
Zerihun Jalata
Department of Plant Sciences, Faculty of Agriculture, Wollega University, Ethiopia
LiveDNA: 251.24518
Firew Mekbib
School Plant Sciences, College of Agriculture and Environmental Sciences, Haramaya University, P.O. Box 138, Dire Dawa, Ethiopia
LiveDNA: 251.22688
Berhane Lakew
Ethiopian Institute of Agricultural Research (EIAR), Holetta Agricultural Research Center (HARC), P.O. Box 2003 or 20420, Code 1000, Addis Ababa, Ethiopia
Seid Ahmed
Biodiversity and Integrated Gene Management Program, International Center for Agricultural Research in the Dry Areas (ICARDA), Morocco
Journal of Applied Sciences
Year: 2019 | Volume: 19 | Issue: 2 | Page No.: 88-95







ABSTRACT
Background and Objectives: Barley is one the most important crops worldwide mainly for food, feed and drinks including Ethiopia. The aim of this study was to assess the combining ability of barley parents and estimate the inheritance pattern for some agro-morphological traits for future breeding. Materials and Methods: This study was carried out on 28 barley genotypes (21 F1 and 7 parents) in a randomized complete block design with three replications during 2015 cropping season at Holetta, Ethiopia. Results: There was significant difference due to both GCA and SCA among barley genotypes for all the characters studied except for days to heading, days to maturity and grain yield per plant which had non-significant SCA revealing the predominance of additive gene effects to non-additive gene effects for all characters except for thousand kernel weight. Every barley parents were best general combiners for one or more important agronomic characters. Large range of heterosis values (-23.8 to 86 and -40.6 to 43.2%) for grain yield per plant over mid parent and better parent, respectively. Conclusion: The finding suggests that yield can be improved through multiple crossing of parents with best GCA effects for the desired characters followed by selection at later generation.
PDF Abstract XML References Citation
Received: December 28, 2018;
Accepted: February 01, 2019;
Published: April 05, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Zerihun Jalata, Firew Mekbib, Berhane Lakew and Seid Ahmed, 2019. Gene Action and Combining Ability Test for Some Agro-morphological Traits in Barley. Journal of Applied Sciences, 19: 88-95.
DOI: 10.3923/jas.2019.88.95
URL: https://scialert.net/abstract/?doi=jas.2019.88.95
DOI: 10.3923/jas.2019.88.95
URL: https://scialert.net/abstract/?doi=jas.2019.88.95
INTRODUCTION
Barley (H. vulgare L.) is one of a major staple crop in the highlands of Ethiopia and it holds unique place in farming, contributes to food security as food, drinks, feed and other uses1. Barley production covers an area about 1 million ha with an estimated annual production of 2 Mt with national average productivity of about 2.1 t ha–1 in Ethiopia2, while the world barley average productivity level was about 3.0 t ha–1 even in some top producing countries exceeds3 5 t ha–1. In Ethiopian breeding program emphasis has been given to the evaluation of landraces and exotic germplasm for different purposes. To enrich and improve the germplasm base, exotic germplasm of both food and malt types have been introduced from ICARDA and other sources though introduced germplasm show poor in some agronomic traits and less adapted. Despite the efforts made so far to generate improved barley technologies yet there are several constraints of which the low yielding nature of the available cultivars among them4. Therefore, to undertake genetic improvement and achieve genetic gain the knowledge on the genetic background of the available cultivars is important. Thus, there were barley cultivars included in the present study which have several desirable characteristics, some were introduced from abroad and other released domestically, but their genetic background was not studied which otherwise could be useful in breeding programs. Hence, the evaluation of these cultivars is useful to generate information on the inheritance mechanism of traits and identify the suitability of the parents for hybridization which may be important to improve the efficiency of barley selection.
Grain yield is a complex quantitative trait which is an ultimate product of the action and interaction of a number of yield component characters. For an effective breeding program, aimed at improvement of complex characters such as yield requires a sound knowledge of the genetics of yield and its related characters is useful to make selection decisions and efficient management of breeding material. Hence, advances in the barley yield requires information on the combining ability of the parents available in the genetic material to be used in the hybridization program and on the nature of the gene actions involved in the expression of quantitative traits of major importance5. In addition the knowledge on nature and magnitude of gene effects controlling inheritance of agro-morphological characters related to crop productivity will in turn become helpful in formulating an effective and efficient breeding program6. General (GCA) and specific combining ability (SCA) effects are very effective genetic parameters in deciding the next phase of breeding programs. Diallel method of genetic analysis provides a unique opportunity to test a number of lines in all possible combinations5. The GCA effect is a good indication of genes with primarily additive effects while SCA is an indication of genes with dominance or epistatic effects7 and high GCA effects are mostly due to additive gene effects or additive×additive interaction effects8.
Research reports on barley showed that both GCA and SCA analysis in both F1 and F2 barley indicated highly significant for days to heading, plant height, spike length, number of grains per spike, thousand kernel weight and grain yield per plant. But the GCA:SCA ratio revealed the preponderance of non-additive gene effects for these traits5. Another diallel analysis showed that seed yield, days to heading and spike length were controlled by the dominance gene effects and spike weight, thousand seed weight as well as spike length conditioned by over-dominance effect9. Therefore, development of high yielding cultivars with desirable traits are required to improve the yielding potential of the existing cultivars which are adaptable to different production systems1,4. Hence, genetic characterization of the breeding materials is useful to improve the breeding efficiency and to achieve genetic gain. Therefore, in the present study, seven barley cultivars with different desirable traits and origin or adaptation were evaluated in a 7×7 half diallel fashion with the aim of assessing the combining ability of barley parents and estimate the inheritance pattern for some agro-morphological traits for future breeding.
MATERIALS AND METHODS
Site description: The study was conducted at Holetta Agricultural Research Center which is about 30 km west of Addis Ababa, Ethiopia, in 2015 main season. Holetta is located at an altitude of 2390 m above sea level (m.a.s.l), latitude of 09°04'N and longitude of 38°30'E. Most of the national barley breeding experiments are carried out at this center. The maximum and minimum temperature of Holetta is 22.2 and 6.13°C, respectively. While average rainfall is about 1100 mm and the soil types are vertisol and nitisol (http://www.eiar.gov.et).
Experimental design and methods of planting methods: Seven barley parents (Table 1) with diverse agronomic characters and origin were crossed in all possible cross combination, excluding reciprocals, in 2014/15 main season by hand emasculation and pollination in the field to obtain 21 F1 cross seeds. Seeds of the 21 F1 and their parents were sown in the field at Holetta in a randomized complete block arrangement each with three replications in the 2014/15 main cropping season.
| Table 1: | Description of seven barley cultivars used in half diallel crossing in 2014/15 |
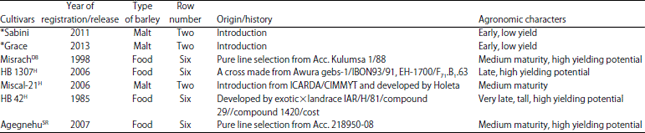 | |
*Introduced (personal communication with Dr. Berhane Lakew),DBDeveloped by Holetta Agricultural Research Center and released by Debre Berhan Agricultural Research Center, HReleased and developed by Holeta Agricultural Research Center, SRReleased and developed by Sirinka Agricultural Research Center. Sources: Lakew and Alemayeh16 and Abtew et al.17 | |
Each genotype was sown in a plot with two rows of 2.5 m length and 0.40 m width at 0.15 m between plants in rows. All recommended agronomic and cultural practices are applied.
Data collection: Ten random plants in the parents and F1s progenies were taken for recording observations on days to heading, days to maturity, spike length, number of grains per spike, thousand kernel weight and grain yield per plant.
Statistical analysis: Analysis of variance (ANOVA) was performed following GLM procedures of Statistical Analysis Systems10 Software program. Significance test of the characters were determined before diallel analysis. Then diallel analysis was performed according to Griffing’s Method 2 and model I (fixed effect)8 using the SAS program for Griffing’s diallel analysis11,12. The combining ability ratio13 was used to predict the most gene action controlling the traits. Mid parent heterosis (MPH), better parent heterosis (BPH) or heterobeltiosis was estimated14 and significance tests were made using the method of Wynne et al.15.
RESULTS
Analysis of variance and mean performance for yield related traits and yield: Analysis of variance for all traits studied in the present study showed significant (p<0.01) difference among the genotypes (7 parents+21 F1 crosses) (Table 2). Mean performance of barley parents and their hybrids for some yield and yield a component is presented in Table 3. Sixteen F1 hybrids showed greater performance than parental mean yield (26.39 g). Barley hybrid HB1307×HB42 (58.5 g) showed 102% greater than the parental mean followed by Grace×Misrach hybrid (40.6 g). For thousand kernel weight, all F1 hybrids gave greater than the parental mean (41.8 g). Only 6 out of 21 hybrids had more number of kernels/spike than parental mean (40). For spike length, 18 hybrids had better than parental mean. However, few genotypes gave lower mean performance for days to heading and days to maturity than parental means (Table 3).
Combining ability effects: Analysis of variance for combining ability (Table 2) showed that the variance due to GCA and SCA was highly significant (p<0.01) for all characters studied except days to heading, days to maturity and grain yield per plant for which SCA was non-significant. Baker ratio was closer to unity for all traits studied except thousand kernel weight and narrow semse heritability estimates (Table 2) for all the parameters except thousand kernel weight was high. The GCA estimates of HB1307 and HB42 parents showed highly significant GCA effect for grain yield per plant and number of kernels per spike; HB42 and Miscal-21 for thousand kernel weight, Sabini, Misrach and Agegnehu parents for days to heading and days to maturity and Grace and Sabini for spike length as well as Misrach and Agegnehu for number of kernels per spike (Table 4).
The usefulness of a particular cross in exploiting heterosis is judged mainly by the SCA effect of component genotypes. The SCA effects for different yield and yield related traits for F1 hybrids are shown in Table 5. Hence, the cross HB1307×HB42 followed by Grace×Misrach proved to be the best specific combinations to improve grain yield per plant. The combinations such as Grace×HB1307, Grace×HB42, Grace×Misrach, Sabini×HB42, Sabini×HB1307, Sabini×Misrach, Miscal-21×HB42, HB1307×Miscal-21 and Misrach×Miscal-21 can be used to develop desirable progenies for increased thousand kernel weight (Table 5). The cross combinations which were significant and desirable for the development of improved spike length were Grace×HB1307, Grace×HB42, Grace×Misrach, Sabini×HB42, Sabini×HB1307, Grace×Miscal-21, HB1307×Miscal-21 and Misrach×Miscal-21 (Table 5).
| Table 2: | Analysis of variance for some yield related traits and yield in 7×7 half diallel cross of barley |
 | |
DF: Degree of freedom, DH: Days to heading, PH: Plant height (cm), SPL: Spike length (cm), Nk/Spk: Number of kernels per spike, TKW: Thousand kernel weight (g), GY/Pl: Grain yield per plant weight (g), CV(%): Coefficient of variation (%), GCA: General combining ability, SCA: Specific combining ability, M’e: Error variance for combining ability, h2n: Narrow sense heritability | |
| Table 3: | Mean performance of barley parents and their hybrids for some yield components and yield |
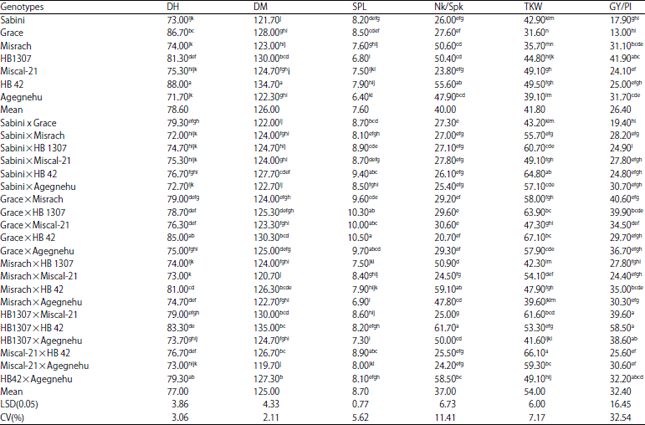 | |
| SE: Standard error, means in the same column with the same letters are not significantly different at 0.05 level of probability | |
| Table 4: | General combining ability of parents for some yield related traits and yield in 7x7 half diallel cross of barley |
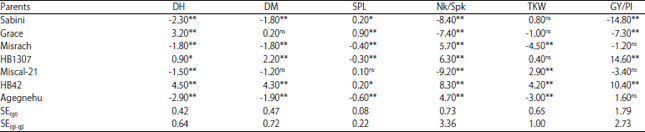 | |
*,**Significant at the 0.05 and 0.01 probability level, respectively, ns: Non significance, SE(gi): Standard error for all GCA, SE(gi-gi): Standard error for testing the significance among two superior significant GCA effects | |
| Table 5: | Estimated specific combining ability (SCA) for some yield related and yield traits in 21 barley F1 hybrids |
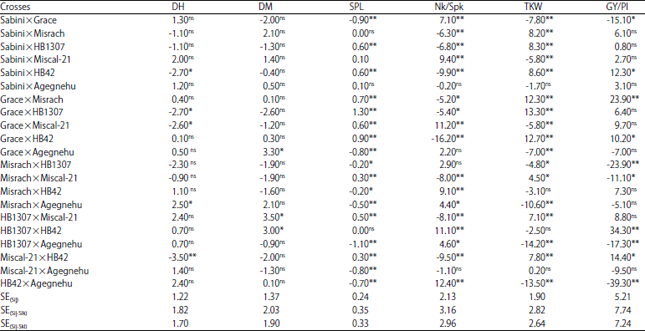 | |
*,**Significant at the 0.05 and 0.01 probability level, respectively, ns: Non significance, S.E(Sij): Standard error for testing all SCA, S.E(Sij-ik): Standard error for comparing the difference between two SCA having one common parent, S.E (Sij-kl): standard error for testing the SCA of two crosses having no parent in common | |
| Table 6: | Estimation of heterosis (%) for 7 traits over mid parent (MPH) and better parent (BPH) in 7x7 half diallel cross of barley |
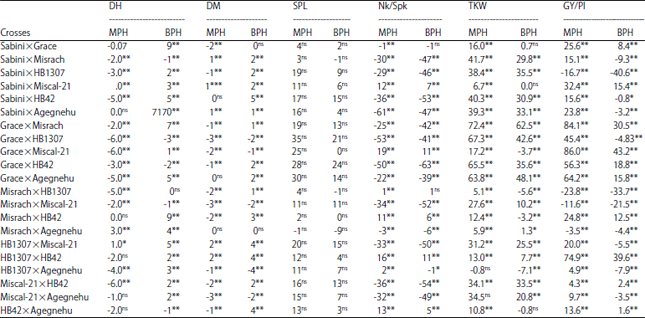 | |
*,**Significant at the 0.05 and 0.01 probability level, respectively, ns = Non significance, DH: Days to heading, DM: Days to maturity, SPL: Spike length (cm), Nk/Spk: number of grains per spike, TKW: Thousand kernel weight (g), GY/Pl: Grain yield per plant weight (g) | |
Heterosis of barley: Heterosis estimates of barley F1 hybrids over the mid and better parent for some yield components and yield were shown in Table 6. Thus, except spike length, highly significant heterosis over both mid parent and better parent was recorded in almost all F1 hybrids in both directions. Heterosis values for grain yield per plant varied from -23.8 to 86 and -40.6 to 43.2% for mid parent and better parent heterosis or heterobeltiosis, respectively (Table 6). Mid parent and better parent heterosis estimates ranged from the lowest value of -0.8% to a maximum of 72.4 and -7.1 to 62.5%, respectively, for thousand kernel weight. Despite the fact that most of crosses showed negative heterosis for number of kernels per spike (Table 6) but cross combination among two rowed barleys Grace×Miscal-21 showed maximum (19%) heterosis over mid parent followed by cross between six rowed, HB1307×HB42 (16%) and the same cross had 11% heterosis over better parent for number of kernels per spike. Regarding spike length, heterosis over mid parent ranged from -1% (Misrach×Agegnehu) to 35% (Grace×HB1307) whereas, the heterosis over better parent showed a maximum of 24% (Grace×HB42). Most of the heterosis for days to heading and days to maturity characters were highly significantly negative which is in the desirable direction (Table 6).
DISCUSSION
Yield improvement can be achieved through multidimensional strategies and selection of superior parents for hybridization and selection area important activities of plant breeder. So it is important to ensure the suitability of parents for breeding by investigating the inheritance of genes for different desirable characters. Hence, in the present the analysis of variance for all traits studied showed highly significant (p<0.01) (Table 2) difference among the genotypes indicating the presence of wide genetic variability for improvement. Similarly, significant differences between F1 barley hybrids and F2 progenies were obtained for all the traits except for spike length5 in F2 and also the mean performances of parents and their crosses for various studied traits showed high variation. Furthermore, the existence of significant (p<0.01) difference for general combining ability (GCA) and specific combining ability (SCA) for all characters except days to heading, days to maturity and grain yield per plant indicated the importance of both additive and non-additive gene effects in the inheritance of these traits in barley. In another study on barley highly significant GCA and SCA was reported for days to heading, days to maturity, spike length, number of grains per spike and grain yield per plant in F1 and F2 generations5,18. In contrary the report of non-additive (non-fixable) gene effects governing the inheritance of number of grains per spike and grain yield also stated19,20. Inheritance was controlled by additive genes in both F1 and F2 barley generations for spike length21. It is partly coincided with the report of Eshghi and Akhundova6 on inheritance of both additive and non-additive genes for thousand kernel weight but only additive gene effect for number of kernels per spike under non-drought stress condition. Moreover it partly coincides with reports on seed yield, days to heading and spike length were controlled by dominance effects of genes but thousand seed weight and spike length were controlled by over-dominance effect in barley9.
Furthermore, Eshghi and Akhundova22 also indicated additive genets for days to maturity. This study coincided with the report of Assef and Labuschagne23 for grain yield, days to heading and days to maturity under both drainage free and water logging. The findings of Verma et al.24 on the number of grains per spike and grain yield which were controlled by additive gene effects in barley. In contrast to this study non-additive gene action involved in controlling days to flowering, days to maturity, spike length, grains per spike, thousand kernel weight and grain yield in barley25. As Baker13 suggested that the combining ability ratio is useful to assess the relative importance of GCA and SCA in determining progeny performance so that as the ratio closer to unity, the greater the predictability of selecting superior progeny based on estimates of GCA alone. In this study, the Baker ratio was closer to unity for all traits studied except thousand kernel weight and similarly narrow semse heritability estimates for the same characters except thousand kernel weight were high, Table 2 showing further evidence of predominance of additive genes ((fixable) to non-additive gene effects conditioning the characters so that selection would be effective in F2 and later generations. The narrow heritability is a measure of fixable component of variance through selection leading to increased magnitude of quantitative traits and so it is a measure of the predictability of offspring trait values that is based on parental trait values26.
The GCA estimates revealed that HB1307 and HB42 were best general combiners for grain yield per plant and number of kernels per spike; HB42 and Miscal-21 for thousand kernel weight; Sabini, Misrach and Agegnehu parents for days to heading and days to maturity and Grace and Sabini for spike length as well as Misrach and Agegnehu for number of kernels per spike (Table 4). In general this finding suggested every parent was best general combiner for one or more agronomic traits indicating that every parent has a potential to significantly contribute desirable genes to their progenies. Thus the best progeny is produced, when parents with the highest GCA for a specific trait are used. Except Grace×Misrach cross, the best performing crosses for grain yield per plant in this study had at least one parent with highly significant GCA effects27. Large extent of mid parent and better parent heterosis or heterobeltiosis values was obtained for grain yield per plant and other important agronomic traits (Table 6) in both directions showing significant involvement of non-additive gene effects in heterosis of barley. Both positive and negative heterosis can be useful depending on the type of trait. Hence, F1 hybrids with positive heterosis are desirable for increased yield, thousand kernel weight, spike length and number of grains per spike. On the contrary, negative values of heterosis are desirable in the case of days to heading, days to maturity and plant height of barley traits28. Similar findings on barley showed that the average heterosis values of F1's based on mid parent means for yield, kernel weight, kernels per head and height were 21.5, 5.9, 7.1 and 3.2%, respectively and heterosis over better parent for yield was 9.1% in barley29. Absence of significant heterosis in some traits could be due to the internal cancellation of heterosis components. Heterosis is mainly a function of non-additive (dominance and epistasis) genetic effects14,30. Desirable heterosis will be achieved when better-parent heterosis value is positive for all studied characters; except days to heading that have a negative heterosis would be useful for earliness breeding31.
Some studies recommend that the use of biparental mating, population development, diallel selective mating of segregants methods as useful for utilizing both additive and non-additive genes. Bi-parental mating offers good opportunity for increasing the frequency of genetic recombinants speeding up the rate of genetic improvement. Population breeding is also suggested in the form of bi-parental mating between selected recombinants to exploit the additive and non-additive effects18,32. Bi-parental mating and/or diallel selective mating allows the intermating of selected lines in different cycles and used to exploits both additive and non-additive gene effects in barley5. To exploit non-additive gene action, one or two cycles of inter-mating or some form of population improvement schemes may be employed in later generation25. A population may be developed with most of the favorable genes accumulated, make use of multiple parents, which are good general combiners for several characters, in multiple crossing programs. Diallel selective mating system is a good technique which delays the fixation of gene complexes, permit breakdown of linkages, fosters recombination and concentrates favorable genes or gene complexes in to central gene pool by a series of multiple crosses18.
CONCLUSION
The combining ability analysis, Baker ratio and narrow sense heritability confirmed the preponderance of additive gene effects over non-additive effects in all the agronomic characters except for thousand kernel weight which revealed the potential success for selection. All parents were best general combiners for one or more desirable traits which imply the importance of population improvement schemes. High heterosis values over mid parent and better parent was obtained for grain yield per plant and yield components. Hence the result suggests that improvement in yield can be achieved through multiple crossing of parents with best GCA effects for the desired traits followed by selection at later generation.
ACKNOWLEDGMENTS
The authors would like to thank the Ethiopian Ministry of Education (MOE), Haramaya University and Wollega University for financing our research. The Ethiopian Institute of Agricultural Research, Holeta Agricultural Research Center, Barley Breeding Program, is acknowledged for their provision of research facilities and experimental land to carry out research activities.
REFERENCES
- Mulatu, B. and B. Lakew, 2011. Barley Research and Development in Ethiopia: An Overview. In: Barley Research and Development in Ethiopia, Mulatu, B. and S. Grando (Eds.). Ethiopian Institute of Agricultural Research, International Center for Agricultural Research in the Dry Areas (ICARDA), Aleppo, Syria, ISBN: 92-9127-249-3, pp: 1-16.
- Sharma, Y., S.N. Sharma, P. Joshi and R.S. Sain, 2003. Combining ability in the F1 and F2 generations of a diallel cross in six-rowed barley (Hordeum vulgare L.). Acta Agron. Hung., 51: 281-288.
CrossRefDirect Link - Eshghi, R. and E. Akhundova, 2010. Genetic diversity in hulless barley based on agromorphological traits and RAPD markers and comparison them with storage protein analysis. Afr. J. Agric. Res., 5: 97-107.
Direct Link - Sprague, G.F. and L.A. Tatum, 1942. General vs. Specific combining ability in single crosses of corn. J. Am. Soc. Agron., 34: 923-932.
CrossRefDirect Link - Griffing, B., 1956. A generalised treatment of the use of diallel crosses in quantitative inheritance. Heredity, 10: 31-50.
CrossRefDirect Link - Aghamiri, S.M.M.S., K. Mostafavi and A. Mohammadi, 2012. Genetic study of agronomic traits in barley based diallel cross analysis. Adv. Environ. Biol., 6: 62-68.
Direct Link - Zhang, Y., M.S. Kang and K.R. Lamkey, 2005. DIALLEL-SAS05: A comprehensive program for Griffing's and Gardner-Eberhart analysis. Agron. J., 97: 1097-1106.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Wynne, J.C., D.A. Emery and P.W. Rice, 1970. Combining ability estimates in Arachis hypogaea L. II. Field performance of F1 hybrids. Crop Sci., 10: 713-715.
CrossRefDirect Link - Abtew, W.G., B. Lakew, B.I. Haussmann and K.J. Schmid, 2015. Ethiopian barley landraces show higher yield stability and comparable yield to improved varieties in multi-environment field trials. J. Plant Breed. Crop Sci., 7: 275-291.
Direct Link - Kakani, R.K., S. Yogendra and S.N. Sharma, 2007. Combining ability of barley genotypes in diallel crosses. SABRAO J. Breed. Genet., 39: 117-126.
Direct Link - Ciulca, S., C. Adriana, E. Madosa and V. Giancarla, 2012. Diallel analysis of variance-covariance regression for spike length in six-row winter barley. J. Hortic. For. Biotechnol., 16: 82-86.
Direct Link - Eshghi, R. and E. Akhundova, 2009. Genetic analysis of grain yield and some agronomic traits in hulless barley. Afr. J. Agric. Res., 4: 1464-1474.
Direct Link - Assef, A. and M.T. Labuschagne, 2013. Expression of traits of barley (Hordeium vulgare L.) landrace crosses under waterlogged and free drainage conditions. Global J. Sci. Front. Res., 13: 1-14.
Direct Link - Verma, A.K., S.R. Vishwakarma and P.K. Singh, 2007. Genetic architecture for yield and quality component traits over two environments in barley (Hordeum vulgare L.). Barley Genet. Newslett., 37: 24-28.
Direct Link - Pawar, K.K. and A.K. Singh, 2013. Combining ability analysis for grain yield and its attributing traits in barley. Int. J. Agric. Sci. Vet. Med., 1: 83-87.
Direct Link - Kholghi, M., H.H. Maleki and R. Darvishzadeh, 2014. Diallel analysis of yield and it's related traits in sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Agric. Conspectus Scient., 79: 175-181.
Direct Link - Choudhary, A.K., L.B. Chaudhary and K.C. Sharma, 2000. Combining ability estimates of early generation inbred lines derived from two maize populations. Indian J. Genet. Plant Breed., 60: 55-61.
Direct Link - Upadhyaya, B.T. and D.C. Rasmusson, 1967. Heterosis and combining ability in barley. Crop Sci., 7: 644-647.
CrossRefDirect Link - Rizkalla, A.A., B.A. Hussien, A.M.F. Al-Ansary, J.E. Nasseef and M.H.A. Hussein, 2012. Combining ability and heterosis relative to RAPD marker in cultivated and newly hexaploid wheat varieties. Aust. J. Basic Applied Sci., 6: 215-224.
Direct Link - Farshadfar, E., F. Rafiee and A. Yghotipoor, 2012. Comparison of the efficiency among half diallel methods in the genetic analysis of bread wheat (Triticum aestivum L.) under drought stress condition. Ann. Biol. Res., 3: 1607-1622.
Direct Link