
Mesut Sahin
Department of Biology, Molecular Biology Section, Faculty of Science and Art, Celal Bayar University, Muradiye Campus, 45047, Manisa, Turkey
Erdal Balcan
Department of Biology, Molecular Biology Section, Faculty of Science and Art, Celal Bayar University, Muradiye Campus, 45047, Manisa, Turkey
Journal of Biological Sciences
Year: 2006 | Volume: 6 | Issue: 2 | Page No.: 294-300







ABSTRACT
In the present study, programmed cell death of skeletal muscle tissues of the regressing tail of Euroasian green toad Bufo viridis (Amphibia: Anura) tadpole was investigated with Hematoxylin-eosin (H+E) and TUNEL methods in the three groups of different tail lengths. TUNEL stainings indicate that, during the tail regression, apoptotic skeletal muscle tissue constitutes fragmentations and the progression of cell death occurs from the tip and outer of the tail to its base. In addition, muscle cells apoptosis occurs first near the subepidermal fibrolast layer proposed that these cells may interfere the skeletal muscle cells apoptosis during the tail regression. When the of the glycosaminoglycan alterations of these different sizes of shortening tail was analyzed histochemically. After the Alcian Blue-Periodic Acid Schiff`s (AB-PAS) sequences at critical electrolyte concentrations and different pH values, we observed carboxylated glycosaminoglycans (i.e., hyaluronic acid, HA) are increased in apoptotic muscle cells throughout the tail regression. We concluded that carboxylated glycosaminglycans may play an important role in shortening of the tail via muscle cell apoptosis.
PDF Abstract XML References Citation
How to cite this article
Mesut Sahin and Erdal Balcan, 2006. The Glycoconjugate Changes of Apoptotic Skeletal Muscle Tissues in Regressing Eurasian Green Toad, Bufo viridis (Amphibia: Anura) Tadpole Tail. Journal of Biological Sciences, 6: 294-300.
DOI: 10.3923/jbs.2006.294.300
URL: https://scialert.net/abstract/?doi=jbs.2006.294.300
DOI: 10.3923/jbs.2006.294.300
URL: https://scialert.net/abstract/?doi=jbs.2006.294.300
INTRODUCTION
During the amphibian metamorphosis, drastic physiological, morphological and biochemical changes occur. Tadpoles change from aquatic herbivores to carnivorous, land-dwelling frogs. The tail resorption is seen during this time (Pretty et al., 1995). Cell death without replacement in the tail is an important component of the metamorphic climax.
Apoptosis or programmed cell death is a regulatory process and is crucial for development and tissue homeostasis. This process is characterized by unique morphological and biochemical alterations, such as membrane blebbing, nuclear breakdown, chromosomal fragmentation and the bundling of cellular contents into vesicles called apoptotic bodies that are marked for phagocytosis, which make it distinct from necrosis, a form of cell death result from a serious physical or chemical damage (Kerr et al., 1972). Regardless of the mode of apoptotic initiation, the morphological characteristics of apoptosis are consistent in most cell types.
The decreasing of skeletal muscle mass with age, via a reduction in fiber number and atrophy of the remaining muscle fibers, is arranged by unidentified mechanisms (Holloszy et al., 1991; Lexell, 1995). While the proteolytic (Dean et al., 1997; Grune et al., 1997; Chevion et al., 2000), neurological and hormonal mechanismsPP (Roubenoff and Hughes, 2000) play an important role in skeletal muscle loss, apoptosis may also be important.
Complex carbohydrates are found all living organisms and play a major role in many biological processes, such as cell to cell and host-microbe interactions and signal transmission. With the finding of cell surface carbohydrate binding proteins, lectins (Drickamer, 1988), it was shown that carbohydrate chains of cell surface glycoproteins, glycolipids and proteoglycans have an important function as recognition molecules in many different cellular processes such as fertilization, development and pathogen-host interactions (Tang et al., 1985; Feizi, 1991; Brandley, 1991; 1992; Varki, 1997; Gabius, 2002; Kilpatrick, 2002; Sharon and Lis, 2004). Within the last decade, many reports regarding apoptosis-glycan relationships have been notified (Kepler et al., 1994; Suzuki et al., 2003; Kim et al., 2004; Murrell et al., 2004; Eda et al., 2004).
Glycosaminoglycans (GAGs), linear polymers of repeating disaccharides of hexosamine plus a uronic acid such as glucuronic acid, are found in the cell surface and extracellular matrix and responsible for the matrix organizations, cell migration, adhesion, differentiation and apoptosis (Davies et al., 2001).
In the present study, using histochemical techniques we compared the GAG contents in apoptotic skeletal muscle tissues of Euroasian green toad Bufo viridis tadpole tails which divided into three groups according to the 15, 10 and 5 mm tail lengths.
MATERIALS AND METHODS
Preparation of samples: Eurasian green toad Bufo viridis were collected on March 2003 and March 2004, as eggs from ponds located the neighbourhood of Celal Bayar University Campus Area and were allowed to hatch in plastic containers (50 cm diameter; 25 L volume). After hatching, larvae were fed ad libitum a diet of proteinated nutritions. All focal tadpoles of Bufo viridis used in the experiment were cold-anesthetized in +4 °C. Tails were cut with a razor blade and grouped into three lengths containing 15, 10 and 5 mm. Thereafter, tail grups were fixed with formalin, Bouin`s and Saint Marie fixatives, dehydrated and embedding to paraffin according to routine procedures and 5 μm sections were prepared.
Staining procedures
Apoptosis assay: In order to determine of apoptotic cells, TUNEL (terminal deoxyribonucleotidyl transferase-mediated dUTP-biotin nick end labeling) assays on tissue sections were performed using Promega's DeadEnd™ ColorimetricTUNEL System (cat # G7130) according to the manufacturer`s instructions. Briefly, following postfixation in 4% paraformaldehyde, permeabilisation with proteinase K treatment, 4% paraformaldehyde again and PBS washes, samples were incubated with biotynlated nucleotide and TdT enzyme at 37 °C for 30 min. Following washes and blocking endogen peroxidase activity using H2O2, the biotin labels was detected with streptavidin-HRP (horse radish peroxidase) and a DAB (diaminobenzidine tetrahydrochloride dihydrate) color reaction.
Alcian blue staining at critical electrolyte concentrations: We performed mucin histochemical studies on three different tail lengths using combined Alcian Blue (AB)-Periodic Acid Schiff (PAS) sequences as described previously (Mowry, 1963; Spicer et al., 1967; Yamabayashi, 1987). GAGs were identified by critical electrolyte concentrations at which the polyanions changed from binding AB to MgPP++ (Scott and Dorling, 1965). AB stained polyanions with increasing selectivity as the MgCl2 concentration in the staining solution increased: in the presence of MgCl2 concentrations below 0.2 M, nucleic acids, carboxylated GAGs (i.e., hyaluronic acid, HA) and sulfated GAGs, including heparan sulfate (HS) are stained; at 0.2 M MgCl2 and above, staining for HA is lost but staining for sulphated GAGs (i.e., chondroitin sulfate, CS, dermatan sulfate, DS, heparan sulfate, HS and keratan sulfate, KS) is retained (Table 1).
Briefly, sections were immersed in 0.05% Alcian Blue 8GX (Fluka, Russian Fed.) in 0.025M acetat buffer solution containing 0.025 and 0.3 M MgCl2 at a final pH 5.8 for 18 h to stain GAGs (the critical electrolyte concentration procedure, referred in 27) and counterstained with Periodic acid-Schiff`s reagent. In order to determine neutral and acidic glycoconjugates, sections treated with 1% Alcian Blue 8GX pH 2.5 for 30 min and PAS used as counterstain.
RESULTS
Apoptosis: During the tail regression skeletal muscle cells undergo apoptosis in case of fragmantations (Fig. 1 and 2). and the apoptotic process of muscle tissues begins from just below of the subepidermal fibroblast layer of tail and continues to inside (Fig. 3 and 4). In addition to skeletal muscles, epithelial cells of skin, notochord, extracellular matrix components are also be degraded during the tail regression (not shown).
Alcian blue stainings
15 mm length tails: With AB (pH 5.7)-PAS sequence at 0.025 M MgCl2, which stains carboxylated and sulphated GAGs and sulphated glycoproteins, the apoptotic and intact muscle tissues were weakly positive, while majority of the intact muscle tissues were PAS positive. At the 0.3 M MgCl2 concentration of AB (pH 5.7)-PAS, which stains sulphated GAGs, an AB positivity was not seen in apoptotic and non-apoptotic muscle tissues. These staining characteristics were same with those
| Table 1: | Staining specificities of AB-PAS sequences at different pH values and electrolyte concentrations |
 | |
| PAS: Periodic acid Schiff`s, AB: Alcian blue, M: Magenta, B: Blue, T: Turquoise, .GP: Glycoprotein, GAG: Glycosaminoglycan, GC: glycoconjugate | |
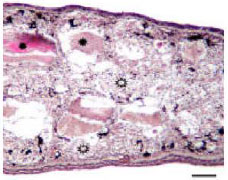 | |
| Fig. 1: | Apoptotic (star frame) and non-apoptotic (filled star) skeletal muscle tissues in regressing tadpole tail. H+E staining. Bar: 100 μ |
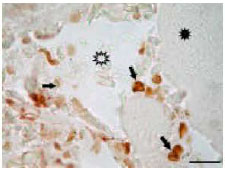 | |
| Fig. 2: | Apoptotic (star frame) and non-apoptotic (filled star) skeletal muscle tissues in regressing tadpole tail. Arrows indicate apoptotic bodies. TUNEL staining. Bar:20 μ |
 | |
| Fig. 3: | Apoptosis begins from outside and continues to inside during the tail regression. Star frame: apoptotic muscle tissues, filled star: non-apoptotic tissue, N: notochord, arrow: spinal cord. TUNEL + Hematoxylin Bar: 100 µ |
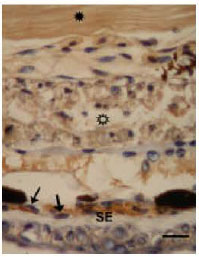 | |
| Fig. 4: | Apoptotic muscle cells (star frame) found just below subepidermal fibroblast layer (SE). Filled star: non-apoptotic muscle tissues, arrows: fibroblasts. TUNEL + Hematoxylin Bar: 10 |
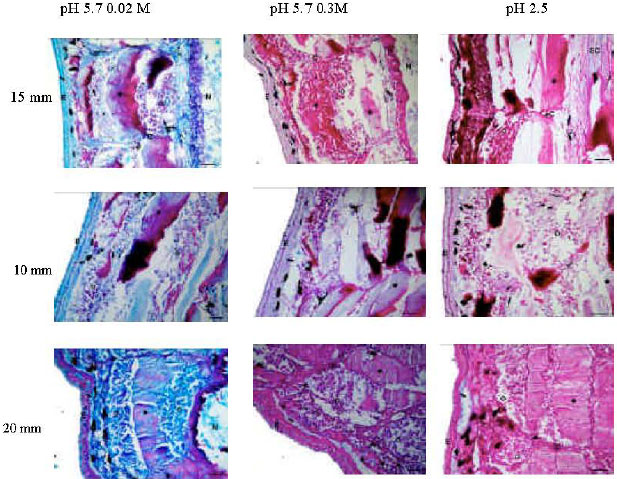 | |
| Fig. 5: | AB-PAS sequences of three different tail lengths at different pH values and electrolyte concentrations. E: epithelial cell layer, N: notochord C: cleft between two muscle tissues, star frames: apoptotic muscle tissues, filled stars: non-apoptotic muscle tissues, SC: spinal cord, arrows: subepidermal fibroblast layers Bars: 50 μ |
| Table 2: | Staining specificities of different parts of tail groups |
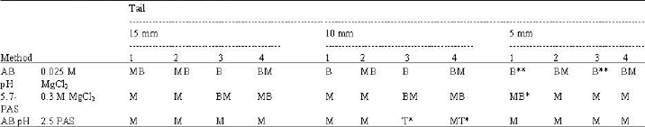 | |
| M: Magenta, B: Blue, T: Turquoise, *: Weak staining, **: Strong staining, 1: Apoptotic muscle cells, 2: Non-Apoptotic muscle cells, 3: Tip, 4: Skin | |
of AB pH 2.5-PAS, which specific for acid mucins (Fig. 5 and Table 2).
10 mm length tails: At the 0.025 M MgCl2 concentration of AB (pH 5.7)-PAS sequence, the almost of intact muscle tissues give PAS positive reaction. Alcianophilia of apoptotic muscle cells was increased when compared with 15 mm at this concentration. As in the 15 mm, skin and tip of the 10 mm tail positively stained with AB. Alcianophilia of apoptotic muscle cells was decreased at 0.3 M MgCl2 concentration of AB-PAS sequence, while PAS staining was increased in not only intact muscle cells, but in apoptotic ones. Meanwile, the AB staining of skin and the tip of tail were heavily decreased at this concentration. At the AB (pH 2.5)-PAS staining, while the intact muscle cells give strong PAS positivity, apoptotic cells stain weakly with PAS. In addition, alcianophilia of apoptotic muscle cells was more decreased (Fig. 5 and Table 2).
5 mm length tails: When compared with intact muscle tissue, alcianophilia of apoptotic muscle cells were more increased at 0.025 M MgCl2 AB (pH 5.7)-PAS sequence. Some portions of intact tissue were PAS positive. At the 0.3 M MgCl2 AB (pH 5.7)-PAS, alcianophilia of the intact and apoptotic tissues was very decreased, while PAS positivity was increased. The AB (pH 2.5)-PAS stainings were similar to the 0.3 M.
At the different electrolyte concentrations and pH, we indicated that carboxylated GAGs (i.e., HA) were main glycoconjugates of apoptotic and non apoptotic muscle tissues of the resorbing tail, because alcianophilia of these tissues at 0.025 M MgCl2, which specific for sulphated glycoproteins, carboxylated and sulphated GAGs, was decreased at 0.3 M concentration. Meanwhile when compared the alcianophilias of three different tail lengths at 0.025 M concentrations of AB (pH 5.7)-PAS sequences, we indicated that HA is increased in apoptotic muscle tissues during the tail regression (Fig. 5 and Table 2).
DISCUSSION
Apoptosis, or programmed cell death a physiological process which essential for embryonic development and homeostasis. Apoptotic mechanisms play a role in many tissue and organ development during the embryogenesis (Renehan et al., 2001).
Throughout the anuran metamorphosis, removal of the larval tissues presumably accomplished by cell death (Shi et al., 2001). Kerr et al. (1974), who are the first to demonstrate that apoptosis occurs during anuran metamorphosis and others (Watanabe and Sasaki, 1974; Nishikawa and Hayashi, 1995) found that the epidermal and muscle cells of shortening tail undergo a series of morphological changes of apoptosis, including the condensation of the cytoplasm and the nuclear chromatin and the subsequent formation of the apoptotic bodies.
In the sagittal and longitudinal sections of European green todd, Bufo viridis tadpoles were analysed in this study, TUNEL positive cells between the intact muscle tissues indicated that, the muscle tissues undergo apoptosis as fragmetations throughout the tail resorbtion (Fig. 2 and 3). In addition to muscle tissues, extracellular matrices, spinal cord, notocorda and blood vessels are also breakdown (not shown). Therefore, body decreasing of tail occurs through from outside to inside and tail have a jelly-like structure and epithelial layer of tail becomes multilayered (Fig. 1 and 3)
It has been suggested that, this reorganization may result increasing with development of thyroid hormone (Zhou and Brown, 1993; Brown et al., 1996; Nakajima et al., 2005). Sachs et al. (2000) shown that thyroid receptors can promote both cell proliferation and apoptosis during metamorphosis, depending upon the cell type in which they are expressed.
Nowadays two models are proposed for the cell death in reducing tadpole tail: first, suicide model, suggest that cells directly respond the hormon increases and the second, murder model, where cells killed with another cells or extracellular matrix (Nakajima et al., 2005). The murder model supported with “anoikis”? which is a programmed cell death occurring with breakdown of normal epithelial cell-extracellular matrix interactions (Meredith et al., 1993; Boudreau et al., 1995; Frisch and Screaton, 2001).
When the sagittal and longitudinal sections were analyzed, body decreasing of tail occurs through from outside to inside which starting from terminal side of tail in consequence of muscle cell death and breakdown of extracellular matrices, spinal cord, notocorda and blood vessels. Thus, tail transforms to a jelly-like structure and epithelial cell layer of tail becomes multilayered, probably by thyroid hormone (Das et al., 2002; Nakajima and Yaoita, 2003; Nakajima et al., 2005). On the other hand, it was suggest that matrix metalloproteinases appear to regulate not only extracellular matrix degradation but also programmed cell death, cell migration and invasion during the morphogenic processes (Werb and Chin, 1998).
It was shown that, the muscle cell apoptosis in the tail is at least in part facilitated by remodeling of the surrounding extracellular matrix directly or indirectly mediated by some matrix metalloproteinases (Damjanovski et al., 1999) which are induced in subepidermal fibroblast layer (Nakajima et al., 2005). Interestingly muscle cell, apoptosis occurs first near the subepidermal fibroblast layer suggest that, as a result of murder model, these cells may play a role in muscle cell apoptosis (Fig. 4) via releasing of some extracellular matix degrading proteinases which detaches the muscle cells from extracellular matrix (Nakajima et al., 2005). These results suggest that both of two death models in muscle cells can be seen in regression of tadpole tail during the development of Bufo viridis.
While the larval myoblast replace with adult myotubes in the trunk, in the tail, they undergoes complete regression during metamorphosis (Shimizu-Nishikawa et al., 2002). The earliest studies showed that the primary GAGs synthesized from both myoblasts and newly fused myotubes are hyaluronic acid and chondroitin sulfate (Ahrens et al., 1977; Angello and Hauschka, 1979). On the other hand, during the transformation of myoblasts to myotubes in the cell lines a proportional increase in the synthesis of heparan sulfate and proportional decreases in the chondroitin sulfate and hyaluronic acid synthesis was suggested (Pacifici and Molinaro, 1980). However, we found that the carboxylated GAG (i.e., HA) levels of apoptotic muscle tissues are increased throughout the tail regression.
The knowledge about the GAGs and their roles in the resorbing frog tail are not sufficient at present. In the resorbing tail, while the carboxylated GAG levels are increasing, sulphated GAGs are decreased (Fig. 5 and Table 2). Moreover, the decreasing of AB pH 2.5 stainings in shortening tail thought that acid mucin levels can diminish during the tail regression.
The different AB (pH 5.7)-PAS staining patterns at critical electrolyte concentrations in apoptotic muscle cells show that carboxylated GAGs (i.e., HA) are found heavily in these region when compared to the intact part of muscle tissue (Fig. 5 and Table 2). The possible reason of the increasing of carboxylated GAG levels of this region is these molecules have a crucial role in muscle apoptosis.
As a conclusion, the programmed cell death occurred in tadpole tail is a different process which may variable in stage by stage and may contain different death types involving the extracellular matrix and GAGs.
REFERENCES
- Ahrens, P.B., M. Solursh and S. Meier, 1977. The synthesis and localization of glycosaminoglycans in striated muscle differentiating in cell culture. J. Exp. Zool., 202: 375-388.
PubMedDirect Link - Boudreau, N., C.J. Sympson, Z. Werb and M.J. Bissel, 1995. Supression of ICE and apoptosis in mammary epithelial cells by extracellular matrix. Science, 267: 891-893.
PubMedDirect Link - Brandley, B.K., N. Rao, M. Nashed, F. Dasgupta, S. Abbas, D. Asa, T. Gant and J. Musser, 1992. Carbohydrates and their Analogs as Selectin Ligands and Antagonists. In: Leukocyte Adhesion. Basic and Clinical Aspects, Gahmberg, C.G., T. Mandrup-Poulsen, L.B. Wogensen and B. H�kfelt (Eds.). Elsevier Science Publisher, B.V., pp: 99-111.
- Brown, D.D., Z. Wang, J.D. Furlow, A. Kanamori, R.A. Schwartzman, B.F. Remo and A. Pinder, 1996. The thyroid hormone-induced tail resorption program during Xenopus leavis metamorphosis. Proc. Natl. Acad. Sci. USA., 93: 1924-1929.
PubMedDirect Link - Chevion, M., E. Berenshtein and E.R. Stadtman, 2000. Human studies related to protein oxidation: Protein carbonyl content as a marker of damage. Free Radic. Res., 33: S99-S108.
Direct Link - Damjanovski, S., A. Ishizuya-Oka and Y.B. Shi, 1999. Spatial and temporal regulation of collagenases-3,-4 and stromelysin-3 implicates distinct functions in apoptosis and tissue remodeling during frog metamorphosis. Cell Res., 9: 91-105.
CrossRefDirect Link - Das, B., A.M. Schreiber, H. Huang and D.D. Brown, 2002. Multiple thyroid hormone-induced muscle growth and death programs during metamorphosis in Xenopus laevis. Proc. Natl. Acad. Sci. USA., 99: 12230-12235.
Direct Link - Davies, J.A., C.E. Fisher and M.W. Barnett, 2001. Glycosaminoglycans in the study of mammalian organ development. Biochem. Soc. Trans., 29: 166-171.
PubMedDirect Link - Dean, R.T., S. Fu, R. Stocker and M.J. Davies, 1997. Biochemistry and pathology of radical-mediated protein oxidation. Biochem. J., 324: 1-18.
CrossRefPubMedDirect Link - Drickamer, K., 1988. Two distinct classes of carbohydrate-recognition domains in animal lectins. J. Biol. Chem., 263: 9557-9560.
PubMedDirect Link - Eda, S., M. Yamanaka and M. Beppu, 2004. Carbohydrate-mediated phagocytic recognition of early apoptotic cells undergoing transient capping of CD43 glycoprotein. J. Biol. Chem., 279: 5967-5974.
PubMedDirect Link - Feizi, T., 1991. Carbohydrate differentiation antigens: Probable ligands for cell adhesion molecules. Trends Biochem. Sci., 16: 84-86.
PubMedDirect Link - Frisch, S.M. and R.A. Screaton, 2001. Anoikis mechanisms. Curr. Opin. Cell Biol., 13: 555-562.
PubMedDirect Link - Gabius, H.J., 2002. Animal lectins and life: A guided tour into the realm of sugar code. Biochim. Biophys. Acta, 1572: 163-164.
CrossRefDirect Link - Grune, T., T. Reinheckel and K.J. Davies, 1997. Degradation of oxidized proteins in mammalian cells. FASEB J., 11: 526-534.
Direct Link - Holloszy, J.O., M. Chen, G.D. Cartee and J.C. Young, 1991. Skeletal muscle athrophy in old rats: Differential changes in three fiber types. Mech. Ageing Dev., 60: 199-213.
PubMedDirect Link - Keppler, O.T., M.E. Peter, S. Hinderlich, G. Moldenhauer and P. Stehling et al., 1999. Differential sialylation of cell surface glycoconjugates in a human B lymphoma cell line regulates susceptibility for CD95 (APO-1/Fas)-mediated apoptosis and for infection by a lymphotropic virus. Glycobiology, 9: 557-569.
Direct Link - Kerr, J.F., A.H. Wyllie and A.R. Currie, 1972. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer, 26: 239-257.
PubMedDirect Link - Kerr, J.F., B. Harmon and J. Searle, 1974. An electron-microscope study of cell deletion in the anuran tadpole tail during spontaneous metamorphosis with special reference to apoptosis of striated muscle fibers. J. Cell Sci., 14: 571-585.
Direct Link - Kilpatrick, D.C., 2002. Animal lectins: A historical introduction and overview. Biochim. Biophys. Acta, 1572: 187-197.
CrossRefDirect Link - Kim, E.J., S.G. Sampathkumar, M.B. Jones, J.K. Rhee, G. Baskaran, S. Goon and K.J. Yarema, 2004. Characterization of the metabolic flux and apoptotic effects of O-hydroxyl-and N-acyl-modified N-acetylmannosamine analogs in Jurkat cells. J. Biol. Chem., 30: 18342-18352.
Direct Link - Lexell, J., 1995. Human aging, muscle mass and fiber type composition. J. Gerontol. Biol. Sci. Med. Sci., 50: 11-16.
PubMedDirect Link - Meredith, J.E.J., B. Fazeli and M.A. Schwartz, 1993. The extracellular matrix as a cell survival factor. Mol. Biol. Cell, 4: 953-961.
Direct Link - Mowry, R.W., 1963. The special value of methods that color both acidic and vicinal hydroxyl groups in the histochemical study of mucins. With revised directions for the colloidal iron stain, the use of alcian blue 8GX and their combination with the periodic-acid-Schiff reaction. Ann. N.Y. Acad. Sci., 106: 402-423.
- Murrell, M.P., K.J. Yarema and A. Levchenko, 2004. The system biology of glycosylation. Chembiochem, 5: 1334-1347.
CrossRefDirect Link - Nakajima, K. and Y. Yaoita, 2003. Dual mechanisms governing muscle cell death in tadpole tail during amphibian metamorphosis. Dev. Dyn., 227: 246-255.
PubMedDirect Link - Nakajima, K., K. Fujimoto and Y. Yaoita, 2005. Programmed cell death during amphibian metamorphosis. Semin. Cell Dev. Biol., 16: 271-280.
CrossRefDirect Link - Nishikawa, A. and H. Hayashi, 1995. Spatial, temporal and hormonal regulation of programmed muscle cell death during metamorphosis of the frog Xenopus laevis. Differentiation, 59: 207-214.
Direct Link - Pacifici, M. and M. Molinaro, 1980. Developmental changes in glycosaminoglycans during skeletal muscle cell differentiation in culture. Exp. Cell Res., 126: 143-152.
PubMedDirect Link - Pretty, R., T. Naitoh and R.J. Wassersug, 1995. Metamorphic shortening of the alimentary tract in anuran larvae (Rana catesbeiana). Anat. Rec., 242: 417-423.
PubMedDirect Link - Renehan, A.G., C. Booth and C.S. Potten, 2001. What is apoptosis and why is it important?. BMJ, 322: 1536-1538.
Direct Link - Roubenoff, R. and V.A. Hughes, 2000. Sarcopenia: Current concepts. J. Gerontol. A Biol. Sci. Med. Sci., 55: M716-M724.
Direct Link - Sachs, L.M., S. Damjanovski, P.L. Jones, Q. Li and T. Amano et al., 2000. Dual functions of thyroid hormone receptors during Xenopus development. Comp. Biochem. Physiol. B Biochem. Mol. Biol., 126: 199-211.
PubMed - Scott, J.E. and J. Dorling, 1965. Differential staining of acid glycosaminoglycans (Mucopolysaccharides) by alcian blue in salt solutions. Histochemie, 5: 221-233.
CrossRefDirect Link - Sharon, N. and H. Lis, 2004. History of lectins from hemagglutinins to biological recognition molecules. Glycobiology, 14: R53-R62.
CrossRefDirect Link - Shi, Y.B., L. Fu, S.C. Hsia, A. Tomita and D. Buchholz, 2001. Thyroid hormone regulation of apoptotic tissue remodeling during anuran metamorphosis. Cell. Res., 11: 245-252.
CrossRefDirect Link - Shimizu-Nishikawa, K., Y. Shibota, A. Takei, M. Kuroda and A. Nishikawa, 2002. Regulation of specific developmental fates of larval-and adult-type muscles during metamorphosis of the frog Xenopus. Dev. Biol., 251: 91-104.
CrossRefDirect Link - Suzuki, O., Y. Nozawa and M. Abe, 2003. Sialic acids linked to glycoconjugates of Fas regulate the caspase-9-dependent and mitochondria-mediated pathway of Fas-induced apoptosis in Jurkat T cell lymphoma. Int. J. Oncol., 23: 769-774.
Direct Link - Tang, P.W., H.C. Gool, M. Hardy, Y.C. Lee and T. Feizi, 1985. Novel approach to the study of the antigenicities and receptor functions of carbohydrate chains of glycoproteins. Biochem. Biophys. Res. Commun., 132: 474-480.
PubMedDirect Link - Yamabayashi, S., 1987. Periodic acid-Schiff-alcian blue: A method for the differential staining of glycoproteins. Histochem. J., 19: 565-571.
CrossRefDirect Link - Varki, A., 1997. Sialic acids as ligands in recognition phenomena. FASEB J., 11: 248-255.
PubMedDirect Link - Watanabe, K. and F. Sasaki, 1974. Ultrastructural changes in the tail muscles of anuran tadpoles during metamorphosis. Cell Tissue Res., 155: 321-336.
CrossRefDirect Link - Werb, Z. and J.R. Chin, 1998. Extracellular matrix remodeling during morphogenesis. Ann. N. Y. Acad. Sci., 857: 110-118.
PubMedDirect Link - Zhou, W. and D.D. Brown, 1993. Thyroid hormone-induced gene expression program for amphibian tail resorption. J. Biol. Chem., 268: 16270-16278.
Direct Link