMs) revealed that rutin could suppress the expression and production of VEGF and IL-1β and stimulate the production of TNF-α. Hence, the observed antiangiogenic activity of rutin is related to the regulation of these cytokines and growth factors by TAMs.
PDF Abstract XML References Citation
How to cite this article
C. Guruvayoorappan and Girija Kuttan, 2007. Antiangiogenic Effect of Rutin and its Regulatory Effect on the Production of VEGF, IL-1β and TNF-α in Tumor Associated Macrophages. Journal of Biological Sciences, 7: 1511-1519.
DOI: 10.3923/jbs.2007.1511.1519
URL: https://scialert.net/abstract/?doi=jbs.2007.1511.1519
DOI: 10.3923/jbs.2007.1511.1519
URL: https://scialert.net/abstract/?doi=jbs.2007.1511.1519
INTRODUCTION
Angiogenesis is a complex process, which involves the interactions of several cell types and mediators to establish a specific microenvironment suitable for the formation of new capillaries from pre-existing vessels (Kerbel and Folkman, 2002). The process of forming new blood vessels from an existing vascular bed, normally involves a series of steps, which include endothelial cell activation, breakdown of the basement membrane, migration, proliferation and tube formation of the endothelial cell (Folkman and Shing, 1992). This is a key event that feeds tumor growth and cancer metastasis (Roy et al., 2002). It is widely accepted that angiogenesis is the result of a net balance between the activities exerted by positive and negative regulators. Inflammatory cells, namely monocytes/-macrophages, T lymphocytes and monocytes, fully participate in the angiogenic process by secreting pro-and anti-inflammatory cytokines that could control the angiogenic swith (Bergers and Benjamin, 2003; Lingen, 2001). Neoplastic tissues present an inflammatory infiltrate and is characterized by the presence of Tumor Associated Macrophages (TAMs) which have the ability to both positively and negatively regulate tumor growth (Mantovani et al., 1992); tumor cell cytostasis may be induced, but in contrast tumor cell survival may be promoted by release of angiogenic and mitogenic cytokines in the tumor microenvironment such as VEGF (Barbera-Guillem et al., 2002) and proinflammatory cytokines such as IL-1β and TNF-α (Kataki et al., 2002; Ghezzi et al., 1991; Naldini et al., 2001; Balkwill and Mantovani, 2001; Cheng and Chen, 2001).
Several polyphenolic compounds are recognized as cancer chemopreventive agents. Flavonoids are especially well known to suppress tumor cell growth via cell-cycle arrest and by induction of apoptosis in several tumor cell lines (Romero et al., 2002; Choi et al., 1998; Davis et al., 1998; Lian et al., 1998; Matsukawa et al., 1993). Moreover, flavonoids inhibit endothelial cell proliferation and angiogenesis in vitro, the latter is endothelial cell cultures on collagen gels (Fotsis et al., 1993). Rutin, a member of the flavone family that is present at high levels in many vegetables has been found to possess antitumor (Deschner et al., 1991), anti-inflammatory (Ihme et al., 1996) and antioxidant (Bombardelli and Morrazzoni, 1993; Oomah and Mazza, 1996; Metodiewa et al., 1997) properties. We have reported earlier the antimetastatic activity of rutin in C57BL/6 mouse models (Lata et al., 1995). However the antiangiogenic potential of rutin and its effect on tumor associated macrophages has not been revealed. In this study, we tested the inhibitory effect of rutin on proliferation, migration and tube formation of endothelial cells, the key steps of angiogenesis. Moreover, the effect of rutin on the expression of VEGF, IL-1β and TNF-α on tumor associated macrophages were also evaluated.
MATERIALS AND METHODS
The present study was carried out in the Department of Immunology, Amala Cancer Research Centre.
Reagents and chemicals: Rutin (Fig. 1) was purchased from Sigma Chemical Co. (St. Louis, MO). All chemicals used were of reagent or higher grade.
ELISA kits: ELISA kits for mouse IL-1β and TNF-α were purchased from Pierce Endogen (Rockford, USA). Mouse VEGF ELISA kits were purchased from R and D System (Minneapolis, MN).
Animals: C57BL/6 (5-6 weeks-old) male mice (20-25 g) were purchased from National Institute of Nutrition, Hyderabad, India. All mice were housed, cared for and used in strict accordance with the rules and regulations of Institutional Animal Ethics Committee, Government of India.
Cell lines: Human Umbilical Vein Endothelial Cells (HUVECs) were isolated from human umbilical cord veins by collagenase treatment as described previously (Jaffe et al., 1973). HUVECs were maintained in gelatin-coated 75 cm2 flasks using M199 (HiMedia, Mumbai, India), supplemented with 20% (v/v) FCS (Biological Industries, Israel), 100 units mL-1 Penicillin, 100 μg mL-1 Streptomycin, 500 ng mL-1 basic fibroblast growth factor (bFGF; Pepro Tech Inc. Rocky Hill, NJ) and 100 ng mL-1 VEGF (Pepro Tech Inc. Rocky Hill, NJ). B16F-10 mouse melanoma cells (National Centre for Cell Sciences, Pune, India) were maintained as a monolayer culture in DMEM (HiMedia, Mumbai, India) supplemented with 10% (v/v) FCS and antibiotics. All the cell lines were maintained at 37°C in a humidified atmosphere containing 5% CO2.
Isolation and culture of Tumor Associated Macrophages (TAMs): Solid tumors (3-4 weeks old) were dissected and mashed aseptically in RPMI-1640 and the cell suspension was kept undisturbed for 2 h.
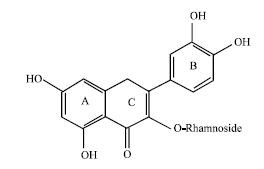 | |
| Fig. 1: | Chemical structure of rutin |
The supernatant was discarded and the adhered macrophages were scraped out using rubber policeman and cultured in RPMI-1640 supplemented with 10% (v/v) FCS and antibiotics.
In vivo angiogenesis assay: Male C57BL/6 mice were divided into two groups (6 animals group-1). Angiogenesis was induced by injecting B16F-10 (1x106) cells subcutaneously on the shaved ventral side of the mice. One group was kept as untreated control and the other group was treated with rutin (200 μM kg-1 B.wt.) for five consecutive days. After 9 days, animals were euthanized by cervical dislocation, ventral skin cut removed and the number of tumor directed capillaries per cm2 around the tumor was counted manually under dissection microscope (10x magnification).
In vitro angiogenesis
Rat aortic ring assay: Rat aorta, collected from 8-10 weeks old male Sprague Dawley rats were cut into 1 mm long cross section and were cultured in collagen coated Twenty-four well tissue culture plates in the presence of conditioned medium (100 μL) and escalating concentrations of rutin (5-25 μg mL-1) of rutin for six days. The number of microvessels sprouted from the aortic ring were counted manually under phase contrast microscope (40x magnification).
Endothelial cell proliferation assay: Endothelial cells (5x104 cells) were cultured for 24 h in the presence of escalating concentrations of rutin (5-25 μg mL-1) and the proliferation of endothelial cells were determined by the [3H]-thymidine incorporation assay (Lee et al., 1999). The recovered radioactivity was measured using Rack Beta Liquid Scintillation Counter (Wallac 1209, Pharmacia, Sweden) and expressed as counts per minute (cpm).
Wound migration assay: Endothelial cells were grown to confluence in 96-well collagen coated culture plates. The endothelial monolayers were wounded using a pipette tip. The media and dislodged cells were aspirated and the wounded cells were incubated in the test medium (M199 + escalating concentrations of rutin). After 48 h incubation, the cells were stained with crystal violet (0.1%) and cells migrated to the wounded area were counted under phase contrast microscope (40x magnification).
Tube formation assay: Endothelial cell tube formation was performed by Matrigel Assay (Gupta et al., 2002). Endothelial cells (5x104 cells) were cultured for 48 h in Matrigel (Sigma, St. Louis, USA) coated wells in the presence of escalating concentrations of rutin (5-25 μg mL-1). The cells were fixed with formaldehyde (5%), stained with Diff Quick (Dade Behring, Newark, DE) and photographed under phase contrast microscope (40x magnification).
| Table 1: | Results of PCR analysis using specific primers for VEGF, 1L-1B, TNF-α and GAPDH |
 | |
Quantification of VEGF, IL-β and TNF-α levels: Tumor associated macrophages (5x104 cells) were cultured for 24 h in the presence of escalating concentrations of rutin (5-25 μg mL-1) and the VEGF, IL-1β and TNF-α levels in the culture supernatant (0.1 mL) were quantified using ELISA kits as per the manufacturer’s instructions. Values are expressed in picogram mL-1.
Gene expression studies: Tumor associated macrophages (5x104 cells) were cultured in the presence of escalating concentrations of rutin (5-25 μg mL-1) for 24 h and the cDNA synthesis was carried out using cDNATM II kit (Ambion) as per the manufacturer’s instructions. PCR was performed using specific primers for VEGF, IL-1β, TNF-α and GAPDH (Table 1). The PCR products were electrophoresed on a 1.8% agarose gel and stained with ethidium bromide and photographed under UV light.
Bioassay for TNF-α: L929 cells (5000 cells well-1) were cultured for 48 h in the presence of TAM culture supernatant (0.1 mL) from the above mentioned experiment. The cells were fixed with formaldehyde (5%), stained with crystal violet (1%) and photographed under phase contrast microscope (40x magnification).
Statistical analysis: The data are expressed as mean±SD. Statistical comparisons were performed by one way analysis of variance (ANOVA) followed by Dunnett’s test. Unpaired Student t-test was used for statistical analysis, when only two groups are compared. The results were considered statistically significant if the p-values were less than 0.05.
RESULTS
Inhibition of angiogenesis by rutin in vivo: Control animals had an average number of 39.9±1.5 capillaries around the tumor. Rutin treatment showed a significant inhibition (43.35%) in the formation of tumor directed capillaries (Fig. 2).
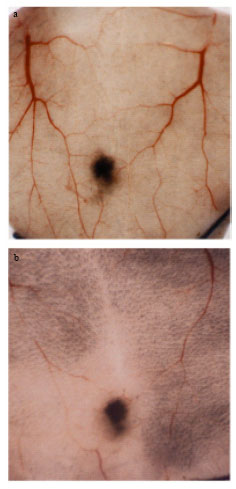 | |
| Fig. 2: | Inhibition of tumor directed capillaries by rutin in vivo. B16F-10 (106 cells/mice) were injected s.c. on shaved ventral side of C57BL/6 mice and treated simultaneously with 200 μM kg-1 b.wt. for five consecutive days. The tumor directed capillaries were counted at day 9 after implantation of B16F-10 cells. (a) control and (b) Rutin at 200 μM kg-1 b.wt. |
Effect of rutin on the inhibition of microvessel sprouting: Incubation of aortic rings with rutin showed a significant and dose dependent inhibition of microvessel outgrowth from the rat aorta ring induced by the conditioned medium from B16F-10 cells (Fig. 3), with 52.8% inhibition in microvessel out growth at a concentration of 25 μg mL-1.
Effect of rutin on endothelial cell proliferation: Rutin treatment showed a significant inhibition of endothelial proliferation. Rutin at a concentration of 25 μg mL-1 showed 41.02% inhibition in the proliferation of endothelial cells (Fig. 4).
Effect of rutin on endothelial cell migration: Rutin treatment showed a significant and concentration-dependent inhibition of endothelial cell migration. Rutin at a concentration of 25 μg mL-1, showed 87.5% inhibition of endothelial cell migration. At 5 and 10 μg mL-1, the percentage inhibition was found to be 32.8 and 42.18%, respectively (Fig. 5).
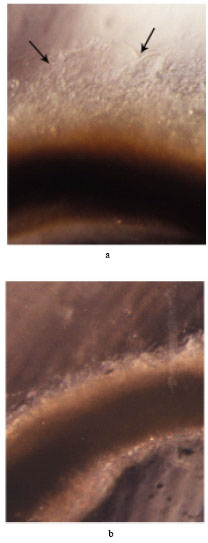 | |
| Fig. 3: | Inhibition of microvessel sprouting from rat aortic ring by rutin; Microvessel sprouting after 6 days in collagen matrices incubated in (a) conditioned medium from B16-F10 cells and (b) conditioned medium and rutin at 25 μg mL-1 |
Effect of rutin on endothelial cell tube formation: Rutin treatment could significantly inhibit the endothelial cell tube formation in a concentration dependent manner and maximum inhibition was observed at 25 μg mL-1 (Fig. 6).
Effect of Rutin on VEGF, IL-1β and TNF-α production and gene expression: Rutin treatment could significantly inhibit the production of VEGF and IL-1β from TAMs. Rutin at a concentration of 25 μg mL-1, showed 26.81 and 34.21% inhibition in the production of VEGF and IL-1β, respectively. Gene expression studies also showed remarkable inhibition of VEGF and IL-1β mRNA (Fig. 7a, b). Rutin treatment could significantly stimulate the TNF-α production and mRNA expression in TAMs (Fig. 7c).
Effect of rutin on TNF-α bioassay: The culture supernatants from rutin treated TAMs showed maximum cytotoxic effect to TNF-α sensitive L929 cells at a concentration of 25 μg mL-1 compared to those treated with 5 and 10 μg mL-1 (Fig. 8).
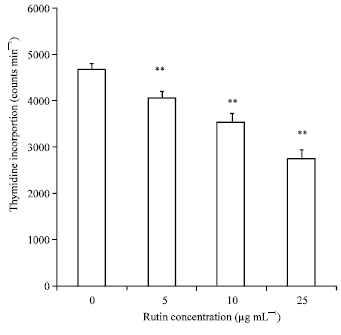 | |
| Fig. 4: | Inhibition of Human Umbilical Vein Endothelial Cell (HUVEC) proliferation by rutin. HUVEC (5x104 cells) were cultured for 24 h in the presence of increasing concentrations of rutin (5-25 μg mL-1) followed by 18 h incubation after adding [3H] thymidine (1 μCi). Measured radioactivity were expressed as mean counts per minute (cpm)±SD. **p<0.001 rutin treated versus untreated groups; Representative of three separate experiments |
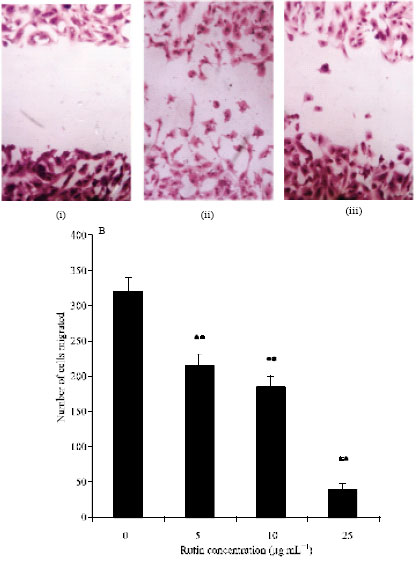 | |
| Fig. 5: | Crystal violet staining (a) and migration rates (b) of endothelial cells; (a) Wound migration assay was performed on the confluent monolayer of endothelial cells. Increasing concentrations of rutin (5-25 µg mLG1) were added to HUVEC monolayer after creating the wound and incubated for 48 h followed by crystal violet (0.1%) staining. (i) 0 h control (ii) 48 h control (iii) 48 h rutin at 25 µg mLG1 (B) Migration rates of endothelial cells. Values are expressed as mean ± SD (n = 3). The asterisks indicate statistic significance (p<0.001) as a comparison between the treated and respective control groups. Note that rutin produced dose-dependent inhibition of endothelial migration |
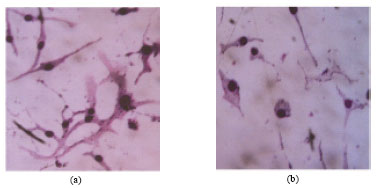 | |
| Fig. 6: | Diff-Quick staining of endothelial cell tube formation. Endothelial cells (5H104) were cultured with increasing concentrations of rutin (5-25 µg mLG1) and for 48 h followed by Diff-Quick staining (a) 48 h Control and (b) 48 h rutin at 25 µg mLG1 |
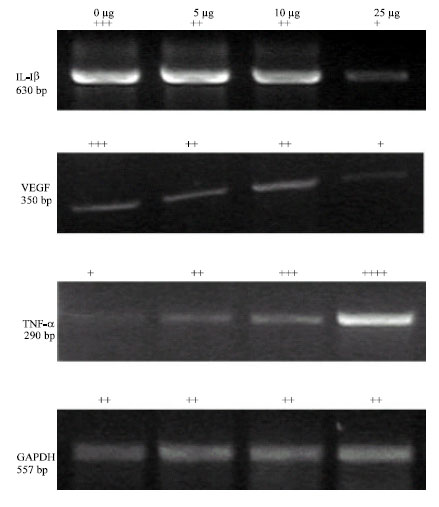 | |
| Fig. 7: | Effect of Rutin on the expression of IL-1β, VEGF and TNF-α in TAMs. TAMs (5x104 cells) were cultured with increasing concentrations of rutin (5-25 μg mL-1) for 24 h; cDNA was synthesized followed by amplification using specific primers for IL-1β, VEGF and TNF-α |
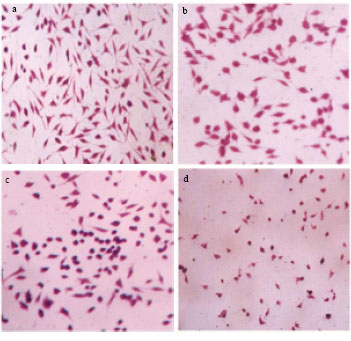 | |
| Fig. 8: | Effect of Rutin on TNF-α bioassay (a) 48 h control, (b) rutin at 5 μg mL-1 (c) rutin at 10 μg mL-1 and (d) rutin at 25 μg mL-1 |
DISCUSSION
In the present study we evaluated the inhibitory effect of rutin on the angiogenic process both in vitro and in vivo. We for the first time showed that rutin could alter the VEGF, IL-1β and TNF-α production in tumor associated macrophages. It has been proved conclusively that angiogenesis is essential for a tumor to become large and malignant; hence inhibition of the process of angiogenesis either by inhibiting endothelial cells or by suppression of angiogenic growth factors should provide a novel therapeutic window for cancer therapy. Here in, we focused on studying the potential effect of rutin on the process of angiogenesis itself using vascular endothelial cells as a model. We demonstrated that rutin could inhibit tube formation of HUVECs using in vitro matrigel assay. Matrigel is a matrix of mouse basement membrane neoplasm, which represents a complex mixture of basement membrane proteins including type IV collagen, entactin/nitrogen, proteoheparan sulfate and other growth factors. Matrigel can induce endothelial cells to differentiate as shown by morphological changes and is widely accepted model to study biochemical changes associated with angiogenesis (Ponce et al., 2001). The inhibition of tube formation, as observed in our study after rutin treatment might be due to the reduced proliferation of endothelial cells. The concentrations of rutin used in the present studies did not interfere with the cell viability of endothelial cells. The decreased number of tumor directed capillaries in rutin treated animals also indicates its potential to selectively inhibit the tumor specific capillary sprouting. This inhibition of capillary formation might be one of the reasons for the antimetastatic effect of rutin (Lata et al., 1995).
A prime regulator for angiogenesis is believed to be Vascular Endothelial Growth Factor (VEGF) and low oxygen tension dramatically induces the expression of this major angiogenic factor in tumor associated macrophages (Schoppmann et al., 2002; Forsythe et al., 1996; Schioppa et al., 2003). VEGF expression is closely associated with increased intratumoral microvessel density in breast cancer patients (Toi et al., 1996). Rutin treatment showed significant inhibition of microvessel sprouting from aortic ring induced by VEGF rich conditioned medium. Quantification of growth factor VEGF by ELISA in our studies indicated a decreased amount of VEGF in rutin treated TAM culture supernatant. This may be due to the down regulatory action of rutin on the VEGF mRNA expression as there was only low level expression of VEGF mRNA when TAMs were treated with rutin.
During tumor development, the cross-talk between tumor associated macrophages and surrounding cells is crutial. With regards to the cross-talk between inflammatory cells and endothelial cells present in the tumor microenvironment, IL-1β stimulates the proliferation of endothelial cells, adhesion molecule expression and production of cytokines and inflammatory molecules in vitro; suggesting that IL-1 related molecules may be deeply involved in the control of angiogenesis process (Dianarello, 1996, 1998). IL-1β is a pleiotropic cytokine that induces immunosuppression under different experimental conditions (Dianarello, 1996). The relevance of microenvironmental IL-1 in angiogenesis and in the invasiveness of different tumor cells has been recently demonstrated using IL-1β knockout (KO) mice (Voronov et al., 2003). IL-1β upregulates HIF-1α in a human lung cancer cell line (A549) activating NF-êB/Cox-2 pathway (Jung et al., 2003). Present studies revealed that rutin could inhibit the expression of IL-1β in TAMs. The inhibition of IL-1β expression by rutin as observed in our results also might have occurred through similar mechanisms. Further studies in this area are under progress to fully elucidate the mechanism of action of rutin on the regulation of IL-1β in TAMs.
In inflammation and angiogenesis, TNF-α plays a dual role: it controls both tissue destruction and recovery and these roles are determined by the context in which this cytokine acts (Fajardo et al., 1992). With regards to cancer development and progression, high doses of TNF-α induce death of tumor cells (Lejeune et al., 1998). However, low concentrations of TNF-α promote tumor growth and spread (Foa et al., 1990). Thus TNF-α might stimulate tumor development by promoting vessel growth and participate in tumor and endothelium destruction by direct cytotoxicity. Present studies reveal that rutin treatment could stimulate the production of TNF-α in TAMs and these levels could induce cytotoxicity to tumor cells as evidenced by TNF-α sensitive L929 bioassay.
In conclusion, the present study demonstrates the antiangiogenic potential of rutin and its regulatory effect on the production of VEGF, IL-1β and TNF-α in TAMs. Since macrophages present within the solid tumor contribute to the initiation of angiogenesis, the present study is of significant relevance of developing rutin as potential therapeutic target against TAMs in controlling angiogenesis.
ACKNOWLEDGMENT
The authors would like to thank Dr. Ramadasan Kuttan, Research Director for his active support during the period of study.
REFERENCES
- Balkwill, F. and A. Mantovani, 2001. Inflammation and cancer: Back to virchow. Lancet, 357: 539-545.
Direct Link - Barbera-Guillem, E., J.K. Nyhus, C.C. Wolford, C.R. Friece and J.W. Sampsel, 2002. Vascular endothelial growth factor secretion by tumor-infiltrating macrophages essentially supports tumor angiogenesis and IgG immune complexes potentiate the process. Cancer Res., 62: 7042-7049.
Direct Link - Bergers, G. and L.E. Benjamin, 2003. Tumorigenesis and the angiogenic switch. Nat. Rev. Cancer, 3: 401-410.
Direct Link - Bombardelli, E. and P. Morazzoni, 1993. The flavonoids: New perspectives in biological activities and therapeutics. Chim. Oggi., 11: 25-28.
Direct Link - Cheng, N. and J. Chen, 2001. Tumor necrosis factor-alpha induction of endothelial ephrin A1 expression is mediated by a p38 MAPK-and SAPK/JNK-dependent but nuclear factor-kappa B-independent mechanism. J. Biol. Chem., 276: 13771-13777.
Direct Link - Choi, Y.H., L. Zhang, W.H. Lee and K.Y. Park, 1998. Genistein-induced G2/M arrest is associated with the inhibition of cyclin B1 and the induction of p21 in human breast carcinoma cells. Int. J. Oncol., 13: 391-396.
Direct Link - Davis, J.N., B. Singh, M. Bhuiyan and F.H. Sarkar, 1998. Genistein-induced upregulation of p21WAF1, downregulation of cyclin B and induction of apoptosis in prostate cancer cells. Nutr. Cancer, 31: 123-131.
Direct Link - Deschner, E.E., J. Ruperto, G. Wong and H.L. Newmark, 1991. Quercetin and rutin as inhibitors of azoxymethanol-induced colonic neoplasia. Carcinogenesis, 12: 1193-1196.
PubMed - Dianarello, C.A., 1996. Biologic basis for interleukin-1 in disease. Blood, 87: 2095-2147.
Direct Link - Dinarello, C.A., 1998. Interleukin-1, interleukin-1 receptors and interleukin-1 receptor antagonist. Int. Rev. Immunol., 16: 457-499.
PubMed - Fajardo, I.F., H.H. Kwan, J. Kowalski, S.D. Prionas and A.C. Allison, 1992. Dual role of tumor necrosis factor-alpha in angiogenesis. Am. J. Pathol., 140: 539-544.
Direct Link - Foa, R., M. Massaia, S. Cardona, A.G. Tos and A. Bianchi et al., 1990. Production of tumor necrosis factor-alpha by β-cell chronic lymphocytic leukemia cells: A possible regulatory role of TNF in the progression of the disease. Blood, 76: 393-400.
Direct Link - Forsythe, J.A., B.H. Jiang, N.V. Iyer, F. Agani, S.W. Leung, R.D. Koos and G.L. Semenza, 1996. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol. Cell. Biol., 16: 4604-4613.
PubMedDirect Link - Fotsis, T., M. Pepper, H. Adlercreutz, G. Fleischmann, T. Hase, R. Montesano and L. Schweigerer, 1993. Genistein, a dietary-derived inhibitor of in vitro angiogenesis. Proc. Natl. Acad. Sci. USA., 90: 2690-2694.
Direct Link - Ghezzi, P., C.A. Dinarello, M. Bianchi, M.E. Rosandich, J.E. Repine and C.W. White, 1991. Hypoxia increases production of interleukin-1 and tumor necrosis factor by human mononuclear cells. Cytokine, 3: 189-194.
Direct Link - Gupta, K., S. Kshirasagar, L. Chang, R. Schwartz and P.Y. Law, 2002. Morphine stimulates angiogenesis by activating proangiogenic and survival promoting signaling and promotes breast tumor growth. Cancer Res., 62: 4491-4498.
Direct Link - Ihme, N., H. Kiesewetter, F. Jung, K.H. Hoffmann, A. Birk, A. Muller and K.I. Grutzner, 1996. Leg edema protection from a buckwheat herb tea in patients with chronic venous insufficiency. A single-center, randomized, double-blind, placebo controlled clinical trial. Eur. J. Clin. Pharmacol., 50: 443-447.
PubMed - Jung, Y.J., J.S. Issacs, S. Lee, J. Trepel and L. Neckers, 2003. IL-1β-mediated up-regulation of HIF-1α via and NF-κB/Cox-2 pathway identifies HIF-1 as a critical link between inflammation and oncogenesis. FASEB J., 17: 2115-2117.
Direct Link - Kataki, A., P. Scheid, M. Piet, B. Marie, N. Martinet, Y. Martinet and J.M. Vignaud, 2002. Tumor infiltrating lymphocytes and macrophages have a potential dual role in lung cancer by supporting both host-defense and tumor progression. J. Lab. Clin. Med., 140: 320-328.
CrossRefDirect Link - Kerbel, R. and J. Folkman, 2002. Clinical translation of angiogenesis inhibitors. Nat. Rev. Cancer, 2: 727-739.
CrossRefDirect Link - Menon, L.G., R. Kuttan and G. Kuttan, 1995. Inhibition of lung metastasis in mice induced B16F-10 melanoma cells by polyphenolic compounds. Cancer Lett., 95: 221-225.
Direct Link - Lejeune, F.J., C. Ruegg and D. Lienard, 1998. Clinical applications of TNF-α in cancer. Curr. Opin. Immunol., 10: 573-580.
Direct Link - Lian, F., M. Bhuiyan, Y.M. Li, N. Wall, M. Kraut and F.H. Sarkar, 1998. Genistein-induced G2-M arrest, p21WAF1 upregulation and apoptosis in a non-small-cell lung cancer cell line. Nutr. Cancer, 31: 184-191.
Direct Link - Lingen, M.W., 2001. Role of leukocytes and endothelial cells in the development of angiogenesis in inflammation and wound healing. Arch. Pathol. Lab. Med., 125: 67-71.
Direct Link - Mantovani, A., B. Bottazzi, F. Colatta, S. Sozani and L. Ruco, 1992. The origin and function of tumor associated macrophages. Immunol. Today, 13: 265-270.
Direct Link - Matsukawa, Y., N. Marui, T. Sakai, Y. Satomi, M. Yoshida, K. Matsumoto, H. Nishino and A. Aoike, 1993. Genistein arrests cell cycle progression at G2-M. Cancer Res., 53: 1328-1331.
Direct Link - Metodiewa, D., A. Kochman and S. Karolczak, 1997. Evidence for antiradical and antioxidant properties of four biologically active N, N-diethylaminoethyl ethers of flavanone oximes: A comparison with natural polyphenolic flavonoid (rutin) action. Biochem. Mol. Biol. Int., 41: 1067-1075.
Direct Link - Naldini, A., A. Pucci and F. Carraro, 2001. Hypoxia induces the expression and release of interleukin 1 receptor antagonist in mitogen activated mononuclear cells. Cytokine, 13: 334-341.
Direct Link - Oomah, B.D. and G. Mazza, 1996. Flavonoids and antioxidative activities in buckwheat. J. Agric. Food Chem., 44: 1746-1750.
CrossRefDirect Link - Ponce, M.L., M. Nomizu and H.K. Kleinman, 2001. An angiogenic laminin site and its antagonist bind through the alpha (v) beta 3 and alpha 5 beta 1 integrins. FASEB J., 15: 1389-1397.
Direct Link - Romero, I., A. Paez, A. Ferruelo, M. Lujan and A. Berenguer, 2002. Polyphenols in red wine inhibit the proliferation and induce apoptosis of LNCaP cells. BJU Int., 89: 950-954.
Direct Link - Roy, S., S. Khanna, H.M. Alessio, J. Vider, D. Bagchi, M. Bagchi and C.K. Sen, 2002. Anti-angiogenic property of edible berries. Free Radic. Res., 36: 1023-1031.
CrossRefDirect Link - Schioppa, T., B. Uranchimeg, A. Saccani, S.K. Biswas and A. Doni et al., 2003. Regulation of the chemokine receptor CXCR4 by hypoxia. J. Exp. Med., 198: 1391-1402.
Direct Link - Schoppmann, S.F., P. Birner, J. Stockl, R. Ullrich and C. Caucig et al., 2002. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Int. J. Pathol., 161: 947-956.
Direct Link - Toi, M., S. Kondo, H. Suzuki, Y. Yamamoto, K. Inada, T. Imazawa, T. Taniguchi and T. Tominaga, 1996. Quantitative analysis of vascular endothelial growth factor in primary breast cancer. Cancer, 77: 1101-1106.
Direct Link - Voronov, E., D.S. Shouval, Y. Krelin, E. Carnano and D. Benharroch et al., 2003. IL-1 is required for tumor invasiveness and angiogenesis. Proc. Natl. Acad. Sci. USA., 100: 2645-2650.
Direct Link