
Noha A. Khalil
Laboratory of Fish Reproduction, National Institute of Oceanography and Fisheries, Alexandria, Egypt
Abd El-Hakim E. El-Gamal
Laboratory of Fish Reproduction, National Institute of Oceanography and Fisheries, Alexandria, Egypt
Sohair A. Gaber
Laboratory of Fish Reproduction, National Institute of Oceanography and Fisheries, Alexandria, Egypt
Mostafa A. Mousa
Laboratory of Fish Reproduction, National Institute of Oceanography and Fisheries, Alexandria, Egypt
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 7 | Page No.: 1102-1111







ABSTRACT
The present study investigated the concomitant changes of mGnRH and SL immunoreactivities within the brain and the pituitary gland at different stages of the gonadal cycle of Lates niloticus. mGnRH cell bodies were observed in the mediobasal hypothalamus within the Nucleus Lateralis Tuberis Pars posterior (NLTP). Cells with immunoreactivity for SL were detected in the Pars Intermedia (PI) of the pituitary gland. The synthetic and secretory activity of both mGnRH-ir neurons and SL-IR cells were increased during sexual maturation and spawning. Both mGnRH-ir neurons and SL-ir cells were small with moderate immunoreactivity in immature fish. However, the synthetic activity of these cells was increased as indicated by the increased numbers and sizes of cells with more immunoreactivity during gonadal development. In late stages of maturation (prespawning), the synthetic activity of mGnRH-ir neurons and SL-ir cells reached their maximum in ripe fish. Finally, during spawning, mGnRH-ir neurons and SL cells showed an increase in the secretory activity in spent fish as reflected by their small sizes, vacuolated appearance and weak immunoreactivity. Furthermore, mGnRH-ir fibers, which are in close association with SL-ir cells, showed similar seasonal changes in the density and immunoreactivity as for mGnRH-ir neurons and SL cells. The dominance of mGnRH in the hypothalamus and its close contact with SL-ir cells in the pituitary gland, together with the concurrent changes of GnRH and SL immunoreactivity in relation to seasonal changes in the reproductive conditions suggest that mGnRH plays an important role for SL release to regulate some biological events related to the reproduction in L. niloticus.
PDF Abstract XML References Citation
How to cite this article
Noha A. Khalil, Abd El-Hakim E. El-Gamal, Sohair A. Gaber and Mostafa A. Mousa, 2007. Immunohistochemical Localization of Gonadotropin-Releasing Hormone and Somatolactin During Sexual Maturation and Spawning of Lates niloticus. Journal of Biological Sciences, 7: 1102-1111.
DOI: 10.3923/jbs.2007.1102.1111
URL: https://scialert.net/abstract/?doi=jbs.2007.1102.1111
DOI: 10.3923/jbs.2007.1102.1111
URL: https://scialert.net/abstract/?doi=jbs.2007.1102.1111
INTRODUCTION
The Nile perch, Lates niloticus, is the most economically important fish species in Egypt living in tropical and semitropical waters. It grows fast but is less salt-tolerant than Lates calcarifer and attains 190 cm in length with a maximum weight of 200 kg. The characterization and the anatomical distribution of adenohypophyseal hormones (Mousa, 2001) and GnRH forms (Mousa and Mousa, 2003) was demonstrated in the Nile perch, L. niloticus. Gonadotropin-releasing hormone (GnRH) is a decapeptide neuroendocrine hormone that is considered to play important roles in the regulation of teleosts reproduction, mainly by stimulation of gonadotropin release from the pituitary gland (Peter et al., 1991).
Teleost fishes lack a functional-hypophyseal portal system, but GnRH nerve fibers terminate in the vicinity of the pituitary gonadotrophs and SL producing cells (Muske, 1993; Mousa and Mousa, 2003). SL is involved in different physiological processes. Several studies suggested the involvement of SL in the acid-base and calcium regulation (Kaneko and Hirano, 1993; Kakizawa et al., 1996; 1997), phosphate and fat metabolism (Lu et al., 1995; Company et al., 2000) and background adaptation (Zhu and Thomas, 1996, 1998; Nguyen et al., 2006) in fish. SL has also been linked to energy mobilization in response to stressors (Rand-Weaver et al., 1993; Johnson et al., 1997), exhaustive exercise (Kakizawa et al., 1995a) and fasting (Company et al., 2001; Mingarro et al., 2002). Interestingly, other observations suggested that SL may act as a reproductive hormone. For instance, SL stimulated a weak production of ovarian and testicular steroids in a dose-dependent manner in coho salmon in vitro (Planas et al., 1992) and gonadectomy significantly reduced plasma SL levels and pituitary content in Atlantic salmon, Salmo salar (Mayer et al., 1998). This concurs with the finding that the SL-ir cells in the PI are activated during the reproductive phase in the genus Oncorhynchus (Olivereau and Rand-Weaver, 1994a and b) and Odontesthes bionariensis (Vissio et al., 2002). In addition, Mousa and Mousa (1999 and 2000) observed seasonal variations in the synthetic and secretory activity of SL-ir cells concomitant with the development of the gonads and spawning in O. niloticus and Mugil cephalus. Recently, a growing body of data suggests the involvement of GnRH in regulating the secretion of various pituitary hormones including gonadotropin hormones (GTHs). Direct morphological evidences of a close association of mGnRH fibers with various pituitary hormone secreting cells including SL-ir cells were observed in L. niloticus (Mousa and Mousa, 2003). Also, evidence suggests that SL-producing cells are also regulated by GnRH in Oncorhynchus (Kakizawa et al., 1997; Taniyama et al., 2000; Kumar Bhandari et al., 2003). Furthermore, Parhar and Iwata (1994) have observed GnRH-ir fibers projecting to SL-ir cells in the steelhead trout, Oncorhynchus. mykiss. In addition, direct morphological evidence of a close association of GnRH fibers with Growth Hormone (GH), prolactin (PRL), GTH and SL-ir cells was observed in the pejerry, Odontesthes bonariensis (Vissio et al., 1999). Therefore, the aim of the present study was to investigate mGnRH and SL immunoreactivities at different stages of gonadal cycle to assess the possible actions of these hormones in sexual maturation and spawning of L. niloticus.
MATERIAL AND METHODS
Fish collection: Immature and mature Lates niloticus of both sexes with standard length (larger than 20 cm) were collected alive at intervals of about one month throughout the year from the Nile River (Delta region). However, during the prespawning and spawning seasons (March to July), fish were collected at intervals of about 15 days to ensure that all stages of gonad maturation were included.
Measurements and classification of maturity stages: After the collection of fishes, their total and standard lengths were measured to the nearest 0.1 cm. Before dissection, the fishes were anesthetized in a solution (40 mg L-1) of clove oil (Sigma) before handling (Mousa, 2004b). The gonads were extirpated from the body cavity, weighed to the nearest 0.01 g. Gonadosomatic index GSI = (Gonad weight/Gutted fish weight) X100, oocyte diameter and histological appearance of the gonads were used as indices of different maturity stages (Mousa, 2004a). Five sexual maturity stages were signified for males: stage I (immature), stage II, stage III, stage IV (ripe) and stage V (spent or postspawning). For females, six ovarian stages were distinguished according to morphological, microscopical appearance, egg diameter and GSI data: stage I, stage II, stage III, stage IV, stage V and stage VI (spent).
Tissue processing: Anesthetized fishes were perfused via the ascending aorta with 20 mL of 0.9% saline, followed by 50 mL of Bouin`s fluid at 4°C. Immediately after the dissection, the pituitary gland, attached to the brain, was postfixed in Bouin's fluid for 24 h at 4°C. Thereafter, the brain and pituitaries were dehydrated through up graded ethanol solution, cleared and then embedded in paraplast (M.P. 56-58°C). Consecutive median sagittal sections of the brain and the pituitary gland were made at 4 μm thickness.
Immunocytochemical procedures: Antibodies: Rabbit antiserum directed against mammalian GnRH (Lot No. 83LRF) was kindly provided by Dr. G. Tramu (Talence, France). In addition, antiserum directed against chum salmon somatolactin (Lot No. 8906) was obtained from Dr. H. Kawauchi (School of Fisheries Science, Kitasato University, Iwate, Japan).
Immunocytochemical reactions: To study co-localization of mGnRH and SL using double peroxidase immunohistochemistry as described previously (Mousa and Mousa, 2003), generally performed with a vectastain ABC (Avidin-biotin peroxidase complex) Kit (Vector Laboratories). The sections of the pituitary gland and brain were deparaffinized in xylene, rehydrated through graded ethanol and washed in phosphate-buffered saline (PBS). Then, the sections were treated with 0.6% H2O2 for 30 min to inactivate peroxidase in ABC and washed in several changes of PBS. Sections stained for mGnRH were then incubated with a second primary antibody against SL (Lot No. 8906, 1:1000 dilution) (H. Kawauchi, School of Fisheries Science, Kitasato University, Iwate, Japan). All incubations were done overnight at 4°C, slides were then washed in PBS, exposed to the biotinylated secondary antibody (Vector Laboratories) for 1 h. and with avidin- biotin- conjugated peroxidase for 45 min. Finally, the sections were washed and stained with 3`, 3`- diaminobenzidine tetrahydrochloride (DAB) (Sigma) including 0.01% H2O2 in 0.05 M Tris-buffered saline (pH 7.6) for 3-5 min. Sections stained with anti-GnRH antibody were washed in several changes of PBS. Sections were then incubated, overnight at 4°C, with a second primary antibody against chum salmon somatolactin, washed in PBS, exposed to the biotinylated secondary antibody (Vector Laboratories) for 1 h. and with avidin-biotin-conjugated peroxidase for 45 min. Finally, the sections were washed and stained with 3`, 3`- diaminobenzidine tetrahydrochloride (DAB) (Sigma) including 0.01% H2O2 in 0.05 M Tris-buffered saline (pH 7.6) for 3-5 min. During staining with DAB, the nickel solution was used to differentiate between the double immunostaining. The chromogen DAB used for the first primary antiserum appeared black, whereas the one used for the second primary antiserum appeared brown. After the enzyme reaction, the sections were washed in tap water, dehydrated in alcohol, cleared in xylene and mounted in DPX.
Specificity controls: To demonstrate specificity of staining, the following controls were included as mentioned in details elsewhere (Mousa and Mousa, 2003): (1) Preabsorption of antibody against mGnRH and SL with a synthetic peptide for mGnRH (Sigma) and SL (H. Kawauchi) for 24 h at 4°C (2) omission of either the primary antisera, the secondary antibodies or avidin-biotin complex (3) omission of either the first or second primary antibody and either the first or second and secondary antibody.
Cell size measurements: Quantification of SL-immunoreactive cells in the PI was calculated from four sections of each individual animal (n = 8 in each maturity stage). The soma sizes of neurons immunoreactive to mGnRH in the NLTP as well as SL-ir pituitary cells were measured using computer-aided analysis (the Image-Pro Analysis package, Media Cybernetics) of digital images viewed via microscope (Axioskope, Zeiss, Oberkochen, Germany). A3CCD color video camera, (Sony) was used for a minimum of 50 mGnRH-ir neurons from each brain area under investigation (NLTP) per animal as well as 50 SL-ir cells from PI, the cross-sectional area was measured for neurons with the nucleus in the plane of section.
Statistical analysis: All values are expressed as the mean the standard deviation of the mean. Results were analyzed with the SPSS (Statistical Package for Social Sciences) statistical package. Paired-samples t test was applied to compare means. The level for accepted statistical significance was p≤0.05.
RESULTS
Immunolocalization of mGnRH and SL in the brain and pituitary of L. niloticus: The present immunocytochemical study, using specific antibodies for mGnRH and chum salmon SL, demonstrated that mGnRH-ir cell bodies exist in the mediobasal hypothalamus in the NLTP in the brain of female (Fig. 1A, C, E and G) and male (Fig. 2A, C, E and G) L. niloticus during sexual maturation and spawning. SL-ir cells and were only detected in the PI of female (Fig. 1B, D, F and H) and male (Fig. 2B, D, F and H). Immunoreactivities were observed neither in the Rostral Pars Distalis (RPD) nor in the Proximal Pars Distalis (PPD), indicating no cross-reactions with PRL- or GH-expressing cells.
Immunostaining of mGnRH and SL during ovarian maturation and spawning in female L. niloticus: The changes in number, size and intensity of the immunoreactivities of mGnRH-ir neurons and SL-ir cells in the brain and pituitary gland, respectively, represent seasonal variations, concomitant with the development of the ovaries and spawning as illustrated in Fig. 1 and Table 1.
In the previtellogenic female shown (stage I) (Fig. 1A, B and Table 1), mGnRH- and SL-ir cells were small and few in number when compared to that of vitellogenic and ripe females. With the onset of vitellogenesis (yolk deposition; early-, mid- and late-vitellogenesis), mGnRH- and SL-ir cells exhibit strong immunoreactivity and increased in both number and size (Fig. 1C, D and Table 1). The majority of mGnRH- and SL-ir cells exhibited an increase in immunoreactivity and only few of them appeared to empty their secretory contents as indicated by small cell size and weak immunoreactivity. During the ripening stage of the ovaries, the synthetic activity of mGnRH- and SL-ir cells reaches their maximum as reflected with strong immunoreactivity and increased sizes and numbers (Fig. 1E, F and Table 1). Also, mGnRH-ir neurons equipped with long and thick axons (Fig. 1E). By approach of the postspawning phase of the ovary, the immunostaining intensities and size of mGnRH- and SL-ir cells were markedly reduced compared to that of ripe female (Fig. 1G, H and Table 1). In the postspawned fish with spent ovaries, most of mGnRH- and SL-ir cells appeared to empty their secretory contents as indicated by their small size and vacuolated cytoplasm (Fig. 1G, H and Table 1).
mGnRH-ir fibers showed similar pattern of activity. They exhibited high intensities and gradual increase of mGnRH immunoreactivity during ovarian maturation (Fig. 1D). At ripening of the ovary, mGnRH-ir fibers exhibited high density and increased immunoreactivity compared to that of late-vitellogenic females (Fig. 1F). After spawning, the density and immunoreactivity of mGnRH-ir fibers were decreased (Fig. 1H).
Immunostaining of mGnRH and SL during testicular maturation and spawning in male L. niloticus: Similar to female L. niloticus, mGnRH- and SL-ir cells in the brain and pituitary of male showed seasonal changes in their synthetic and secretory activity during testicular maturation and spawning.
| Table 1: | Maturational changes in gonadosomatic index (GSI %), GnRH-ir soma number and sizes (μm2) in the nucleus preopticus (NLTP) and SL-ir cell number and sizes (μm2) in the pituitary during ovarian maturation in females of L. niloticus |
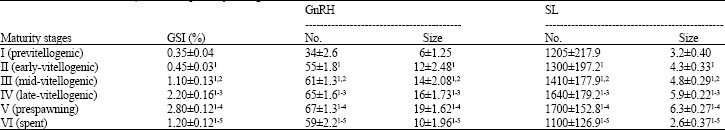 | |
| Each value represents the mean±SD of 8 fishes, (1): Mean was significantly higher than that of Previtellogenesis (p<0.05), (2): Mean was significantly higher than that of Early Vitellogenesis (p<0.05), (3): Mean was significantly higher than that of Mid-vitellogenesis (p<0.05), (4): Mean was significantly higher than that of Late-vitellogenesis (p<0.05), (5): Mean was significantly higher than that of prespawning (p<0.05) | |
| Table 2: | Maturational changes in gonadosomatic index (GSI %), GnRH-ir soma number and sizes (μm2) in the nucleus preopticus (NLTP) and SL-ir cell number and sizes (μm2) in the pituitary during testicular maturation in males of L. niloticus |
 | |
| Each value represents the mean±SD of 8 fishes, (1): Mean was significantly higher than that of immature stage (p<0.05), (2): Mean was significantly higher than that of stimulating spermatogenesis (p<0.05), (3): Mean was significantly higher than that of rapid spermatogenesis (p<0.05), (4): Mean was significantly higher than that of ripe (p<0.05) | |
Both the synthetic and secretory activities of mGnRH- and SL-ir cells were gradually increased during spermatogenesis as reflected by increase in number, size and immunoreactivity (Fig. 2 and Table 2). In the immature males (prespermatogenesis; stage I), mGnRH- and SL-ir cells were small in sizes with moderate immunoreactivity (Fig. 2A, B and Table 2). With the onset of spermatogenesis (stimulating spermatogenesis; stage II) and as spermatogenesis progressed (rapid spermatogenesis and ripening), mGnRH- and SL-ir cells showed an increase in number and in the synthetic activity as reflected by large sizes and strong immunoreactivity (Fig. 2C and D).
During the ripening stage of the testis, the synthetic activity of mGnRH- and SL-ir cells reaches their maximum as reflected with strong immunoreactivity and increased sizes and numbers (Fig. 2E, F and Table 1). Also, mGnRH-ir neurons equipped with long and thick axons (Fig. 1E).
After spawning, mGnRH- and SL-ir cells showed a marked decrease in the synthetic activity as reflected by decreased cell numbers, sizes and weak immunoreactivity in the postspawned fish with spent testes, compared with those of spermatogenic and ripe males (Fig. 2G, H and Table 2). In spent males, most of mGnRH- and SL-ir cells appeared to empty their secretory contents as indicated by their small size and vacuolated cytoplasm (Fig. 2G, H and Table 2). Also, mGnRH-ir fibers exhibited high intensities and gradual increase of immunoreactivity during testis maturation (Fig. 2B, D and F). After spawning, in spent males, the density and staining intensities of mGnRH-ir fibers were greatly reduced compared with those of ripe males (Fig. 2H).
DISCUSSION
In the present immunohistochemical study, using specific antibodies for mGnRH and chum salmon SL, we investigated the expression of these hormones in the brain and pituitary gland of L. niloticus during sexual maturation and spawning. We showed that mGnRH cell bodies were located in the mediobasal hypothalamus in the NLTP. However, SL-ir cells were only detected in the PI of the examined fishes. Our double immunostaining revealed a close association between mGnRH-ir fibers and SL-ir cells within the pituitary gland. We demonstrated also that the gradual increase of the synthetic activity of mGnRH-ir neurons as well as SL-ir cells was concomitant with the gonads maturation and this increase reached their maximum during ripening stages in both sexes. However, there is an apparent decrease in the synthetic activity of these cells after spawning. Taken together, the present findings suggest that mGnRH plays an important role in the neurohormonal regulation of SL pituitary hormone in the Nile perch.
The present study demonstrated GnRH-ir nerve fibers in close contact with SL-ir cells within the pituitary gland. This finding confirms our previous study (Mousa and Mousa, 2003) and is in agreement with the previous studies in O. mykiss (Parhar and Iwata, 1994) and O. bonariensis (Vissio et al., 1999), showing GnRH-ir fibers ending in contact with SL-ir cells within the pituitary gland. SL is a novel pituitary hormone which is considered to be involved in various physiological functions related to environmental adaptation and sexual maturation (Kaneko, 1996) in teleost fishes. However, GnRH is a principal mediator for secretion of pituitary hormones with specific regards to GTH. GnRH agonist binding sites were observed in GH-, prolactin (PRL)- and SL-immunoreactive cells of other teleost species (Stefano et al., 1999). Consistently, sGnRH stimulated release of GH from steroid-primed pituitaries of rainbow trout (Holloway and Leatherland, 1997), PRL from tilapia pituitaries (Weber et al., 1997) and SL from cultured pituitaries of rainbow trout (Kakizawa et al., 1997).
Extending our previous study (Mousa and Mousa, 2003) we showed that, with the onset of vitellogenesis (yolk deposition; early- mid- and late-vitellogenesis), mGnRH-ir neurons in the hypothalamus as well as SL-ir cells within the pituitary exhibit a gradual increase in the synthetic activity as reflected by increased cell sizes and immunoreactivity with few cells appeared to empty their secretory contents as indicated with small sizes and weak immunoreactivity in L. niloticus. Similarly, as spermatogenesis progressed (rapid spermatogenesis), the increase in the synthetic activity of mGnRH- and SL-ir cells becomes prominent as reflected by enlarged cell sizes and stronger immunoreactivity. Also, mGnRH-ir fibers density was gradually increased during gonadal maturation. These observations are consistent with the previous immunocytochemical studies obtained in Oncorhynchus nerka and O. keta (Olivereau and Rand-Weaver, 1994a, b), O. niloticus (Mousa and Mousa, 1999), M. cephalus (Mousa and Mousa, 2000) and in O. bonariensis (Vissio et al., 2002) showing an increase in the synthetic activity of SL-ir cells as reflected with the increase in cell number and sizes during sexual maturation of these teleost species. The present immunocytochemical results are in accordance with the biochemical studies on Sebastes rastrelliger (Collins et al., 2001) and Myxine glutinosa (Kavanaugh et al., 2005), which showed seasonal changes of GnRH levels during the annual reproductive cycle of these fishes. The present results are also in agreement with biochemical studies on Oncorhynchus kisutch which showed that SL stimulates gonadal steroidogenesis in vitro (Planas et al., 1992). Rand-Weaver et al. (1992 and 1995) found that in O. kisutch during the period of gonadal growth, plasma SL levels increased and were highly correlated to estradiol levels in females and 11-ketotestosterone levels in males. The previous studies have demonstrated that GnRH analogs stimulate the expression of genes encoding GH, PRL and SL during particular periods of the life cycle using primary pituitary cell cultures of masu salmon (Taniyama et al., 2000; Onuma et al., 2005). Therefore, the authors suggested that GnRH appears to directly and/or indirectly regulate gene expression for GH, PRL and SL in combination with the pituitary-gonadal axis, such as sex steroid hormones. Taken together, the increase of synthetic activity of mGnRH-ir neurons in the brain together with SL-ir cells in the pituitary during sexual maturation support the notion that mGnRH has stimulatory effects on SL expression and/or release during sexual maturation of teleosts. This notion concurs with the finding that the morphology of GnRH neurons is sensitive to the influence of gonadal steroids in Rana pipiens (Tsai and Jones, 2005). In addition, in Salmo salar, the mature testes have a stimulatory effect on the synthesis and release of SL (Mayer et al., 1998).
During the ripening stage of the ovaries in L. niloticus, the synthetic activity of mGnRH- and SL-ir cells reached their maximum as reflected with a marked hypertrophy and strong immunoreactivity. During this stage, mGnRH-ir fibers exhibited high density and increased immunoreactivity. These observations are in agreement with previous studies showing that the pituitary amounts of sGnRH in the pre-spawning period were higher than those in early phases of gonadal maturation in masu salmon (Amano et al., 1992, 1993).
These results suggest that the involvement of SL not only in gonadal development but also in energy mobilization is related to reproduction, since the biological events, during spawning process require a great deal of energy (Rand-Weaver et al., 1993; Johnson et al., 1997). Activated SL cells identified immunocytochemically, were also seen in spawning Oncorhynchus nerka, O. keta and O. tshawytscha (Olivereau and Rand-Weaver, 1994a, b) and in M. cephalus (Mousa and Mousa, 2000). The present immunocytochemical results received a good support from biochemical studies, which showed that SL levels have been shown to increase during the spawning migration of chum salmon Oncorhynchus keta (Kakizawa et al., 1995b).
The activity of mGnRH- and SL-ir cells seems to be decreased after spawning (spent) L. niloticus. In this stage, the synthetic activity of mGnRH- and SL-ir cells were greatly reduced and most of cells appeared to empty their secretory contents as indicated by decreased cells size and immunoreactivity with highly vacuolated cytoplasm. Similar immunocytochemical observations were obtained in O. niloticus (Mousa and Mousa, 1999), M. cephalus (Mousa and Mousa, 2000) and in O. bonariensis (Vissio et al., 2002). The marked decrease in the activity of GnRH together with SL-ir cells after spawning as reflected by the complete exhausted immunoreactivities of these hormones within cells, support the notion that SL under the stimulatory influence of GnRH are involved in spawning process.
In summary, our double immunostaining revealed a close association between mGnRH-ir fibers and SL-ir cells within the pituitary gland. We demonstrated also that the gradual increase in the synthetic activity of mGnRH cell bodies located in the mediobasal hypothalamus and SL-ir cells within pituitary was concomitant with the gonads maturation and this increase reached their maximum during ripening stages. After spawning, we showed a dramatic decrease in the synthetic of these cells. Taken together, the present findings suggest that SL under GnRH stimulation may be involved in the control of some biological events concerned with sexual maturation and spawning of L. niloticus.
ACKNOWLEDGMENTS
The authors are extremely grateful to Dr. H. Kawauchi and G. Tramu for donating the antisera used in this study. and to Prof. Shaaban Mousa (Klinik für Anaesthesiologie, Charité B Universitätsmedizin Berlin) for critical review of the manuscript.
REFERENCES
- Amano, M., K. Aida, N. Okumoto and Y. Hasegawa, 1992. Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary and GTH contents in the pituitary in female masu salmon, Oncorhynchus masou, from hatching through ovulation. Zool. Sci., 9: 375-386.
Direct Link - Amano, M., K. Aida, N. Okumoto and Y. Hasegawa, 1993. Changes in levels of GnRH in the brain and pituitary and GTH in the pituitary in male masu salmon, Oncorhynchus masou, from hatching to maturation. Fish Physiol. Biochem., 11: 233-240.
Direct Link - Collins, P.M., D.F.O. Neill, B.R. Barron, R.K. Moore and N.M. Sherwood, 2001. Gonadotropin-releasing hormone content in the brain and pituitary of male and female grass rockfish (Sebastes rastrelliger) in relation to seasonal changes in reproductive status. Biol. Reprod., 65: 173-179.
Direct Link - Company, R., J.A. Calduch-Giner, M. Mingarro and J. Perez-Sanchez, 2000. CDNA cloning and sequence of European sea bass (Dicentrarchus labrax) somatolactin. Comp. Biochem. Physiol. B, Biochem. Mol. Biol., 127: 183-192.
Direct Link - Company, R., A. Astola, C. Pendon, M.M. Valdivia and J. Perez-Sanchez, 2001. Somatotropic regulation of fish growth and adiposity: Growth Hormone (GH) and somatolactin (SL) relationship. Comp. Biochem. Physiol. C, Toxicol. Pharmacol., 130: 435-445.
Direct Link - Holloway, A.C. and J.F. Leatherland, 1997. The effects of N-methyl-D, L-aspartate and gonadotropin-releasing hormone on in vitro growth hormone release in steroid-primed immature rainbow trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol., 107: 32-43.
Direct Link - Johnson, L., B. Norberg, M.L. Willis, H. Zebroski and P. Swanson, 1997. Isolation, characterization and radioimmunoassay of Atlantic halibut somatolactin and plasma levels during stress and reproduction in flatfish. Gen. Comp. Endocrinol., 105: 194-209.
Direct Link - Kakizawa, S., T. Kaneko, S. Hasegawa and T. Hirano, 1995. Effects of feeding, fasting, background adaptation, acute stress and exhaustive exercise on the plasma somatolactin concentrations in rainbow trout. Gen. Comp. Endocrinol., 98: 137-146.
Direct Link - Kakizawa, S., T. Kaneko, T. Ogasawara and T. Hirano, 1995. Changes in plasma somatolactin levels during spawning migration of chum salmon (Oncorhynchus keta). Fish Physiol. Biochem., 14: 93-101.
Direct Link - Kakizawa, S., T. Kaneko and T. Hirano, 1996. Elevation of plasma somatolactin concentrations during acidosis in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol., 199: 1043-1051.
Direct Link - Kakizawa, S., T. Kaneko and T. Hirano, 1997. Effects of hypothalamic factors on somatolactin secretion from the organ-cultured pituitary of rainbow trout. Gen. Comp. Endocrinol., 105: 71-78.
Direct Link - Kaneko, T. and T. Hirano, 1993. Role of prolactin and somatolactin in calcium regulation in fish. J. Exp. Biol., 184: 31-45.
Direct Link - Kavanaugh, S.I., M.L. Powell and S.A. Sower, 2005. Seasonal changes of gonadotropin-releasing hormone in the Atlantic hagfish Myxine glutinosa. Gen. Comp. Endocrinol., 140: 136-143.
Direct Link - Bhandari, R.K., S. Taniyama, T. Kitahashi, H. Ando and K. Yamauchi et al., 2003. Seasonal changes of responses to gonadotropin-releasing hormone analog in expression of growth hormone/prolactin/somatolactin genes in the pituitary of masu salmon. Gen. Comp. Endocrinol., 130: 55-63.
PubMedDirect Link - Lu, M., P. Swanson and L. Renfro, 1995. Effect of somatolactin and related hormones on phosphate transport by flounder renal tubule primary cultures. Am. J. Physiol. Regul. Integr. Comp. Physio., 268: 577-582.
Direct Link - Mayer, I., M. Rand-Weaver and B. Borg, 1998. Effects of gonadectomy and steroids on plasma and pituitary levels of somatolactin in Atlantic salmon. Salmo salar. Gen. Comp. Endocrinol., 109: 223-231.
Direct Link - Mingarro, M., S. Vega-Rubin de Celius, A. Astola, C. Pendon, M.M. Valdivia and J. Perez-Sanchez, 2002. Endocrine mediators of seasonal growth in gilthead sea bream (Sparus aurata): The growth hormone and somatolactin paradigm. Gen. Comp. Endocrinol., 128: 102-111.
Direct Link - Mousa, M.A. and S.A. Mousa, 1999. Immunocytochemical study on the localization and distribution of the somatolactin cells in the pituitary gland and the brain of Oreochromis niloticus (Teleostei, Cichlidae). Gen. Comp. Endocrinol., 113: 197-211.
Direct Link - Mousa, M.A. and S.A. Mousa, 2000. Implication of somatolactin in the regulation of sexual maturation and spawning of Mugil cephalus. J. Exp. Zool., 287: 62-73.
Direct Link - Mousa, M.A. and S.A. Mousa, 2003. Immunohistochemical localization of gonadotropin releasing hormones in the brain and pituitary gland of the Nile perch, Lates niloticus (Teleostei, Centropomidae). Gen. Comp. Endocrinol., 130: 245-255.
Direct Link - Mousa, M.A., 2004. Seasonal changes in gonads of the Nile perch, Lates niloticus (Teleostei, Centropomidae) during the reproductive cycle in the Nile River. J. Egypt. Ger. Soc. Zool., 43: 23-44.
Direct Link - Muske, L.E., 1993. Evolution of gonadotropin-releasing hormone (GnRH) neuronal systems. Brain Behav. Evol., 42: 215-230.
Direct Link - Nguyen, N., M. Sugimoto and Y. Zhu, 2006. Production and purification of recombinant somatolactin β and its effects on melanosome aggregation in zebrafish. Gen. Comp. Endocrinol., 145: 182-187.
Direct Link - Olivereau, M. and M. Rand-Weaver, 1994. Immunocytochemical study of the somatolactin cells in the pituitary of pacific salmon, Oncorhynchus nerka and O. keta at some stages of the reproductive cycle. Gen. Comp. Endocrinol., 93: 28-35.
Direct Link - Olivereau, M. and M. Rand-Weaver, 1994. Immunoreactive cells in the pituitary of young, migrating and spent chinook salmon (Oncorhynchus tshawytscha). Fish Physiol. Biochem., 13: 141-151.
Direct Link - Onuma, T., H. ando, N. Koide, H. Okada and A. Urano, 2005. Effects of salmon GnRH and sex steroid hormones on expression of genes encoding growth hormone/Prolactin/Somatolactin family hormones and a pituitary-specific transcription factor in masu salmon pituitary cells in vitro. Gen. Comp. Endocrinol., 143: 129-141.
Direct Link - Parhar, S.I. and M. Iwata, 1994. Gonadotropin-releasing hormone (GnRH) neurons project to growth hormone and somatolactin cells in the steelhead trout. Histochemistry, 102: 195-203.
Direct Link - Peter, R.E., V.L. Trudeau, B.D. Sloley, C. Peng and C.S. Nahorniak, 1991. Actions of catecholamines, peptides and sex steroids in regulation of gonadotropin-II in the goldfish. Proceedings of the 4th International Symposium on the Reproductive Physiology of Fish, July 7-12, 1991, University of East Anglia Printing Unit, Sheffield, pp: 30-34.
- Rand-Weaver, M., P. Swanson, H. Kawauchi and W.W. Dickhoff, 1992. Somatolactin, a novel pituitary protein: Purification and plasma level during reproductive maturation of coho salmon. J. Endocrinol., 133: 393-403.
Direct Link - Rand-Weaver, M., T.G. Pottinger and J.P. Sumpter, 1993. Plasma somatolactin concentrations are elevated by stress. J. Endocrinol., 138: 509-515.
Direct Link - Rand-Weaver, M., T.G. Pottinger and J.P. Sumpter, 1995. Pronounced seasonal rhythms in plasma somatolactin levels in rainbow trout. J. Endocrinol., 146: 113-119.
Direct Link - Stefano, A.V., P.G. Vissio, D.A. Paz, G.M. Somoza, M.C. Maggese and G.E. Barrantes, 1999. Colocalization of GnRH binding sites with gonadotropin-somatotropin-somatolactin- and prolactin-expressing pituitary cells of the pejerrey, Odontesthes bonariensis, in vitro. Gen. Comp. Endocrinol., 116: 133-139.
Direct Link - Taniyama, S., T. Kitahashi, H. Ando, M. Kaeriyama, Y. Zohar, H. Ueda and A. Urano, 2000. Effects of gonadotropin-releasing hormone analog on expression of genes encoding the growth hormone/Prolactin/Somatolactin family and a pituitary-specific transcription factor in the pituitaries of prespawning sockeye salmon. Gen. Comp. Endocrinol., 118: 418-424.
Direct Link - Tsai, P.S. and J.T. Jones, 2005. Steroid-induced changes in the morphology of GnRH neurons in male leopard frogs, Rana pipiens: correlation with plasma gonadotropin and gonadal size. Gen. Comp. Endocrinol., 141: 152-160.
Direct Link - Vissio, P.G., A.V. Stefano, G.M. Somoza, M.C. Maggese and D.A. Paz, 1999. Close association of gonadotropin-releasing hormone fibers and gonadotropin, growth hormone, somatolactin and prolactin expressing cells in pejerrey, Odontesthes bonariensis. Fish Physiol. Biochem., 21: 121-127.
Direct Link - Vissio, P.G., L. Andreone, D.A. Paz, M.C. Maggese, G.M. Somoza and C.A. Strussmann, 2002. Relation between the reproductive status and somatolactin cell activity in the pituitary of Pejerrey, Odontesthes bonariensis (Atheriniformes). J. Exp. Zool., 293: 492-499.
Direct Link - Weber, G.M., J.F. Powell, M. Park, W.H. Fischer and A.G. Craig et al., 1997. Evidence that gonadotropin-releasing hormone (GnRH) functions as a prolactin-releasing factor in a teleost fish (Oreochromis mossambicus) and primary structures for three native GnRH molecules. J. Endocrinol., 155: 121-132.
PubMedDirect Link - Zhu, Y. and P. Thomas, 1996. Elevations of somatolactin in plasma and pituitaries and increased a-MSH cell activity in red drum exposed to black background and decreased illumination. Gen. Comp. Endocrinol., 101: 21-31.
Direct Link - Zhu, Y. and P. Thomas, 1998. Effects of light on plasma somatolactin levels in red drum (Sciaenops ocellatus). Gen. Comp. Endocrinol., 111: 76-82.
Direct Link