
Magdy M. Mohamed
Department of Chemistry, Taif Teachers�s Collage, 1070 Taif, Saudi Arabia
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 7 | Page No.: 1046-1059







ABSTRACT
This study was conducted to evaluate the immunogenicity and protective efficacy of B. abortus protein antigen. A gene that encode 0.85 kbp was isolated from B. abortus genome λgt11 expression library by probing with a Polled Human Abs (PHAbs) infected with B. abortus or B. melitensis. Sequence analysis of the cloned gene revealed the presence of an Open Reading Frame (ORF) of 259 amino acids encoding for a protein with calculated molecular weight of 23 kDa (Bp23). The predicted amino acid sequences of this gene shows 83% similarity with B. abortus outer membrane 26 kDa protein (Omp26) previously sequenced. The recombinant B. abortus protein was identified by human sera infected with either B. abortus or B. melitensis, but not healthy persons as shown by Western blotting. Therefore, the availability of this recombinant protein and the identification of the antigenic determinant recognized by human Ab will allow the evaluation of its potential protective activity for the development of subcellular vaccines against brucellosis. Moreover, the use of this recombinant protein in Western blotting allowed differentiation between vaccinated from naturally infected camels. Hyperimmune rabbit serum against Bp23 recombinant protein could detect naturally infected animals with different specificities, whereas represented in camels (100%), cows (54%) and sheep (33%), but not detected at all in brucellosis free camels. These data indicated that the new isolated Bp23 recombinant protein might be of value as a vaccine candidate for treatment of human brucellosis and as an antigen for serological diagnosis of brucellosis in different animals as well.
PDF Abstract XML References Citation
How to cite this article
Magdy M. Mohamed, 2007. Molecular Characterization and Immunogentic Properties of a Group 3 Membrane Protein Isolated from B. abortus Infection. Journal of Biological Sciences, 7: 1046-1059.
DOI: 10.3923/jbs.2007.1046.1059
URL: https://scialert.net/abstract/?doi=jbs.2007.1046.1059
DOI: 10.3923/jbs.2007.1046.1059
URL: https://scialert.net/abstract/?doi=jbs.2007.1046.1059
INTRODUCTION
Microorganisms belonging to the genus Brucella are intracellular coccobacilli, small, nonmotile, gram negative bacteria, that are able to infect a broad range of wildlife and domestic animal species as well as humans (Young, 1983). The genus Brucella is classified into six species: three major (Brucella melitensis, B. abortus and B. suis) and three minor, (B. ovis, B. canis and B. neotomae). In addition, several biovars are included in the three major species. The six Brucella species and their biovars are currently differentiated by pathogenicity, animal host preference infections, serotyping, phage typing, dye sensitivities, culture and metabolic properties (Alton et al., 1988). Among these different biotypes B. abortus and B. melitensis are the most widespread species for infection of camels (Young, 1995), most causative agent of bovine brucellosis (Young, 1983) and most pathogenic species for infection of humans (Corbel, 1997; Fallatah et al., 2005). Animal brucellosis induced abortions and reduced fertility in camels (Yagupsky, 1999) causes severe economic losses in underdeveloped and developing countries in which animal husbandry is an important industry (Blasco and Diaz, 1993). In addition, severe human brucellosis characterized in its acute phase by undulant fever and in its chronic phase by localization of the pathogen and damage of different organs (Corbel, 1997; Roop et al., 2004). If localized in the brain or the heart, this can result in fatal meningitis or fatal endocarditis, respectively (Fallatah et al., 2005). Furthermore, all these Brucella species are listed as potential bioweapons by the Centers for Disease Control and Prevention (Kaufmann et al., 1997; Kortepeter and Parker, 1999). This is due to the highly infectious nature of three major species. In fact they can be readily aerosolized and an outbreak would be difficult to detect because the initial symptoms are easily confused with those of influenza.
Brucellosis is an endemic zoonotic disease in many areas of Saudi Arabia mainly in the western and southern province, due to a traditional custom fostered by the nomadic heritage and dietary habits of the people, a high cross transmission of Brucella sp. from domestic animals including sheep and camels to humans (Al-Eissa, 1999).
However, camel husbandry is vital for numerous pastoralist groups at Taif area in Western Saudi Arabia. Therefore, infected camels are the main source of human Brucellosis infection. Currently, seroprevelance of Brucellosis in humans at these regions increased from 4 to 15% within the last 20 years (Almuneef et al., 2004), due to increase the rate of camel’s raw milk ingestion for treatment of some diseases. Meanwhile, the overall Brucellosis seroprevalance was also increased from 3 to 22% in camels (Radwan et al., 1995; Abbas and Agab, 2002). Results of laboratory tests indicated that a significant proportion of the population in the southern region (19.2%) had serological evidence of exposure to Brucella antigen and 2.3% had active disease (Radwan et al., 1995). However, recorded data about the main risk factors proved that 45% of human brucellosis population from Northern area of Saudi Arabia was due to ownership or direct contact with domestic camels and 75% was from the ingestion of raw products of animal or unpasteurized milk (Almuneef et al., 2004). The disease present in both sexes and in all ages (Alballa, 1996). Many attempts had been done to evaluate a human vaccine, control brucellosis infection in large camel herd in Saudi Arabia and prevention of cross transmission to humans (Fallatah et al., 2005; Al-Tawfiq, 2006).
Brucella infection is treated with either a combination of antibiotics or attenuated strains (Aguirre et al., 2006); however, in its chronic phase, eradication is difficult since Brucella sp. are intracellular pathogens, which puts them out of reach of humoral immunity and several antibiotics (Kohler et al., 2003; Roop et al., 2004). Vaccination against brucellosis is commonly performed by using living attenuated Brucella strains. These vaccines have several disadvantages, among them lack of a safe, their capacity to produce underlying infections in animals (Abbas and Agab, 2002) and humans (Young, 1983), sometimes becomes pathogens for human (Cloeckaert et al., 2002), its use is prohibited in countries free of it (Jimenez et al., 1994; Lucero et al., 2006) and to elicit antibodies against the lipopolysaccharide (LPS; Cloeckaert et al., 2002) which interfere in the differential diagnosis between vaccinated and infected animals. Therefore, the development of new vaccines and effective therapeutic agents is one of the principal aims in brucellosis research. Numerous attempts to develop killed vaccines that are as effective as attenuated Brucella strains have met with limited success (Cassataro et al., 2005). Subcellular vaccines as protective antigens (Ags) that could avoid the drawbacks of live attenuated vaccines, being safer and not interfering with immunodiagnosis, are selected. Therefore, identification of Brucella sp. antigens with the capacity to elicit a protective immune response is of great interest for the development of subcellular vaccines that do not have the drawbacks of living attenuated vaccines.
The Brucella cell wall consists of a groups of immunogenic and protective antigens which characterized as an outer membrane proteins (Omps) of Brucella or Brucella group antigens (Ags) that focused for vaccine applications and potential diagnostic purposes (Jimenez et al., 1994). These proteins are categorized to seven Brucella Omps Ags according to their molecular weight by using MAbs (Pugh et al., 1991; deWergifosse et al., 1995). Three major Omps classified into two groups; group 2 porin protein Ags sharing a high degree of identity (>85%; Bowden et al., 2000; Estein et al., 2003) with a molecular weight approximately ranged from 36 to 38 kDa, (Omp 2b; Ficht et al., 1989) and group 3 proteins that have two different molecular weights; 25 to 27 kDa Omps (Omp 25; Vizcaino et al., 2001) and 31 to 34 kDa Omps (Omp31; de Wergifosse et al., 1995). Four minor Omps classified into two groups; designed as group 1 Ags that have an approximately molecular mass of 94 kDa (Omp94; Lindler et al., 1996; Halling et al., 2005) and group 4 Ags with three different molecular weights of 10, 16 and 19 kDa Omps (10, 16 and Omp 19, respectively; Cloeckaert et al., 2002). Previous reports proved that antibodies against the group 3 Ags have been detected in mice naturally and experimentally infected with B. melitensis (Zygmunt et al., 1994) and in camels infected with B. abortus and seems to be highly accessible for sheep infected with smooth B. suis (S B. suis; Bowden et al., 1995). Accordingly, the Brucella sp. group 3 Omps might be an interesting candidate for the development of a subcellular vaccine against human as well as domestic camels brcellosis caused by B. melitensis and B. abortus strains.
Our main objective in this present research was to isolate genes coding for surface proteins, which could be useful as protective, therapeutic or diagnostic tools for brucellosis infection. In this study, cloning, nucleotide sequence and expression of the gene coding for the 0.85 kbp from B. abortus that may be useful as a vaccine candidate Ag and/or diagnostic agent.
MATERIALS AND METHODS
Bacterial strains: The B. abortus and B. melitensis strains used in this study were isolated from milk of infected camels by the Brucella Culture Collection staff, a branch of Agriculture and Water Ministry at Taif, Saudi Arabia and classified as rough (R) Brucella strains by standard methods (Radwon et al., 1995). Working culture of Brucella strains were grown and maintained on tryptic soy agar (Gibco-BRL/Life Technologies, Eragny, France) supplemented with 0.1% (wt./vol) yeast extract (Difco Laboratories, Detroit, Mich.) for B. melitensis and with 5% horse serum (Gibco-BRL/Life Technologies, Eragny, France) for B. Abortus. Characterization of cultures was performed by standard procedures (Alton et al., 1988). Stock Brucella cultures were stored in trypticase broth supplemented with 20% glycerol and stored at -80°C (Guilloteau et al., 1999). Escherichia coli JMI09 and Y1090 cell strains (Promega, Madison, WI) used in this study were cultured overnight on Luria-Bertani (LB) mediurn containing 50 μg of ampicillin mL-1.
Genomic DNA preparation and construction of Brucella genomic library: Total genomic DNA was extracted and purified from isolated B. abortus and B. melitensis as described previously (Sambrook et al., 1989). The λgt11 B. abortus gemonic library was prepared in King Fahed institute, Jeddah, Saudi Arabia by digestion of B. abortus chromosomal DNA with Sau 3A1 (Promega, Madison, Wis.). Briefly, B. abortus genomic DNA was partially digested for 30 min at 37°C with Sau3A1, the enzyme concentration giving the highest percentage of fragrnents ranging from 0.5 to 23 kb. DNA fragments were purified by elecroelution of 1% agarose gels then EcoRI linkers were added to the fragment and ligated with T4 DNA ligase (Promega, Madison, Wis) into λgt11 EcoRI cloning site that performed as described by Sambrook et al. (1989). B. melitensis library was also constructed in λgt11 by similar method described above. Recombinant phage DNA were packaged in vitro with the Packagene System (Promega, Madison, Wis) and libraries were titrated for determination the number of PFU by infection of Escherichia coli JM109 cells and spread onto plates containing Luria-Bertani (LB) medium supplemented with ampicillin (50 μg mL-1) as illustrated by the manufacturer.
Sera and Ab preparation: Several sources of human and animals infected serum samples were used in this experiment. Twenty unvaccinated human brucellosis sera were obtained from outpatients of the brucellosis Department at Taif National Hospital (Taif, KSA). Among them twelve sera from infected patients with B. abortus and eight sera from patients infected with B. melitensis. Two sera from normal subjects used as a negative control and six serum samples were collected from King Faisal National Hospital termed human sera Ab. Three of them collected from patients infected with B. abortus and other three sera collected from patients infected with B. melitensis. Those infected patients were confirmed by positive culture for each strain as a routine work in the hospital. Pooling of first twenty serum samples collected from Taif National Hospital were passed on a column containing protein A/G (Pierce) for purification of immunoglobulin G (IgG) as suggested by the manufacturer. This Ab preparation represents a pool of sera with both Brucella sp. and designated as Pooled Human Ab (PHAb). IgG preparation from other six serum samples were also prepared separately as described above. in addition, five serum samples from camels, 7 serum samples from cows and 9 serum samples from sheep infected with B. abortus and two serum samples from vaccinated camels with S19 B. abortus, two serum samples from camels infected with B. melitensis and two serum samples from vaccinated camels with Rev1 B. melitensis were obtained from Disease Prevention and Control Unit (DPCU), at Ministry of Agriculture and Water branch at Taif; Saudi Arabia. Five serum samples from healthy camels (negative by complement fixation test) were obtained from a Brucellosis free herd kindly prepare by Dr. Zakaria, H. from the field with helping of some specialists in animal resources Department Ministry of Agriculture and Water. IgG fraction of anti -23 kDa protein sera were obtained from immunized rabbits with a purified protein extract prepared as follows: Whole cell lysate of E. coli carrying the plasmid that produces fussion 23 kDa protein (pGEX4T-2-23) was subjected to 12.5% SDS-PAGE gels as described by Laemmli (1970) and transferred to nitrocellulose membrane. A Western blot using HPAb sera described above was made with part of the nitrocellulose membrane as described by Towbin et al. (1979) and the corresponding band on the gel (about 50 kDa fussion protein) was sliced, purified by electroelution and concentrated by protein filter tube (Millibore, USA). Fussion protein digested with factor X and purified on GST-column to separate GST protein from recombinant 23 kDa protein. Purified protein mixed with an equal volume of incomplete freund adjuvant (Sigma Chemical Co.) for repeated immunization of rabbit. Hyperimmune rabbit sera was collected from the rabbit by heart puncture and pass over protein A column according to manual (Sambrook et al., 1989).
Library plating and immunoscreening: A bacteriophage suspension (10 μL; approximately 7x03 PFU) in phage dilution buffer (20 mM Tris-HCl, 100 mM NaCI, 10 mM MgSO4; pH 7.4) was incubated for 20 min at 37°C with 0.6 mL of an overnight E. coli JM109 culture grown at 37°C in LB media that supplemented with 50 mg ampicillin, 10 mM MgSO4 and 2% maltose per liter. Platting of E. col infected with λgt11 bacteriophages, overlaying of nitrocellulose membrane, blocking of membranes with 5% free fat milk, probing the membrane with HPAb and color development of expressed protein was done according to Sambrook et al. (1989). The strongest positive bacteriophages were repeated several times until plaque lifts showed more than 99% immunoreactive plaques.
PCR, subcloning and plasmid preparation: B. abortus and B. melitensis DNA inserts were recovered from the isolated bacteriophages by using PCR as described previously (Mullis and Falnoona, 1987) on the purified bacteriophage. Forward and reverse universal λgt11 primers (Stratagene, La Jolla, Calif.) were used under the following conditions. The reaction mixture (100 μL) contained 0.1x103 PFU of recombinant phage, 100 pmol of each primer, 200 mM of each deoxynueleoside triphosphate (dNTPs; Phannacia, Sweden) and 2 U of Taq polymerase (Perkin-Elmer, Cetus, Emeryville, Calif.) with the appropriate 10x polymerase buffer. The reaction mixture was preheated for 10 min at 95°C and then 35 cycles (denaturation at 95°C for 60 sec annealing at 55°C for 90 sec and polymerization at 72°C for 2 min) were performed in a thermocycler (Perkin-Elmer Cetus, Emeryville, Calif). The reaction was completed by heating for 10 min at 72°C as elongation step, then kept at 4°C until used. Different sizes of DNA fragments obtained were subjected to EcoRI digestion, phenol/chlorophorm extraction, ethanol precipitation, then resuspended into 20 μL TE buffer. Purified DNAs were subcloned into a pGEM-5Zf+ vector (Promega-Biotech, Madison, WI) by T4 DNA ligase (Promega) as described previously (Ausubel, 1989). Recombinant plasmids were transformed and propagated in E. coli JM109 as described by Sambrook et al. (1989). Plasmid DNA purification was performed on a Maxi-prep DNA purification kit (Promega) according to the manufacturer instruction.
DNA sequencing and protein analysis: DNA sequence information was obtained from sequencing of both strands of the inserts by primer-directed dideoxy sequencing of double-stranded templates with Sequenase version 2.0 (US Biochemicals, Cleveland, Ohio) using [35S] as isotope (Amarsham; USA). The forward and reverse primers were a universal primers for pGEM plasmid (T7 and SP6, respectively; Stratagene, La Jolla, Calif.) used for sequencing. Sequence was performed using 6 μL of purified DNA product per reaction, 4 μL of forward or reverse primer, 8 μL of primex buffer as described by Sanger et al. (1977). DNA sequence data obtained from sequencing gels were analyzed by the DNA Strider 1.1 program (Marck, 1988). Sequence analyses were performed also on a SUN Spareserver 10/41, FastA, TFastA, PepPlot and Motifs programs. DNA and amino acid sequences were used to search the DNA and protein databases at the National Center for Biotechnology Information (NCBI), National Library of Medicine, Washington, DC., by using the BLAST algorithm (Altschul et al., 1990).
Protein expression, gel electrophoresis and westerm blotting: For expression of the encoded protein, recombinant pGEM-DNA for Brucella insert was purified by using Qia Quick purification kit (Qiagen) as described by the manufacturer. Open Reading Frames (ORF) of encoded genes was generated by constructed two sets of primers; sense primer 5`-GAATTCACTTGCTAGCCGGAC-3` and antisense primer 5`-GGATCCTCGGAGGGATTACTTA-3` for B. abortus containing EcoRI and BamHI cloning site. DNA fragment was generated by PCR of 100 ng DNA with PCR conditions as described above. PCR fragment was recovered and digested with EcoRI and BamHI and ligated into pGEX4T-2 vector. Following transformation of E. coli Y1090 competent cells, colonies were examined by cracking gel electrophoresis to identify which carries the recombinant plasmid. A single bacterial colony containing the recombinant plasmid was grown in a culture and induced with 0.5 mM IPTG to induce recombinant protein expression. Cells were harvested at 2 h intervals by centrifugation at 3000xg and resuspended in fussion protein extraction buffer (Tris-HCl, 50 mM; NaCl, 15 mM; EDTA pH 8.5; Triron-X 100, 1%; PMSF 1 mM). Cells were then lysed by sonication for 2-3 min at 20 kHz with 30 sec intervals using 0.2 nm microtipe and centrifugatied in a sorval SS34 rotor at 1200 rpm at 4°C for 10 min. The pellet was then resuspended in the fussion protein extraction buffer and analyzed together with the supernatant on 15% SDS PAGE gel as described by Laemmli (1970). Western blotting was performed according to the method of Towbin et al. (1979) on the supernatant.
Southern blotting and hybridization: Fragments generated from Sau3A1 restriction enzyme digestion of 1.5 pg of chromosomal DNA Brucella sp. fractionated on 0.7% agarose gel were transferred to Nylon membranes (Schleicher and Schuell, Keene, N.H) and hybridized at 42°C as suggested by the manufacturer with 0.85 Kbp DNA labeled with [32P]. The membranes were washed under high-stringency conditions at 65°C in lx Denhardt's solution (0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.02% bovine serum albumin, 2xSSC (20xSSC is 3 M NaCI and 0.3 M sodium citrate) and three 30 min washes in 0.1xSSC-0.1% SDS by using conventional method according to Sambrook et al. (1989). The membranes were dried and autoradiographed on X-0 mat AR film (Kodak, Rochester. NY).
RESULTS
Isolation, identification and cloning of a B. abortus insert gene: In a first screening, about 250,000 λgtl1 plaques from each B. abortus and B. melitensis libraries were probed separately with a pooled of HPAb. Five positive plaques were identified as strongest positive immunogenic dots (two from B. abortus and three from B. melitensis). The isolated five plaques were purified by repeated plating and probing. The purified bacteriophages were again plated by spotting into two plates containing Y1090 bacteria as in spot blotting technique (Magdy, 1988). One plate was probed with pooled three human sera Ab directed to B. melitensis and the other plate was probed with pooled three human sera Ab directed to B. abortus. Only one plaque (No. 4) that described as a highly immunoreactive bacteriophage with both separate sera Abs which was used for further characterization. To recover the DNA fragment, the purified bacteriophage was used as a template in a PCR reaction with primers flanking the cloning site. The PCR DNA fragments of these five plaques were run on 1% agarose gel to detect their sizes (Fig. 1a). Interestingly, two B. melitensis DNA fragments (No. 1, 3) were almost in the same size as two DNA fragments of the B. abortus (No. 4, 5; 0.85 kbp), however, B. melitensis DNA (plaque No. 2) size is about 3.7 kbp. A 0.85 kbp DNA fragment was purified via EcoRI digestion of PCR product and subcloned into the pGEM5Zf+ vector EcoRI cloning site. A recombinant vector transformed into Y1090 bacteria, then amplified, digested and run on 1% agarose gel electrophoresis (Fig. 1b), which represented DNA fragment with ~ 850 bp.
Nucleotide sequence and amino acid analysis: A complete coding sequence was carried out for clone No. 4 from both strands of pGEM5Zf+ containing DNA fragment by using universal T7 and SP6 primers flanking the parental 850 bp fragment. On one strand, a single open reading frame (ORF) of 777 bp was identified (Fig. 2) with an ATG starting codon and a TAA stop codon and could code for 259 amino acid protein. Indeed, DNA insert contains a seven nucleotide sequence that is homologous to a putative Shine-Dalgarno consensus sequence (TAGGAGA) that was found upstream from the putative initiation codon (position 4). A C+G rich region that could function as transcription terminator sequence was detected at 4 bp down stream of the TAA stop codon. The B. abortus amino acids were predicted by translation of the nucleotide sequence through the current Swiss Prot, EMBL and GenBank databases. The calculated molecular mass of the predicted encoded protein was 23 kDa (Bp23). A search for amino acid sequence homologies revealed that its amino acid sequence exhibits 83.0% identity with 26 kDa Omp (Omp26) of B. abortus and 75.0% identity with Omp25 B. abortus or B. melitensis (Omp25) and much lower identities recorded with other proteins. Alignment of the predicted protein sequence with B. abortus Omp26 protein is shown in Fig. 3.
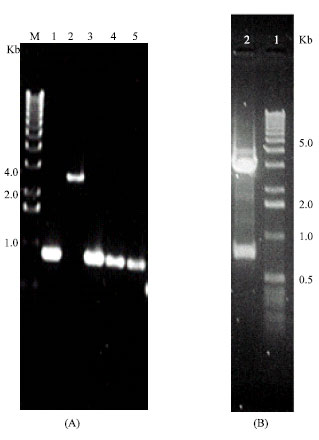 | |
| Fig. 1: | 1% agarose gel electrophoresis: Panel (A) represents a PCR fragment of isolated plaques from B. abortus λgt11 library (lanes 1, 2) and B. meletinsis (lanes 3, 4, 5). M is a molecular weight marker, (1Kbp DNA ladder Gibco). Panel (B) showing the EcoRI digestion of pGEM5Zf+ vector containing DNA fragment purified from lane 4 that showing a high immunoreactivity for PHAb. |
PAGE analysis and western blot: Characterization of Bp23 protein was carried out by subcloned the coding sequence in the expression vector pGEX4T-2 and the protein was expressed in E.coli as a fussion protein with Sj26 (28 kDa). One, three and five hours post induction of pGEX-23 bacterial cells were centrifuged and resuspended in protein resuspension buffer. After sonication, pellets and supernatant of lysed cells were run on 15% SDS-PAGE. The protein was present mostly in the pellets than supernatant fractions. Figure 4 shows a band of about 51 kDa in both fractions, thus the expected size for protein is about 23 kDa that agree with the predicted size.
 | |
| Fig. 2: | Nucleotide and amino acid sequence of B. abortus insert gene. The putative ribosome binding site (rbs) is underlined. The nucleotide and amino acid sequence numbered at the left. The ORF is shown with the deduced amino acid sequence below the third nucleotide in single-letter |
 | |
| Fig. 3: | Alignment of predicted amino acid sequence of B. abortus 23 kDa protein, with B. abortus Omp26 that showing 83% homology, dashes are missing amino acids, * is different amino acids |
In order to detect the existence of antibodies in human sera against the Bp23 protein, Western blot analysis was carried out on purified recombinant Bp23 protein as an antigen against infected human sera (Fig. 5) with either B. abortus (lanes 2-4) or B. melitensis (lanes 5-7). There is a high immuno-reactivity with a protein at a band of 23 kDa molecule with both cases. The recombinant Bp23 bacterial protein was analyzed as a positive control against hyperimmune rabbit sera for the existence of an immunogenic activity (lane 1). Furthermore, Western blot analysis of negative infected sera did not show any immunogenic reactivity with the recombinant Bp23 protein (lane 8).
Western blot strips analysis of sera from naturally infected (which were positive by classic standard serology), vaccinated and uninfected camels against purified recombinant Bp23 antigen protein (Fig. 6) showed that infected animals with either B. abortus and B. melitensis induced antibodies against the 23 kDa protein, as represented by strongly immunoreactive bands (lanes 2 and 3), respectively, while very week reaction was detected of vaccinated camels with either S19 or Rev1 (lanes 5 and 6), respectively where the presence of a 23 kDa band could be hardly observed.
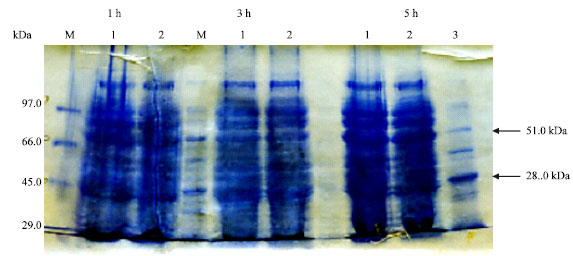 | |
| Fig. 4: | SDS-PAGE from induction experiment of isolated B. abortus 23 kDa protein in the expression vector pGEX4T-2 Lanes 1 and 2 are pellet lysate and supernatant respectively, lane 3 is GST-28 kDa as a marker. M is prestained molecular weight marker (Gibco) |
 | |
| Fig. 5: | Western blot for reactivity of cloned B. abortus with various human sera. The recombinant 23 kDa protein antigen was used on the Western blot. The immunoreactive protein is indicated by the labeled arrowhead. Lane 1; hyperimmune rabbit Ab sera against 23kDa protein as a positive control; lanes 2-4, human serum from patient infected with B. abortus; lane 5-7. human serum from patient infected with B. melitensis; lane 8 is healthy human serum. The dilution of all the primary Abs was 1/1,000 |
 | |
| Fig. 6: | Western blot analysis of B. abortus proteins. Blots were loaded with a 23-kDa recombinant protein and probed against serum sample from a health camels (lanes 1, 4) or infected with B. abortus (lane 2), B. melitensis (lane 3) or vaccinated camels against B. abortus; S19 strain (lane 5) or B. melitensis, Rev 1 strain (lane 6) |
 | |
| Fig. 7: | Western blot analysis of naturally infected sera from camels cows and sheep to detect the immunoreactivity of infected animal serum (Ag) against prepared hyperimmune rabbit serum for 23 kDa protein (Ab) of B. abortus. A represents infected serum running on PAGE and stained with commassiee brilliant blue dye and B is a western blot of corresponding PAGE using hyperimmune rabbit serum prepared as in materials and methods |
Serum from healthy camels as a controls did not show any existence of 23 kDa band (lanes 1 and 4). The present experiments show that camels naturally infected with either B. abortus or B. melitensis produced an Abs that might be directed to same molecular weight antigen.
Analysis of sheep, bovine and camel sera: To evaluate that anti-23 kDa sera (hyperimmune rabbit sera) has immunogenicity against putative Bp23 antigen in animals infected sera in vivo. Serum samples from 9 sheep, 7 cows and 5 camels belonging to infected flocks with B. abortus strain and 5 normal control sera were randomly chosen from different herd. They were ran on 12.5% poloyacrylamide gel (Fig. 7) and stained with Commassie brilliant blue dye as in upper trays in panels (A, B, C and D). Existence of putative antigen in animals sera was tested by Western through reactivity with the hyperimmune rabbit sera as shown in the lower trays in panels (A, B, C and D). Data in Fig. 7 demonstrated that most of the reactivity seen at the 23 kDa position where 100% of infected camels (5/5), 57% of infected cows (4/7) and 33% of infected sheep (3/9) were recognized by this prepared hyperimmune rabbit sera against Bp23 protein. In addition, 5 control serum samples from camels were none reactive with prepared hyperimmun rabbit sera.
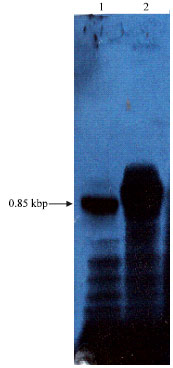 | |
| Fig. 8: | Southern blot hybridization of digested chromosomal Brucella DNAs of B. abortus in lane 1 and B. melitensis in lane 2 with [32P] labeled fragment of 0.85 kbp nucleotide probes. Approximately 1.5 pg of each chromosomal DNA was digested with EcoRl. DNAs in each lane |
Southern blot: The DNA probe of 0.85 kbp PCR fragment was hybridized with Sau3A1-digested chromosomal DNA fragments of B. abortus and B. melitensis as represented in Fig. 8. There is a major band at 0.85 kbp size of both strains indicating that this gene were located on same Sau3A1 fragments and did not vary in size between the chromosomes of different Brucella sp. It was not surprising that this gene might be highly homologous at the genomic level of both B. abortus and B. melitensis.
DISCUSSION
Characterization of 0.85 kbp gene and BP23 protein: In order to identify antigens which provoke a serological immune response in hosts infected with Brucella bacteria and therefore might be used for diagnostic or vaccination purposes. A B. abortus and B. melitensis chromosomal genome were digested with Sau3A1 and cloned into a λgt11 expression library which expressed a protein recognized by a pooled human sera infected with B. melitensis and/or B. abortus. A 0.85 kbp DNA fragment of B. abortus was identified and subcloned into pGEX4T-2 expression vector which expressed a protein recognized by animals or human serum infected with Brucella bacteria (Fig. 5, 6 and 7). The size of DNA fragment was agreed with that located in the chromosomal DNA of B. melitensis digested with Sau3A1 (Fig. 8), indicating that locus of the gene is similar in both Brucella sp. and its expressed protein is consistence. A computer analysis revealed that this gene represents a complete coding sequence in this fragment (Fig. 2) starting with ATG codon and ending within frame stop codon TAA. The C-terminal end of the protein was a phenyalanine that seems to be a signal sequences or transmembrane domains essential for efficient translocation of proteins across the bacterial outer membrane. However no consensus promoter sequence was found upstream from the putative start codon. A Shine-Dalgarno sequence or ribosomal binding site (TAGGAGA) was found 4 bp upstream from the first ATG on the DNA strand encoding gene. Upon inspection of the complete coding sequence of isolated B. abortus DNA fragment revealed that the predicted amino acid sequence derived from only one ORF with calculated molecular mass of 23 kDa. A similar situation has been observed for other Brucella genes (Larsson et al., 2005), in which most of the Brucella sp. DNA strand contains only one ORF of significant length. As reported for other Brucella Omps (Pugh et al., 1991) no region of high overall hydrophobicity were found except for first 15 N-terminal residues (Fig. 2).
A search for homologies of the newly identified gene sequence or the deduced Bp23 protein sequence with other sequenced protein in sequence libraries revealed 83% identity with a previously described antigen of B. abortus designed Omp26 (Fig. 3), an antigen that belongs to group 3 Ag that expressed protein with molecular mass of 26 kDa (Rossetti et al., 1996) and 75% identity with B. abortus Omp25 (de Wergifossee et al., 1995). Both Omp25 and Omp26 proteins are belonging to the Omp group 3 Ags that were present in membrane blebs of both B. abortus and B. melitensis. Comparison of our data with both proteins demonstrated that Bp23 may be present on similar blebs and lends credence to classify this Ag as one of the Omp group 3 Ags. Other cloned genes of Brucella sp. that express proteins with similar molecular masses characterized by other research groups showed very low or no significant homology with Bp23. Lindler et al. (1996) cloned a 28 kDa outer membrane protein but the predicted amino acid sequence is mostly different from the one reported here (37%); the 31 kDa protein cloned by Vizcaino et al. (1996) does not show any similarity with BP23 either. The protein (Omp17) characterized by Goldbaum et al. (1993) and sequenced by Hemmen et al. (1995) does not show any sequence similarities with isolated gene although sharing some properties with BP23 as reaction against brucellosis sera from sheep and patients but not against vaccinated animals. Moreover, nucleotide and amino acid sequences of BP23 showed no homology with the 29 kDa protein (Cloeckaert et al., 2002) of different Brucella sp. or with other bacterial proteins in a data bank. This data clearly reflect that Bp23 is a newly identified gene and strongly belonging to Omp group 3 Ags of B. abortus where antigens of molecular mass approximately ranging from 23 to 34 kDa were designated as Omp group 3 Ags (Verstreate et al., 1982; Larsson et al., 2005; Patrick et al., 2005).
The predicted size of the protein agreed very well with the size of the protein observed on Western blots of serum from naturally infected animals with B. abortus probed with hyperimmune rabbit sera against Bp23 recombinant protein (Fig. 7 panels A, B and C), where a 23 kDa putative protein band was identified. This data together with Western blot of recombinant Bp23 protein against infected human serum with either B. abortus or B. melitensis (Fig. 5 lanes 2-7) indicating that the natural Brucella antigen and the recombinant E. coli-produced protein are very similar. Moreover, the corresponding antigen in B. melitensis and B. abortus seemed to be very similar because of their molecular masses and antigenicities.
A vaccine candidate antigen for human brucellosis: Due to epidemic potential of brucellosis affecting different parts of the human body (Ersoy et al., 2005), absence of a human vaccine, drawback of current vaccine strains in terms of safety and the efficiency of aerosol infection, this airborne pathogen is classified as a biosafety level 3 pathogen and considered to be a potential bioterrorism agent. Several attempts have been made to control human disease by vaccination with attenuated Brucella strains (Schurig et al., 2002). However, practical acceptance of these preparation has been very limited, mainly because of the unacceptable local inflammatory reactions elicited by them, together with their limited protection (Huang et al., 2005). Clearly, there is a need to develop effective medical protective vaccine against it for better human vaccination for individual who may occupationally exposed to Brucella sp. and for persons who consume unpasteurized dairy products specially in areas where this bacteria is endemic, controlling of brucellosis in domestic animals to reduced animal morbidity and therefore decrease the incidence of human infection and may be for brucellosis eradication.
Omps are research area of intense interest for the development of an effective subunit vaccine against brucellosis that characterized as potential immunogenic and protective antigens. Previous literatures demonstrated that many Omps Brucella proteins to which infected animals developed appropriate immune responses may play a determinant role in the host-acquired protective immunity to brucellosis and it is also possible that these antigens are able to develop another immune response that is important for protection against Brucella infection (Lindler et al., 1996; Cassatro et al., 2005). Meanwhile, Stevens et al. (1995) reported that, Ab recognition of B. abortus or B. melitensis Omp proteins may offer some protection against Brucellosis and these data are in concordance with further reports which proved that humoral immune recognition following smooth (S) Brucella infection is partially protective but the highest level of protection is achieved with serum plus T cells transformation (Lindler et al., 1996). In contrast, immunity to infection with rough (R) Brucella is most likely quite different from protection against challenge with S species. Jimenez et al. (1994) showed that passive transfer of serum Ab is approximately four to six times more effective than transfer of T cells at preventing colonization of mice following exposure to Brucella (R) sp. Recent observations are in agreement with those data which suggested that the Omps may be a good candidate for inducing cellular immunity to infection by S strains and humoral immunity to R strains of Brucella (Cassatro et al., 2005).
Bowden et al. (1995 and 2000) and Estein et al. (2003) suggested that the major Omps group 2 are not very relevant as antigens in infections with B. abortus or B. melitensis, i.e., low or no protective activity in the mouse model of infection and low or no immunogenicity during host infection by using a recombinant protein technology and Monoclonal Antibodies (MAbs). However, group 3 Omp proteins, appear as immunodominant antigen during the course of infection with B. ovis, B. abortus, B. melitensis and as important protective antigen in the B. ovis, B. abortus, B. melitensis mouse model of infection. Furthermore, animals immunized with hot saline extract of B. ovis containing group 3 Ag were highly protected against infection with Brucella sp. (Jimenz et al., 1994). Therefore, Omps group 3 may not be very useful for full immunization against (S) species of Brucella through Ab stimulation (Riezu-Boj et al., 1990), they may induce a protective humoral immune response against infection with R species (Jimenez et al., 1994). Numerous reports are in agreement with this conclusion; Goldbaum et al. (1993) and Ciocchini et al. (2004) described that B. abortus Omp25 protein antigen may be used as a marker of active infection in humans, bovines and sheep with brucellosis. However, Omp25 is also one of the two major Omps in B. melitensis strains with strong immunoreactive properties (Vizcaino et al., 2001; Patrick et al., 2005). Omp2b in B. abortus and B. melitensis strains are highly immunogenic activity (Vizcaino et al., 2001). This finding was not surprising where also Omp26 stimulates humoral immune system in infected animals or humans with either B. abortus or B. ovis, respectively (Riezu-Boj et al., 1990). In this context, Stevens et al. (1995) found that 25 to 29 kDa proteins, are immunodominant in S2308-infected cattle. Characterization, expression and define the epitopes of Omp28 (Lindler et al., 1996) in Brucella sp. and a new variant of B. melitensis that infected human in Argentine (Lucero et al., 2006) may be of useful in vaccine development as well as diagnosis of the disease. Other studies by Vizcaino et al. (2001) for determination the reactivity of sera from B. ovis naturally infected animals against recombinant B. ovis Omp31 by Western blot revealed that recombinant B. ovis Omp31 is a promising candidate for the diagnosis and for vaccination of B. ovis infection in rams. These results are also in accordance with previous observations where good reactivity of sera from infected rams was obtained with the Omp31 protein extracted from B. ovis (Vizcaino et al., 1996). Therefore, it is interesting candidate for both diagnosis of infections caused by B. ovis in rams and the development of a subcellular vaccine against ram epididymits caused by B. ovis. On the other hand, Cassataro et al. (2005) investigated the immune response and protection elicited by DNA vaccine coding for Omp31. Where mice immunized with it were significantly protected against B. ovis and B. melitensis infection. However, similar levels of protection induced by this method to those induced by control vaccine suggesting that using this vaccine preparation is efficiency. Zygmunt et al. (1994) reported that using a set of monoclonal antibodies against some antigens of the cell envelope fraction Omp29-Omp34 for diagnosis of B. melitensis infection in sheep was promising, although they did not characterize these antigens. Therefore, Omps group 3 Ags are very promising as a tool for diagnosis as well as vaccination.
Recorded results demonstrated that infected human with either B. abortus or B. melitensis induced a humoral immune response directed against Bp23 protein Ag but not recognized in sera from negative human infection. In addition, Bp23 is present in the B. melitensis and B. abortus suggesting that this antigen might be specific for Brucellosis spp. (Fig. 5 lanes 2-7). Therefore, Bp23 protein of B. abortus have been considered important in eliciting an immune response related to the protection of the host and also for diagnostic purposes. Accordingly, the OMPs 23, 25, 27, 28 and 31 might be an interesting candidate for the development of a subcellular vaccine against humans and camel brucellosis that caused by B. melitensis and B. abortus strains. Characterization of those protein still need to be tested in human sera of patients with acute or chronic Brucellosis and patients with positive history of brucellosis. Further characterization of their genes, gene products and expression is required before we will understand more fully how these proteins and loci contribute to pathogenesis. However, until now there are no report on a subunit vaccine that is able to confer protection against Brucella. Thus, the identification of the epitope recognized by the Ab form human serum and the availability of recombinant protein will allow determination of their potential protective activity against Brucella infection and their potential for development of subcellular vaccine without the drawbacks of the living attenuated vaccines now commonly used. Being a 23 kDa in B. abortus and B. melitensis, it may have a particular utility for vaccination against human brucellosis and may be a vaccine candidate antigen that could be included in the development of a multisubunit vaccine in the immunoprophylaxis of brucellosis.
Differentiation between infected and vaccinated animals: The development of subcellular protein-based vaccines to prevent brucellosis has several advantages. One of the major advantages is the possibility to differentiate vaccinated animals from diseased animals. However, the current live attenuated vaccines against animal brucellosis induce high-titer Abs against the O- polysaccharide of the LPS which interferes with the serological diagnosis of the disease (Lindler et al., 1996). In contrast large-scale production of recombinant purified protein provides large quantities of Ag and devoid from all other contaminating Brucella macromolecules. In addition, Bowden et al. (1995) reported that, any protein component of the Brucella cell which elicits an Ab response that cross-reacts among all or most members of the genus Brucella would improve the specificity of diagnostic reagents for brucellosis. From this pilot experiment with vaccinated and infected animals, preliminary results seem to indicate that Bp23 antigen is located in both B. melitensis and B. abortus strains and serology response toward it may be very week in vaccinated and protected animals with either S19 or Rev1 strains, whereas it is strongly present in animals naturally infected with either B. abortus or B. melitensis as represented in western blot (Fig. 6). This could be explained in fact that host humoral responses appears different between the vaccinated and infected animal with Brucella strains where it is mounted in B. abortus and melitensis-infected camels compare to vaccinated camels. These data are in agreement with the immunoblotting analysis of a cell envelope fraction with infected and vaccinated sheep sera reported by Zygmunt et al. (1994). Moreover, antigenicity of sera collected from B. abortus and B. melitensis-infected camel flocks were clearly discriminated from sera of Brucella free animals towards Bp23 antigen (Fig. 6 lane 1 and 4). In addition, BP23 antigen was expressed in the pellet lysate buffer of the E. coli in large quantities by plasmid vectors, could be detected by in vivo Abs formed in animals and human serum of naturally infected B. abortus or B. melitensis strains (Fig. 5 and 6) and in vivo 23 kDa Ag could be detected in different animals infected with B. abortus strain by anti-Bp23 hyperimmune rabbit sera (Fig. 7) render this protein a good candidate for the development of a reagent to be used for differentiation diagnostic purposes.
Diagnosis of B. abortus in different domestic hosts: To evaluate the significance of anti- 23 kDa antigen for diagnostic purposes of putative Bp23 protein in sera, Western blot experiments were performed with total sera protein from camels, cows and sheep naturally infected in the field with B. abortus and were positive by classical serology methods. A band migrating at about 23 kDa was specifically recognized in 12 serum samples out of 21 infected animals (57%), among them 5 of 5 serum samples from camels (100%) and 4 of 7 cows (54%) and 3 of 9 serum samples from sheep (33%). The specificity of the Bp23 was assessed with sera from healthy animals that showing negative in all assays and no cross-reaction could be demonstrated which reflecting that this antigen might be specific for Brucella sp. (Fig. 7, panel D). It is clear that the sensitivity obtained with this protein seems no sufficient to allow detection of all infected animal tested here specially for cows (46%) and for sheep (67%) but can distinguished all infected camels. Therefore, the specificity obtained with this new antigen described here seems acceptable, although the sensitivity is not sufficient to allow detection of all infected animals. Difference in antigenicity detected between camels, cows and sheep might be explained if the Bp23 display some minor difference in amino acid sequences and consequently in epitope sequence of putative Bp23 in different animals as recorded in different antigens previously studied (Vizcaino et al., 2001), although the six species of the genus Brucella display a high degree of DNA homology (Vizcain et al., 2001). Also Western blot results demonstrate that rabbit infected with B. abortus recombinant Bp23 protein can elicit an Ab with high specificity which was mainly host specific since some infected sheep and cows do not appears to produced a humoral immune response directed against this Ag. On the other hand, sera from B. abortus infected camels and Ab specific for the B. abortus Bp23 proteins showed good reactivity as well as with the recombinant B. abortus Bp23 protein. Therefore complementation with other antigens will be necessary to enhance the overall sensitivity. Some other bands with a molecular mass not corresponding to Brucella Bp23 were detected in some strips (Fig. 7 panel C; lanes 6). It may be degraded Ag or presumably corresponded to E. coli proteins reacting with serum antibodies developed in response to common exposure of animals to this bacterium. As a result, Bp23 is the best antigen as a diagnostic material for diagnosis of B. abortus infected camels then cows and sheep, but further analysis should be performed with a large number of sera to evaluate its real usefulness.
The availability of the gene encoding this 23 kDa antigen now allows for large-scale production to be used for further analyze and to use this protein in different Brucella serology tests. Further characterization of Bp23 genes, gene products and expression is required for well understand how these proteins and loci contribute to pathogenesis. Studies are currently under way to characterize the expression of 23 k Da in Brucella sp. and define the epitopes on the protein which may be of use in vaccine development as well as diagnosis of the disease.
ACKNOWLEDGMENTS
Author acknowledge Dr. Zaki Youssef (KFH) for helping in preparation of λgt11 libraries, Lab. Group in KFH for the advising in preparation of the Abs and Dr. Zakaria H. for the preparation of the B. abortus lysate. The expert technical assistance from Dr. Mohamed Shohayeb was highly appreciated. Also author thanks Dr. Salh Bazaid for editorial comments. This study was financed in part by an EEC fellowship, Human Brucellosis (fellowship ITC-1426) Makkak Saudi Arabia.
REFERENCES
- Abbas, B. and H. Agab, 2002. A review of camel brucellosis. Prev. Vet. Med., 55: 47-56.
CrossRefDirect Link - Aguirre, S., A.M. Silber, M.E. Brito, M.E. Ribone, C.M. Lagier and I.S. Marcipar, 2006. Design, construction and evaluation of a specific chimeric antigen to diagnose chagasic infection. J. Clin. Microbiol., 44: 3768-3774.
Direct Link - Alballa, S.R., 1996. Epidemiology of human brucellosis in Southern Saudi Arabia. J. Trop. Med. Hyg., 98: 185-189.
Direct Link - Al-Eissa, Y.A., 1999. Brucellosis in Saudi Arabia: Past, present and future. Annu. Saudi Med., 19: 403-405.
Direct Link - Almuneef, M.A., Z.A. Memish, H.H. Balkhy, B. Alotaibi, S. Algoda, M. Abbas and S. A1subaie, 2004. Importance of screening household members of acute brucellosis cases in endemic areas. Epidemiol. Infect., 132: 533-540.
Direct Link - Al-Tawfiq, J.A., 2006. Brucella epididymo-orchitis: A consideration in endemic area. Int. Braz J. Urol., 32: 313-315.
Direct Link - Alton, G.G., L.M. Jones, R.D. Angus and J.M. Verger, 1988. Techniques for the Brucellosis Laboratory. 1st Edn., Institute Nationale de le Rech, France, Paris, Pages: 174.
Direct Link - Altschul, S.F., W. Gish, W. Miller, E.W. Myers and D.J. Lipman, 1990. Basic local alignment search tool. J. Mol. Biol., 215: 403-410.
CrossRefPubMedDirect Link - Blasco, J.M. and R. Diaz, 1993. Brucella melitensis Rev-1 vaccine as a cause of human brucellosis. Lancent, 342: 805-805.
Direct Link - Bowden, R.A., A. Cloeckaert, M.S. Zygnumt and G. Dubray, 1995. Outermembrane protein-and rough lipopolysaccharide-specific monoclonal antibodies protect mice against Brucella ovis. J. Med. Microbiol., 43: 344-347.
Direct Link - Bowden, R.A., S.M. Estein, M.S. Zygnnunt, G. Dubray and A. Cloeckaert, 2000. Identification of protective outer membrane antigens of Brucella ovis by passive immunization of mice with monoclonal antibodies. Microbes Infect., 2: 481-488.
CrossRefDirect Link - Cassatro, J., A.V. Carlos, D. Silvia, S. Estein and L. Bruno et al., 2005. A DNA vaccine coding for the Brucella outer membrane protein 31 confers protection against B. melitensis and B. ovis infection by eliciting a specific cytotoxic response. Infect. Immunol., 73: 6537-6546.
PubMedDirect Link - Ciocchini, A.E., M.S. Roset and R.A. Ugalde, 2004. Membrane topology analysis of cyclic glucan synthase, a virulence determinant of Brucella abortus. J. Bacteriol., 186: 7205-7213.
Direct Link - Cloeckaert, A., N. Vizcaino, J.Y. Paquet, R.A. Bowden and P.H. Elzer, 2002. Major outer membrane proteins of Brucella sp.: Past, present and future. Vet. Microbiol., 90: 229-247.
CrossRefPubMedDirect Link - Corbel, M.J., 1997. Brucellosis: An overview. Emerg. Infect. Dis., 3: 213-221.
CrossRefPubMedDirect Link - De Wergifosse, P., P. Lintermans, J.N. Limet and A. Clocekaert, 1995. Cloning and nucteotide sequence of the gene coding for the major 25-kilodalton outer membrane protein of Brucella abortus. J. Bacteriol., 237: 1911-1914.
Direct Link - Estein, S.M., J. Cassataro, N. Vizcaino, M. Zygmunt, A. Cloeckaert and R.A. Bowden, 2003. The recombinant Omp31 from Brucella melitensis alone or associated with rough lipopolysaccharide induces protection against Brucella ovis infection in BALB/c mice. Microbes. Infect., 5: 85-93.
Direct Link - Fallatah, S.M., A.J. Oduloju, S.N. Al-Dusari and Y.M. Fakunle, 2005. Human brucellosis in Northern Saudi Arabia. Saudi. Med. J., 26: 1562-1566.
Direct Link - Goldbaum, F.A., J. Leoni, J.C. Wallach and C.A. Fossall, 1993. Characterization of an 18 kDa Brucella cytoplasmic protein which appears to be a serological marker of active infection of both human and bovine brucellosis. J. Clin. Microbiol., 31: 2141-2145.
Direct Link - Guilloteau, L.A., K. Laroucau, N. Vizcaino, I. Jacques and G. Dubray, 1999. Immunogenicity of recombinant Escherichia coli expressing the Omp31 gene of Brucella melitensis in BALB/e mice. Vaccine, 23: 353-361.
Direct Link - Halling, S.M., B.D. Peterson-Burch, B.I. Bricker, R.L. Zuerner and Z. Qing et al., 2005. Completion of the genome sequence of Brucella abortus and comparison to the highly similar genomes of Brucella melitensis and Brucella suis. J. Bacteriol., 187: 2715-2726.
Direct Link - Hemmen, F., V. Weynants, T. Scarcez, J. Letesson and E. Saman, 1995. Cloning and sequencing analysis of a newly identified Brucella abortus gene are serological evaluation of the 17 kilodalton antigen that it encodes. Clin. Diagn. Lab. Immunol., 2: 263-267.
Direct Link - Huang, V., L.A. Babiuk and S. van Drunen, 2005. Immunization with a bovine herpesvirus 1 glycoprotem B DNA vaccine induces cytotoxic T-lymphocyte responses in mice and cattle. J. Gen. Virol., 86: 887-898.
Direct Link - Jimenez de Bagues, M.P., P.H. Eizer, J.M. Blasco, C.M. Marin, C. Gamazo and X.J. Winter, 1994. Protective immunity to Brucella ovis in BALB/e mice following recovery from primary infection or immunization with subcellular vaccines. Infect. Immun., 62: 632-638.
Direct Link - Kaufmann, A.F., M.I. Meltzer and G.P. Schmid, 1997. The economic impact of a bioterrorist attack: Are prevention and post-attack intervention programs justifiable? Emerg. Infect. Dis., 3: 83-94.
Direct Link - Kohler, S., S. Michaux-Charachon, F. Porte, M. Ramuz and J.P. Liautard, 2003. What is the nature of the replicative niche of a stealthy bug named Brucella? Trends Microbiol., 11: 215-219.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Larsson, P., P.C. Oyston, P. Chain, M.C. Chia and M. Duffield et al., 2005. The complete genome sequence of Francisella tularensis, the causative agent of tularemia. Nut. Genet., 37: 153-159.
Direct Link - Lindler, L.E., T.L. Hadfield, B.D. Tall, N.J. Snellings and F.A. Rubi
et al., 1996. Cloning of a Brucella melitensis group 3 antigen gene encoding Omp28, a protein recognized by the humoral immune response during human Brucellosis. Infect. Immun., 64: 2490-2499.
Direct Link - Lucero, N.E., S.M. Ayala, G.I. Escobar, M. Grayon and I. Jacques, 2006. A new variant of Brucella melitensis. Clin. Microbiol. Infect., 12: 593-596.
Direct Link - Mullis, K.B. and F.A. Faloona, 1987. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol., 155: 335-350.
CrossRefPubMedDirect Link - Patrick, S.G., D.J. Comerci, M.E. Tolmasky, F.W. Larimer and S.A. Malfatti et al., 2005. Whole-genome analyses of speciation events in pathogenic Brucellae. Infect. Immun., 73: 8353-8361.
Direct Link - Pugh, G.W. Jr., L.B. Tabatabal, M. Phillips and T.J. McDonald, 1991. Establishment of dose-response relationships in BALB/e mice, using Brucella cell surface protein and lipopolysaccharide. Am. J. Vet. Res., 52: 261-268.
Direct Link - Radwan, A.I., S.I. Bekairi, A.A. Mukayel, A.M. Al-Bokmy, P.V. Prasad, F.N. Azar and F.N. Coloyan, 1995. Control of Brucella melitensis infection in a large camel herd in Saudi Arabia using antibiotherapy and vaccination with Rev. 1 vaccine. Revue Scientifique Technique, 14: 719-732.
Direct Link - Riezu-Boj, J.L., I. Moriyou, J.M. Blasco, C. Ganiazo and R. Diaz, 1990. Antibody response to Brucella ovis outer membrane proteins in ovine brucellosis. Infect. Immun., 58: 489-494.
Direct Link - Roop, R.M., B.H. Bellaire, M.W. Valderas and J.A. Cardelli, 2004. Adaptation of the Brucellae to their intracellular niche. Mol. Microbiol., 52: 621-630.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sanger, F., S. Nicklen and A.R. Coulson, 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A., 74: 5463-5467.
CrossRefPubMedDirect Link - Schurig, G.G., N. Sriranganathan and M.J. Corbel, 2002. Brucellosis vaccines: Past, present and future. Vet. Microbiol., 90: 479-496.
CrossRefDirect Link - Stevens, M.G., S.C. Olsen, G.W. Pugh and D. Brees, 1995. Comparison of immune responses and resistance to brucellosis in mice vaccinated with Brucella abortus 19 and R1351. Infect. Immun., 63: 264-270.
Direct Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Vizcaino, N., A. Cloeckaert, M. Aygniunt and G. Dubray, 1996. Cloning, nucleotide sequence and expression of the Brucella melitensis Omp31 gene coding for an immunogenic major outer membrane protein. Infect. Immunol., 64: 3744-3751.
PubMedDirect Link - Vizcaino, N., A. Cloeckaert, M.S. Zygmunt and L. Fernandez-Lago, 2001. Characterization of a Brucella species 25-kilobase DNA fragment deleted from Brucella abortus reveals a large gene cluster related to the synthesis of a polysaccharide. Infect. Immun., 69: 6738-6748.
Direct Link - Yagupsky, P., 1999. Detection of brucellosis in blood cultures. J. Clin. Microbiol. Biol., 37: 3437-3442.
Direct Link - Young, E.J., 1995. An overview of human brucellosis. Clin. Infect. Dis., 21: 283-289.
CrossRefPubMedDirect Link - Zygmunt, M.S., A. Cloeckaert and G. Dubray, 1994. Brucella melitensis cell envelop protein and lipopolysaccarides epitopes involved in humoral immune responses of naturally and experimentaly infected sheep. J. Clin. Microbiol., 32: 2414-2522.
Direct Link