
Muhammad Subhan
Faculty of Agriculture, Gomal University, Dera Ismail Khan, Pakistan
Himayat Ullah Khan
Faculty of Agriculture, Gomal University, Dera Ismail Khan, Pakistan
Riaz-ud-Din Ahmad
Faculty of Agriculture, Gomal University, Dera Ismail Khan, Pakistan
Journal of Biological Sciences
Year: 2001 | Volume: 1 | Issue: 3 | Page No.: 120-123







ABSTRACT
Significant differences among hybrids and their parents were noted in number of bolls per plant, seeds per boll, and lint percentage (GOT), exposed additive type of gene influence while Vr/Wr graphs indicated over-dominance type of gene action for seed cotton yield per plant. The non-significant deviation of regression line from unit slope indicates the absence of non-allelic interaction in number of bolls per plant, yield of seed cotton, and seeds per boll showing absence of epistasis which provides positive out lines for isolating superior genotypes. The significant deviation from unit slope indicates the presence of epistasis in lint percentage (GOT) which confirmes its less effectiveness from selection point of view.
PDF Abstract XML References Citation
How to cite this article
Muhammad Subhan, Himayat Ullah Khan and Riaz-ud-Din Ahmad, 2001. Population Analysis of Some Agronomic and Technological Characteristics of Upland Cotton (Gossypium hirsutum L.), Using Diallel Analysis. Journal of Biological Sciences, 1: 120-123.
DOI: 10.3923/jbs.2001.120.123
URL: https://scialert.net/abstract/?doi=jbs.2001.120.123
DOI: 10.3923/jbs.2001.120.123
URL: https://scialert.net/abstract/?doi=jbs.2001.120.123
INTRODUCTION
Cotton being an important fibre crop also termed a silver fibre, occupies a key position as foreign exchange earning source. It is, therefore, necessary to have knowledge of the heredity mechanism of various characters in cotton. Research work on genetic improvement of cotton have been undertaken by many breeders and as a result,large number of improved varieties have been evolved. Amin et al. (1989) noted that the chracters like number of bolls per plant and yield of seed cotton are found to be goverened by over-dominance type of gene action. Rehman et al. (1988) reported in his study that the number of bolls per plant is controlled by genes which are additive in nature. The involvement of non-allelic interaction was observed in the inheritance of number of bolls per plant and yield of seed cotton. Sayal et al. (1995) confirmed that the number of bolls and numbrer of seeds per boll are controlled by additive type of gene action.
MATERIALS AND METHODS
Eight varieties of cotton Gossypium hirsutum L, viz; AUH-50, CIM-70, NIAB-78, CIM-240, CYTO-21/91, CYTO-14/87, MNH-410 and MNH-156 and their F3-seed of all possible combinations obtained from the Department of Plant Breeding and Genetics, Faculty of Agriculture, Gomal University, Dera Ismail Khan.
The above stated F3 material along with their parents were swon in the crop season 1998. The experiment was arranged in Randomized Complete Block Design with three replications. The experimental plot was kept as 48 x 3.60 m with 30 cm plant to plant distance. While row to row distance was set as 75 cm. Each treatment comprised of single 3.60 m long row. The number of plants per genotype were 12.
Statistical analysis: The data regarding different plant characters were subjected to statistical manipulation for the analysis of variance technique to establish the level of variability among F3 hybrids and their parental lines (Steel and Torrie, 1980). Further analysis for the diallel cross technique as applied by Whitehouse et al. (1958) was adopted for genetic analysis. All the crosses were arranged into arrays in the form of diallel tables and two statistics, the variance (Vr) of the family mean within an array and the covariance (Wr) of these means with the non-recurrent parental values were calculated from each diallel table by the following methods.
Estimation of Variances and Covariances:
Variance was calculated by the following formula:
![]()
Covariance (Wr) was calculated by the following formula:
![]()
Calculation of Limiting Parabola:The information on gene action was inferred by plotting the covariance (Wr) of each array against its variance (Vr). The slope and the position of the regression line fitted to the array points within the limiting parabola (Wr2= Vp--.Vr) indicated the degree of dominance and the presence or absence of gene interaction.The limiting parabola was constructed on the basis of formula, Wr2=VrxVp-- by plotting Vr, (VrxVp--)1/2 points. The corresponding values of Wr for all observed Vr values were calculated as (VrxVp--)1/2; where Vp-- = variance of the parents. The different arrays (varieties) were fitted within the limits of the parabola using the individual variance and covariance as their limiting point. Array nearest to the point of origin possessed most of the dominant genes, while the array that lay the farthest from the origin possessed the most recessive genes and the intermediate position signified the presence of both dominant and recessive genes in the array. The standard error for the regression line slope was estimated.
RESULTS AND DISCUSSION
Number of Bolls Per Plant: Observation of Table 1, reveals that the regression line intercepts the covariance axis above the origin, hence signifies an additive type of gene action with partial dominance for number of bolls per Plant. From the position of array points along the regression line, it is evident that cyto 21/91 possesses the most dominant genes for No. of bolls per plant because of its nearest position to the lower interception of line and parabola while NIAB-78 receives most recessive genes for No. of bolls per plant being away from origin. The regression line does not deviate significantly from the unit slope thus indicating the absence of non-allelic interaction.
| Table 1: | 8 X 8 Diallel table for average number of bolls per plant |
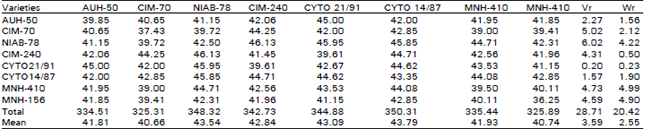 | |
| Table 2: | 8 x 8 Diallel table for average number of seeds per boll |
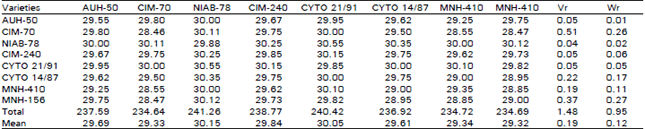 | |
| Table 3: | 8 x 8 Diallel table for lint percentage per plant |
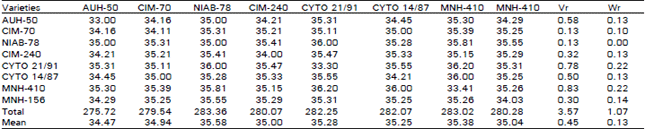 | |
| Table 4: | 8 x 8 Dallel table for seed cotton yield per plant |
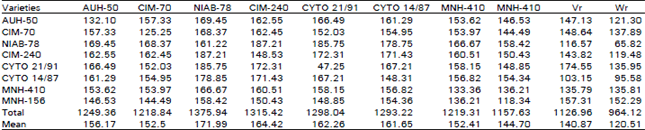 | |
Number of bolls per plant is a quantitative character and is one of the important components of yield of seed cotton. Indeed the alleles which gave the highest number of bolls tend to be dominant, additive type of genetic variance is responsible for this character. These observations are quite in line with those already reported by Mirza and Khan (1984) and Rehman et al. (1988) who reported to be additively controlled. While according to Kassem et al. (1986), Khan and Ghafoor (1986) and Ghafoor and Khan (1987), this character is non-additively controlled and thus did not agree with the present observations. Their differences might largely be attributed to different germplasm experimental materials with a varied genetic background in this respect.
Seed Cotton: An examination of Table 3, show that the regression line cuts the covariance axis below the origin, hence reflecting for overdominance type of gene action. From the position of array points on the regression line it becomes apparent that NIAB-78 being nearest to the origin keeps all the dominant genes for yield of seed cotton per plant. While CYTO-21/91 has the most recessive genes for yield of seed cotton per plant because of being away from the origin. The regression line does not deviate significantly from unit slope,indicating thereby the absence of non allelic interaction. The genetic mechanism of the control of yield of seed cotton is of overdominance type and in a diallel analysis this type of situation is depicted when progenies score more than either of the parents, which in other words means the manifestation of heterotic effect in this respect. Khan (1984), Mirza and Khan (1984) and Sayal et al. (1995) reported additive type of genetic control in respect of yield of seed cotton per plant. While Kassem et al. (1986), Khan and Ghafoor (1986) and Ghafoor and Khan (1987) reported overdominance type of gene action for yield of seed cotton per plant. The deviation with respect of phenotypic manifestation of the parameter like yield of seed cotton per plant might be due to different varieties used under different environmental conditions.
Number of Seeds Per Boll: A reference to Table 2 and 4 indicate that the regression line intercepts the covariance axis slightly above the origin and hence signifies the additive type of gene action with partial dominance. It is clear from the position of array points along the regression line that NIAB-78 being nearest to the origin contains the most dominant genes for number of seeds per boll. While CIM-70,being lying farthest from the origin having the most recessive genes for the character like number of seeds per boll.
The gene action for number of seeds per boll came out to be partial dominance and additive type in F3 generation because the regression line is cutting the covariance axis above the origin. The non -significant deviation of the regression line from unit slope indicates the absence of non-allelic interaction. But the results produced in this recorded by Abo-El-Zahab and Methwaly (1979) and Singh et al. (1982) were different as they observed overdominance. The deviation with this respect to the phenotypic manifestation of this parameter might be due to different varieties utilized under different environmental conditions and because of segregation in F3 generation.
Lint Percentage (GOT): A glance at Table 3, indicates that the regression line intercepts the covariance axis above the origin and hence reflects the additive type of gene action. The locations of array points along the regression line indicates that both CIM-70 and NIAB-78 being nearest to the origin, get most dominant genes for lint percentage. While AUH-50 receives the most recessive genes being away from the origin. The significant deviation of the regression line from the unit slope indicates the presence of non allelic interaction. These results are in quite resemblance with those already reported by Khan and Ghafoor (1986) and Khan et al. (1991). But Kassem et al. (1986), Latif et al. (1988) and Rehman et al. (1988) contradicted with these findings. This type of contradiction might be due to different genetic materials and climatic conditions under which these experiments were performed. The conclusion of the results of the analysis of variance disclosed that the mean genotypic differences between hybrids and their parents in F3 generation were highly significant for the characters like; number of bolls, number of seeds per boll, Lint percentage and yield of seed cotton per plant. In case of staple length for which the level reduced to Non significant only. It can be conferred easily from Vr and Wr graphic presentation that the characters, number of bolls per plant, number of seeds per boll,and lint percentage were controlled by additive type of gene action. Moreover in the range of additive type of gene action the varieties possessing the most dominant genes are Cyto-21/91, Niab-78, Niab-78, CIM-70. This type of gene action provides out lines for reasonable expectation for success in isolating the superior genotypes. Whereas, overdominance type of gene action was predominant for yield of seed cotton per plant. Such type of gene action is less effective in selecting and fixing genotypes because overdominance effects can not be fixed except in F1 hybrids, or following a wide cross with chromosome doubling. So for as interaction of alleles is concerned, the non-significant deviation of regression line from unit slope indicates the absence of non-allelic interaction which was present for the traits under study . Lint percentage (GOT) showing significant deviation from unit slope which surely reflected the presence of epistasis.
Keeping in view the results of the genetic analysis study it is, therefore, strongly suggested that the number of bolls per plant, yield of seed cotton and seeds per boll may be made the criteria for selecting high yielding genotypes of cotton.
REFERENCES
- Whitehouse, R.N.H., J.B. Thompson and M.A.M. Do Valle Ribeiro, 1958. Studies on the breeding of self-pollinating cereals: 2. The use of a diallel cross analysis in yield prediction. Euphytica, 7: 147-169.
CrossRefDirect Link