
Mohamed Farouk El Karamany
Department of Field Crops Research, Agricultural and Biological Division, National Research Centre, P.O. Box 12622, Giza, Egypt
Mervat Shamoon Sadak
Department of Botany, Agricultural and Biological Division, National Research Centre, P.O. Box 12622, Giza, Egypt
LiveDNA: 251.26733
Bakry Ahmed Bakry
Department of Field Crops Research, Agricultural and Biological Division, National Research Centre, P.O. Box 12622, Giza, Egypt
LiveDNA: 20.20713
Journal of Applied Sciences
Year: 2019 | Volume: 19 | Issue: 7 | Page No.: 718-724







ABSTRACT
Background and Objective: Mung bean is an important pulse food and is commonly used in cooking using green and mature pods. Plant growth regulators (PGRs) are known to influence plant growth and development at very low amounts. Thus, this investigation was done to study the synergistic role of IAA and GA3 on growth, some biochemical aspects and yield of mung bean. Materials and Methods: A field experiment was carried out in Researches and Production Station of National Research Centre, Al-Nubaria district, Al Beheira Governorate, Egypt, at two summer seasons. Seeds of mung bean were soaked for 12 h in different concentrations of bio regulator indole acetic acid (IAA) and/or gibberellic acid (GA3) with different concentrations. The experiment design was complete randomized block design in three replicates. Results: Data indicated that all treatments caused significant increases in morphological criteria, biochemical parameters and yield of mung bean plants. IAA or GA3 with 50 mg L–1 concentration recorded the best values growth photosynthetic pigments as well as, endogenous auxins, GA3 and cytokinins. Interaction of IAA 50 mg L–1+GA3 50 mg L–1 gave the most effective treatment. The superiority of treatment IAA 50, GA3 50 mg L–1 and interaction of IAA 50 mg L–1+GA3 50 mg L–1 recorded the same trend due to yield and yield components in characters plant height, pods No./plant, pods weight/plant, seeds No./pod, seed yield, straw yield, biological yield, total carbohydrates, protein in seeds (%) and N in seeds (%). Conclusion: It could be concluded that soaking of mung bean seeds in different concentrations of IAA and/or GA3 increased growth and yield of mungbean. About 50 mg L–1 IAA+50 mg L–1 GA3 was the most effective treatment on increasing growth and yield of mung bean.
PDF Abstract XML References Citation
Received: January 16, 2019;
Accepted: March 02, 2019;
Published: July 24, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mohamed Farouk El Karamany, Mervat Shamoon Sadak and Bakry Ahmed Bakry, 2019. Synergistic Effect of Indole Acetic Acid and Gibberellic Acid on Mung Bean Grown under Sandy Soil Conditions. Journal of Applied Sciences, 19: 718-724.
DOI: 10.3923/jas.2019.718.724
URL: https://scialert.net/abstract/?doi=jas.2019.718.724
DOI: 10.3923/jas.2019.718.724
URL: https://scialert.net/abstract/?doi=jas.2019.718.724
INTRODUCTION
Mung bean (Vigna radiata L. Wilczek) is a summer pulse crop with short duration (70-90 days) and double use (forage and seed) and high nutritive value. Forage contains 12-18% protein but seeds contain 22-28% protein, 60-65% carbohydrates, 1-1.5% fats, 3.5-4.5% ash)1. Mung bean is commonly used in cooking using green and mature pods. Sprouts are rich in vitamins and amino acids also, in broilers diets as un-traditional feeding stuff or its husks as crop by-products for feeding rabbits2. Plant growth regulators (PGRs) are known to influence plant growth and development at very low concentrations. Moreover, the response of plant to PGRs may vary with species, varieties, environmental conditions, physiological and nutritional status, stage of development and endogenous hormonal balance3 delays senescence4. Plants have the ability to store excessive amounts of exogenously supplied hormones in the form of reversible conjugates which release active hormone when the plants need them during the growth period5. Amanullah et al.6 mentioned that plant growth substances are known to enhance the source-sink relationship and stimulate the translocation of photo-assimilates to sink thereby helping in effective flower formation, fruit and seed development and ultimately enhancing the productivity of crops. Various plant growth regulators which regulate growth under normal or stress conditions are auxins. Generally, auxins are the best hormones for use because they are non-toxic to plants over a wide range of concentration and effective in promoting root system of large number of plant species. Auxins might regulate cell elongation, cell division, tissue swelling, formation of adventitious roots, callus initiation, induction of embryogenesis and promoting cell wall loosening at very low concentrations7. The principal auxin in plants is indole-3-acetic acid (IAA) that is produced mainly in the shoot apex bud and young leaves of plants. Other meristematic tissues, flowers, fruits and young seeds have also been shown to be sites of this hormone production8.
Gibberellins (GAs) are the most important natural growth regulators and generally involved in growth and development of different plant. They control seed germination, leaf expansion, stem elongation and flowering9. Gibberellic Acid (GA) is used to induce great changes in the growth characters, chemical composition and yield criteria of plant10. The IAA and/or GA3 have stimulative role on increasing growth and yields of mung bean plant. So, this study was an attempt to improve plant growth and yield quantity and quality of mung bean plant under new reclaimed sandy soil, through improving some physiological and biochemical processes by using naturally occurring substances IAA and GA3.
MATERIALS AND METHODS
Experimental design: Two field experiments were carried out during summer seasons of 2016 and 2017 from June till August in Researches and Production Station of National Research Centre (NRC), Al-Nubaria district, Al Beheira Governorate, Egypt. The experimental soil before sowing was analyzed according to Chapman and Pratt 11. Soil texture was sandy and its characteristics are shown in Table 1.
Temperature averages (°C) during trial period were June (25.60)-July (25.80)-August (27.2). Humidity average (%) June (76.3%)-July (77.0%)-August (76.9%), respectively.
Seeds of mung bean were soaked for 12 h in different concentrations of bio regulator indole acetic acid (IAA) and/or gibberellic acid (GA3) with different concentrations as follows:
| • | Control (IAA 0.0 mg L–1, GA3 0.0 mg L–1) |
| • | IAA 25 mg L–1 |
| • | IAA 50 mg L–1 |
| • | GA3 25 mg L–1 |
| • | GA3 50 mg L–1 |
| • | IAA 25mg L–1+GA3 25 mg L–1 |
| • | IAA 25 mg L–1+GA3 50 mg L–1 |
| • | IAA 50 mg L–1+GA3 25 mg L–1 |
| • | IAA 50 mg L–1+GA3 50 mg L–1 |
The experiment design was complete randomized block design in three replicates. Area of each plot was 10.5 m2 (6 rows×0.50 m width×3.5 m length). Seeds of mung bean variety (kawmy-1) inoculated with specific strain of bacteria and hand seeded in hills 0.30 m between (2 seeds/hill) on the 15 and 17 June in both seasons. The soil was immediately irrigated after sowing using sprinkler irrigation system. Fertilization of NPK at rate of 33:32:24/feddan (4200 m2) in the form of ammonium nitrate 33%N, superphosphate 15.5% P2O5 and potassium sulfate 48% K2O. Other cultural practices were done due to those recommended for mung bean.
| Table 1: | Mechanical and chemical analysis of experimental soil |
Data recorded: Plant samples were taken after 60 DAS from sowing random sample of 0.5×0.5 m2 was cut for determination of growth criteria (shoot length, branches and leaves No./plant, leaves fresh dry weight/plant) and estimation of some biochemical parameters in leaves of plant such as photosynthetic pigments and endogenous phytohormones. At harvest (90 DAS) 10 plants from each central plot were randomly take for yield components measure but the whole yield of each plot (10.5 m2) was harvested for characters measure per feddan and then the following characters were determined: (1) Plant height (cm), (2) Pods No./plant, (3) Pods weight/plant, (4) Seeds No./pod, (5) Seed yield (t/feddan), (6) Straw yield (t/feddan), (7) Biological yield (t/feddan), (8) Total carbohydrates (%), (9) Protein in seeds (%) and (10) N in seeds (%).
Feddan = Fed. (Local unit area equal (4200 m2)
Measurements
Photosynthetic pigments: Total chlorophyll a and b and carotenoids contents in fresh leaves were estimated using the method12. The fresh tissue was ground in a mortar and pestles using 80% acetone. The Optical Density (OD) of the solution was recorded at 662 and 645 nm (for chlorophyll a and b, respectively) and 470 nm (for carotenoids) using a spectrophotometer (Shimadzu UV-1700, Tokyo, Japan). The values of photosynthetic pigments were expressed in mg g–1 FW.
Endogenous growth regulators: Based on the results of first season (2016), the most promising treatments were selected for determination of endogenous regulators. These endogenous growth hormones namely auxins, gibberellic acid, abscisic acid (ABA) and cytokinins were extracted according to Shindy and Smith13. The IAA, GA and ABA contents were determined by Gas Liquid Chromatography (GLC) according to the method described13 and cytokinin content was determined by High Performance Liquid Chromatography (HPLC) according to the method described14.
Total carbohydrate: Determination of total carbohydrates was carried out according to Herbert et al.15. A known mass (0.2-0.5 g) of dried tissue was placed in a test tube and then 10 mL of sulphuric acid (1 N) was added. The tube was sealed and placed overnight in an oven at 100°C. The solution was then filtered into a measuring flask (100 mL) and completed to the mark with distilled water. The total sugars were determined colorimetrically according to the method16 as follows: An aliquot of 1 mL of sugar solution was transferred into test tube and treated with 1 mL of 5% aqueous phenol solution followed by 5.0 mL of concentrated sulphuric acid. The tubes were thoroughly shaken for 10 min then placed in a water bath at 23-30°C for 20 min. The optical density of the developed color was measured at 490 nm using Shimadzu spectrophotometer model UV 1201.
Nitrogen and crude protein: Nitrogen and protein contents were determined with micro Kjeldahl’s apparatus according to the method17. Crude protein was determined according to Bradford18 (multiplying nitrogen contents by 5.75).
Statistical analysis: The obtained results were subjected to statistical analysis of variance according to method described19, since the trend was similar in both seasons the homogeneity test Bartlett's equation was applied and the combined analysis of the two seasons was calculated using SAS software version 9.32 as two factors-factorial analysis. Means were compared by using least significant difference (L.S.D.) at 5% levels of probability.
RESULTS
Changes in growth criteria: Data of the vegetative growth criteria of mung bean under sandy soil are presented in Table 2. The obtained results showed that, application of the two growth regulators (IAA and GA3) individually (Table 2) or in combination (Table 2) caused significant increases in growth criteria (shoot length (cm), branches and leaves No./plant, leaves fresh dry weight/plant (g)) of mung bean plant as compared with control plants. The highest values of growth criteria were obtained from the application of 50 mg L–1 IAA+50 mg L–1 GA3 treatment as compared with those obtained from the control and other treatments. Regarding IAA treatments individually, it was found that treating mung bean by 50 mg L–1 IAA was the most effective treatment. Also, the most effective GA3 treatment individually was 50 mg L–1.
Changes in photosynthetic pigments: The changes in chlorophyll a, chlorophyll b, carotenoids and total photosynthetic pigments in response to IAA and/or GA3 treatments are shown in Table 3. Data revealed that IAA and/or GA3 treatments caused significant increases in photosynthetic pigment constituents as compared with control. The highest values of photosynthetic pigments were recorded by treatment of 50 mg L–1 IAA+ 50 mg L–1 GA3.
| Table 2: | Effect of IAA or GA3 and IAA+GA3 (0, 25 and 50 mg L–1) on growth parameters of mung bean plants grown under sandy soil |
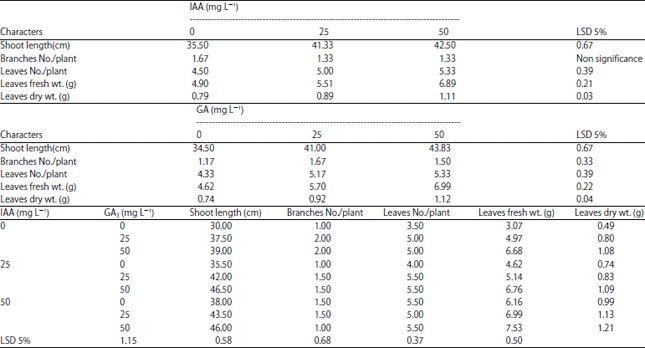 | |
| Table 3: | Effect of IAA or GA3 and IAA+GA3 (0, 25 and 50 mg L–1) on photosynthetic pigments (μg/100 g fresh weight) of mung bean plants grown under sandy soil |
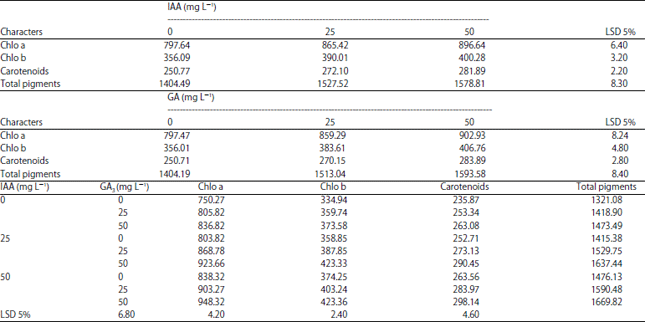 | |
| Chlo: Chlorophyll | |
Endogenous phytohormones: Based on the obtained results of the first season 50 mg L–1 GA3 and 50 mg L–1 IAA as individual treatment as well as 50 mg L–1 GA3+50 mg L–1 IAA as mixture treatment were selected to determine the endogenous phytohormones content as shown in Table 4. Individual treatments of 50 mg L–1 of both GA3 or IAA as well as combined treatment of 50 mg L–1 GA3+50 mg L–1 IAA caused increases in IAA, GA3 and cytokinin (zeatin) contents concomitant with decline in ABA contents of mung bean plants as compared with control plant. Using the combined treatment of both GA3+IAA with 50 mg L–1 was the most effective treatment as shown in Table 4.
| Table 4: | Effect of IAA and GA3 (0 and 50 mg L–1) on endogenous phytohormones (μg/100 g fresh wt.) of mung bean plants grown under sandy soil |
 | |
| Table 5: | Effect of IAA, GA3 and IAA+GA3 (0, 25 and 50 mg L–1) on yield, yield attributes and chemical values of mung bean plant grown under sandy soil |
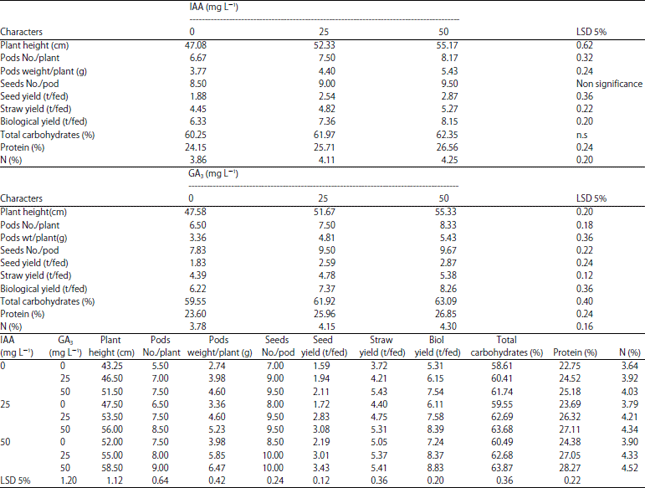 | |
Yield and yield attributes: Data in Table 5 indicated that IAA and/or GA3 treatments caused increases in yield and yield attributes (plant height, pods No./plant, pods weight/plant, seeds No./pod, seed weight, straw and biological yield/feddan) of mung bean. Furthermore, the highest values of yield were obtained from application of 50 mg L–1 IAA+50 mg GA3. Soaking mung bean seeds in different concentrations of IAA or GA3 individually increased markedly yield of mungbean.
Chemical composition of the yielded seeds: Data in Table 5 revealed that IAA and/GA3 treatments significantly increased total carbohydrate, protein and Nitrogen contents in the yielded seeds of the mung bean plant compared with the control plants. Treatment of 50 mg L–1 IAA+50 mg L–1 GA3 was the most effective treatment as it caused the highest total carbohydrate and protein contents of the yielded seeds.
DISCUSSION
Foliar treatment of IAA and/or GA3 caused marked increases in growth parameters of mung bean. These increments are similar to those reported8,20 on different plant species. The IAA or GA3 are considered two major plant growth regulators that were reported to augment growth and development of plants by stimulating a wide range of processes, including cell elongation and tissue growth, phototropism and gravitropism, apical dominance, lateral root initiation, differentiation of vascular tissues, embryogenesis, senescence, fruit setting and ripening7,21. In addition, the promoting effect of IAA may be attributed to enlarging leaves and increasing photosynthetic activities as obtained3 in the present study increasing cell division and accumulation of building units accompanied by greater polysaccharides and total carbohydrates content8.
Findings of this study revealed the positive effect of IAA and/or GA3 on photosynthetic pigments constituents. These obtained results of different treatments are similar to that obtained20 on maize. Jacobs22 found that, IAA presumably acts as a coenzyme in the metabolism of higher plants, thus it plays an important role in the formation of the photosynthetic pigments. These increases in pigments content may be attributed to the promotion of pigments synthesis and/or retardation of its degradation. The IAA and/or GA3 foliar treatment induced marked increases in IAA, GA3 and cytokinins, meanwhile decreased ABA contents of mung bean plants with are similar to those obtained8,23,24 on cowpea, wheat and faba bean plant. These increases in different growth bioregulators could be attributed to the enhancement in their biosynthesis and/or decreases in their degradation7. As well as, different treatment improve endogenous phytohormones via decreasing activity of degrading enzymes as well25. The decrease in ABA contents might be referred to the shift of the isopentenyl pyrophosphate which is the common precursor into cytokinin and/or gibberellins biosynthesis instead of ABA26.
This research indicated that IAA and/or GA3 treatments caused increases in yield and yield attributes. In this concern, Ammanullah6 reported that plant growth substances enhance the source-sink relationship and stimulate the translocation of photo-assimilates to sink thereby helping in effective flower formation, fruit and seed development and ultimately enhancing the productivity of crops. Abdoli et al.27 mentioned that IAA is the major naturally occurring auxin that increases stem elongation, cell expansion, growth rate and yield. Gibberellins are one of the main regulators of plant growth and development and they normally act by signaling the removal of proteins that repress growth thus promoting cell proliferation and elongation28. The promoting effect of Gibberellic acid may be ascribed to stimulating the mobilization of nutrients towards the buds thereby increasing cell division and/or increasing the differentiation of the vascular. Also, the increases in the yield of mung bean plants by application of different treatments relative to untreated plant might result through breaking the apical dominance of mung bean plants leading to the increase in flowering branches and consequently the number of fruits. The increases in seed weights might be due ascribed to the promotive effect of IAA and/or GA3 in increase the assimilates and their translocations from leaves to the fruits where the seed weight increased29,30.
Results also exhibited that IAA and/GA3 treatments significantly increased total carbohydrate, protein and Nitrogen contents in the yielded seeds of the mungbean plant compared with the control plants. Treatment of 50 mg L–1 IAA+50 mg L–1 GA3 was the most effective treatment as it caused the highest total carbohydrate and protein contents of the yielded seeds. These results agreed with some the findings of Sadak et al.8, Ibrahim et al.31 and Choudhury et al.32.
CONCLUSION
It can be concluded that treatment of IAA 50 mg L–1+GA3 50 mg L–1 was effective treatment to increase mung bean growth, photosynthetic pigments, endogenous phytohormones, yield, yield components and chemical composition of the yielded seeds as low input technology for improving yield of mung bean. However, complementary research on mung bean is needed as double purpose crop (forage and seed).
SIGNIFICANCE STATEMENT
This study confirmed that using IAA and/or GA3 could be beneficial for improving growth and yield on mung bean plant under sandy soil. This study will help the researchers to uncover the critical areas of hidden potential of IAA and/or GA3 that many researchers were not able to explore. Thus a new theory on efficient utilization of growth regulators for better growth of plants may be arrived at.
REFERENCES
- El Karamany, F.M., 2006. Double purpose (forage and seed) of mungbean production 1-effect of plant density and forage cutting date on forage and seed yields of mungbean (Vigna radiate (L.) Wilczck). Res. J. Agric. Biol. Sci., 2: 162-165.
Direct Link - Omer, H.A.A.A., M.F. El Karamany, S.M. Ahmed, S.S. Abdel-Magid and B.A. Bakry, 2017. Using field crop by-products for feeding rabbits. Biosci. Res., 14: 224-233.
Direct Link - Naeem, M., I. Bhatti, R.H. Ahmad and M.Y. Ashraf, 2004. Effect of some growth hormones (GA3, IAA and kinetin) on the morphology and early or delayed initiation of bud of lentil (Lens culinaris Medik). Pak. J. Bot., 36: 801-809.
Direct Link - Shah, S., 2007. Effect of kinetin spray on growth and productivity of black cumin plants. Russian J. Plant Physiol., 54: 702-705.
CrossRefDirect Link - Amanullah, M.M., S. Sekar and S. Vincent, 2010. Plant growth substances in crop production: A review. Asian J. Plant Sci., 9: 215-222.
CrossRefDirect Link - Sadak, M.S., M.G. Dawood, B.A. Bakry and M.F. El-Karamany, 2013. Synergistic effect of indole acetic acid and kinetin on performance, some biochemical constituents and yield of faba bean plant grown under newly reclaimed sandy soil. World J. Agric. Sci., 9: 335-344.
Direct Link - Magome, H., S. Yamaguchi, A. Hanada, Y. Kamiya and K. Oda, 2004. Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J., 37: 720-729.
CrossRefPubMedDirect Link - Chapman, H.D. and P.F. Pratt, 1978. Methods of Analysis for Soils, Plant and Water. Califorina University Division Agriculture Science Priced Publication, Califorina, pp: 50-169.
Direct Link - Lichtenthaler, H.K. and C. Buschmann, 2001. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. In: Current Protocols in Food Analytical Chemistry, Wrolstad, R.E., T.E. Acree, H. An, E.A. Decker and M.H. Penner et al. (Eds.)., John Wiley and Sons, New York, USA, pp: F4.3.1-F4.3.8.
Direct Link - Shindy, W.W. and O.E. Smith, 1975. Identification of plant hormones from cotton ovules. Plant Physiol., 55: 550-554.
CrossRefDirect Link - Muller, P. and W. Hilgenberg, 1986. Isomers of zeatin and zeatin riboside in clubroot tissue: Evidence for trans‐zeatin biosynthesis by Plasmodiophora brassicae. Physiol. Planta., 66: 245-250.
CrossRefDirect Link - Herbert, D., P.J. Phipps and R.E. Strange, 1971. Chemical analysis of microbial cells. Methods Microbiol., 5: 209-344.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Miller, L. and J.A. Houghton, 1945. The micro-Kjeldahl determination of the nitrogen content of amino acids and proteins. J. Biol. Chem., 159: 373-383.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Kaya, C., M. Ashraf, M. Dikilitas and A.L. Tuna, 2013. Alleviation of salt stress-induced adverse effects on maize plants by exogenous application of indoleacetic acid (IAA) and inorganic nutrients-A field trial. Aust. J. Crop Sci., 7: 249-254.
Direct Link - MacDonald, H., 1997. Auxin perception and signal transduction. Physiol. Plant., 100: 423-430.
CrossRefDirect Link - El-Saeid, H.M., S.D. Abou-Hussein and W.A. El-Tohamy, 2010. Growth characters, yield and endogenous hormones of cowpea plants in response to IAA application. Res. J. Agric. Biol. Sci., 6: 27-31.
Direct Link - Barakat, N.A.M., 2011. Oxidative stress markers and antioxidant potential of wheat treated with phytohormones under salinity stress. J. Stress Physiol. Biochem., 7: 250-267.
Direct Link - Hopkins, W.G. and N.P.A. Hunter, 2004. Introduction to Plant Physiology. 3rd Edn., John Wiley and Sons Inc., New York, ISBN: 978-0-471-38915-6, Pages: 576.
Direct Link - Abdoli, M., M. Saeidi, S. Jalali-Honarmand and M. Azhand, 2013. The effect of foliar application of Indole-3-Acetic Acid (IAA) and roles of ear photosynthesis on grain yield production of two wheat cultivars (Triticum aestivum L.) under post anthesis water deficit. Int. Res. J. Applied Basic Sci., 4: 1406-1413.
Direct Link - Ubeda-Tomas, S., F. Federici, I. Casimiro, G.T. Beemster and R. Bhalerao et al., 2009. Gibberellin signaling in the endodermis controls Arabidopsis root meristem size. Curr. Biol., 19: 1194-1199.
CrossRefDirect Link - Ibrahim, M.E., M.A. Bekheta, A.D. El-Moursi and N.A. Gaafar, 2007. Improvement of growth and seed yield quality of (Vicia faba L.) plants as affected by application of some bioregulators. Aust. J. Basic Applied Sci., 1: 657-666.
Direct Link - Choudhury, S., N. Islam, M.D. Sarkar and M.A. Ali, 2013. Growth and yield of summer tomato as influenced by plant growth regulators. Int. J. Sustain. Agric., 5: 25-28.
Direct Link